Биологическая Адаптация — это… Что такое Биологическая Адаптация?
- Биологическая Адаптация
- Биологическая адаптация — приспособление организма к условиям окружающей среды (температура, атмосферное давление, влажность, освещенность и пр.). Для человека характерно то, что его биологическая адаптация может осуществляться с помощью различных вспомогательные средства (жилище, одежду), кроме того ему доступна произвольная психической регуляции ряда биологических процессов и состояний.
Психологический словарь. 2000.
- Биогенетический Закон
- Биологическая Обратная Связь
Смотреть что такое «Биологическая Адаптация» в других словарях:
биологическая адаптация — Этимология. Происходит от греч. biо жизнь и лат.
 adapto приспособляю. Категория. Приспособление организма к условиям окружающей среды. Специфика. Осуществляется по отношению к температуре, атмосферному давлению, влажности, освещенности и пр. Для… … Большая психологическая энциклопедия
adapto приспособляю. Категория. Приспособление организма к условиям окружающей среды. Специфика. Осуществляется по отношению к температуре, атмосферному давлению, влажности, освещенности и пр. Для… … Большая психологическая энциклопедияБиологическая адаптация — (от лат. аdaptatio приспособление), процесс приспособления организма к меняющимся условиям внешней среды в процессе эволюции, включающий комплекс морфофизиологических и поведенческих особенностей особи, популяции или вида, обеспечивающий: 1)… … Энциклопедический словарь по психологии и педагогике
Адаптация (биология) — Эта статья должна быть полностью переписана. На странице обсуждения могут быть пояснения. У этого термина существуют и другие значения, см. Адаптация … Википедия
Адаптация мигрантов — (от лат. adaptatio приспособление) адаптация как процесс приспособления человека к новым для него условиям жизни двойственна по своей природе, как двойственна и природа человека.
С одной стороны, это приспособление человека как живого существа, а … Миграция: словарь основных терминовАдаптация человека — одно из ключевых понятий в экологии человека, а также во многих других дисциплинах (физиологии, антропологии, медицинской географии, социологии, этнографии и пр.). Адаптация человека к новой для него среде сложный социально биологический процесс … Экология человека
адаптация — АДАПТАЦИЯ (от лат. adaptatio приспособление) форма освоения организмами воздействий внешней и внутренней среды, заключающаяся в тенденции к установлению с ними динамического равновесия. В процессе А. человека можно выделить два аспекта… … Энциклопедия эпистемологии и философии науки
Адаптация (биологическая) — АДАПТАЦИЯ (на средневековой латыни adaptatio приспособление), в биологии приспособление живых организмов к меняющимся условиям существования в результате изменения морфологических и физиологических особенностей и поведения.
Биологическая программа международная — (МБП; International Biological Program, IBP) многолетняя программа изучения в масштабе всей планеты биологической продуктивности (См. Биологическая продуктивность) естественных и созданных человеком растительных и животных сообществ… … Большая советская энциклопедия
Биологическая обратная связь (biofeedback) — Б. о. с. лучше всего понимать как замкнутый контур обратной связи. Этот контур состоит из организма как системы управления, реакции организма и средств обнаружения и отображения этой реакции для системы управления. Чтобы видоизменять обнаруженные … Психологическая энциклопедия
Биологическая эволюция — Молекула ДНК В биологии, эволюция это изменение наследственных признаков популяции организмов в течение нескольких поколений. Изменения вызываются взаимодействием трёх основных процессов: вариабельности, воспроизведения и селекции.
Гены, которые … Википедия
 adapto приспособляю. Категория. Приспособление организма к условиям окружающей среды. Специфика. Осуществляется по отношению к температуре, атмосферному давлению, влажности, освещенности и пр. Для… … Большая психологическая энциклопедия
adapto приспособляю. Категория. Приспособление организма к условиям окружающей среды. Специфика. Осуществляется по отношению к температуре, атмосферному давлению, влажности, освещенности и пр. Для… … Большая психологическая энциклопедия С одной стороны, это приспособление человека как живого существа, а … Миграция: словарь основных терминов
С одной стороны, это приспособление человека как живого существа, а … Миграция: словарь основных терминов
 Гены, которые … Википедия
Гены, которые … ВикипедияАдаптация (биология) — это… Что такое Адаптация (биология)?
У этого термина существуют и другие значения, см. Адаптация.Биологическая адаптация (от лат.
Приспособленность живых существ к естественным условиям внешней среды была осознана людьми ещё в античные времена.
Адаптации видов в рамках одного биоценоза зачастую тесно связаны друг с другом (одним из наиболее поразительных примеров межвидовой коадаптации является жёсткая привязка строения органов некоторых видов цветковых растений и насекомых друг к другу с целью опыления и питания). Если адаптационный процесс у какого-либо вида не находится в равновесном состоянии, то эволюционировать может весь биоценоз (иногда — с негативными последствиями) даже в стабильных условиях окружающей среды.
Проблема определения адаптации
Философ Ю. Урманцев указывает на то, что по объёму и содержанию «адаптация» — понятие не только междисциплинарное, но и весьма сложное. Он обращает внимание на недостатки определений, предлагаемые энциклопедиями, а именно [1]:
- тавтологичность и отсутствие указаний на существенные и специфические признаки, которые бы выделяли именно адаптацию ;
- охватывание определением «адаптация», как правило, только одного (обычно медико-биологического) вида адаптации из, во много раз большего, числа существующих;
- вызванное этим отсутствие корректного определения вообще, а тем самым и методологических указаний о возможных путях развития учения об адаптации.

Процесс развития жизни на земле предполагает наличие адаптации у организмов. Начинается эта адаптация с самых примитивных видов — приспособления к окружающей среде и к существующим условиям. Возникновение и выживание организмов возможно только при соответствии организмов окружающей среде. Выживают те организмы, которые вырабатывают лучшие формы своего сохранения. Их развитие, переход организмов на более высокую ступень обусловлены необходимостью адаптации. Таким образом, эволюция и адаптация суть процессы, неотделимые друг от друга.
Под адаптацией можно понимать так же способность любой системы получать новую информацию для приближения своего поведения и структуры к оптимальным. Системы адаптивны, если при изменении в их окружении или внутреннем состоянии, снижающем их эффективность в выполнении своих функций, они реагируют или откликаются, изменяя свое собственное состояние или состояние окружающей среды так, чтобы их эффективность увеличилась [2]. Термин адаптация можно рассматривать в трех аспектах:
- адаптация как свойство системы приспосабливаться к возможным изменениям функционирования —
- адаптация как сам процесс приспособления адаптивной системы — собственно, адаптация;
- адаптация как метод, основанный на обработке поступающей информации и приспособленный для достижения некоторого критерия оптимизации — адаптационные алгоритмы.

В более широком смысле, адаптациями в биологии называют возникновение и развитие определенных, конкретных морфофизиологических свойств, значения которых для организма связаны с теми или иными общими или частными условиями его абиотической и биотической среды.
Адаптация, как адаптационный ответ
Адаптация, как адаптационный ответ, может осуществляться на различных уровнях:
- на уровне клетки в виде функциональных или морфологических изменений;
- на уровне органа или группы клеток, имеющих одинаковую функцию;
- на уровне организма как морфологического так и функционального целого, представляющего собой совокупность всех физиологических функций, направленных на сохранение витальных функций и самой жизни.
С учетом этого H. Hensel выделяет различные уровни адаптационных процессов [3]:
- привыкание — начальный процесс адаптации под влиянием кратковременного воздействия стрессора,
- функциональную адаптацию — продолжительное состояние, возникающее под влиянием определенных раздражителей, приводящих к физиологическим изменениям гомеостаза человека,
- трофо-пластическую адаптацию — дальнейшая ступень адаптационных процессов, не принадлежит к терапевтической области реабилитационной медицины, так как при ней наступают морфологические изменения органов и систем человеческого организма.

Главное содержание адаптации, по мнению Т. Пилат, — это внутренние процессы в системе, которые обеспечивают сохранение её внешних функций по отношению к среде. Если структура системы обеспечивает ей нормальное функционирование в данных условиях среды, то такую систему следует считать адаптированной к этим условиям. На этой стадии устанавливается динамическое равновесие, при котором происходит изменение физиологических показателей в границах нормы. Организм клинически здорового человека с высокими адаптивными способностями обладает значительным потенциалом саморегуляции и самовосстановления. Для поддержания здоровья ему достаточно вести здоровый образ жизни, соблюдать принципы сбалансированного питания [4].
При этом, по мнению Ю. Малова, здоровье или норма — состояние относительно стабильное, но оно не может сохраняться в течение всей жизни, ибо в природе не существует абсолютно адаптированных к данной среде индивидов. Длительное пребывание в одних и тех же условиях всё же рано или поздно приведет к развитию какой-либо болезни, которая является механизмом элиминации индивида [5].
История представлений об адаптации в биологии и медицине
«Биологическая адаптивность и есть жизнь». Ганс Селье
В философских предпосылках
В античности Фалес Милетский, Парменид, Демокрит, Гиппократ, Платон, Аристотель, Плутарх, Гален, Флавий Ренат Вегеций, а также позже Авиценна в том или ином виде замечали и указывали, что заболевания, поведение и некоторые другие виды психической деятельности зависят от внешних факторов, внешней среды окружающей человека.
Парацельс — считал, что все органические функции вызваны действием единого универсального «жизненного принципа». Если деятельность «жизненного принципа» внутри формы происходит нормальным и регулярным образом, который не нарушают никакие препятствия (в том числе и внешние), такое состояние называется здоровьем. Если же его деятельность по какой-либо причине нарушается или если он действует ненормально или нерегулярно, такое состояние называется болезнью.
Мишель Монтень писал: «Мы знаем по опыту, что на нас оказывает влияние воздух, климат, земля того места, где мы родились; причем они влияют не только на цвет нашей кожи, на наш рост, телосложение и осанку, но и на наши душевные качества. Люди меняют свой нрав, если их переселить в другое место, совершенно так же, как и деревья» [6].
Люди меняют свой нрав, если их переселить в другое место, совершенно так же, как и деревья» [6].
В теориях эволюции
Основная статья: Адаптация (теории эволюции)
Тем не менее, научное обоснование идей адаптации разрабатывалось медленнее, чем философские воззрения и эмпирические знания на этот счёт и кардинально не менялось практически до 1809 года, когда французский естествоиспытатель Жан Батист Ламарк создал первое целостное учение об эволюции живой природы, основные идеи которого были изложены в «Философии зоологии».
На основе законов Ламарка сложилось направление, обычно называемое ламаркизмом или неоламаркизмом, а более точно эктогенезом — эктос — внешний (греч.), эктогенез — эволюция под действием внешних сил. В рамках этого направления эволюция рассматривалась как адаптивный процесс, основанный на всеобщем свойстве живых существ — «наследовании благоприобретённых свойств» (признаков). Следовательно, приспособленность рассматривалась как причина, а не результат эволюции. Единицей эволюции в рамках эктогенеза, является потомство родителей, которые передали своим потомкам результаты «упражнения или неупражнения органов», то есть череда поколений.
Единицей эволюции в рамках эктогенеза, является потомство родителей, которые передали своим потомкам результаты «упражнения или неупражнения органов», то есть череда поколений.
Дальнейшее развитие вопросы влияния внешней среды на человека получили в трудах английского естествоиспытателя, создателя теории эволюции Чарльза Роберта Дарвина. В 1859 году он выпустил книгу «Происхождение видов путём естественного отбора или сохранение благоприятствуемых пород в борьбе за жизнь», в которой было показано, как развивались все виды живого, согласно выдвинутому им положению о «выживании наиболее приспособленных».
Дарвин рассматривал приспособление (адаптацию) лишь в качестве средства для выживания. Согласно дарвинизму, эволюция представляет собой процесс адаптации организмов к условиям окружающей среды, причем приспособленность организмов является результатом эволюции.
В физиологии
Основная статья: Адаптация (физиология)
Во многом идеи К. Бернара получили продолжение в работах физиолога Ивана Петровича Павлова и американского ученого Уолтера Брейдфорда Кеннона.
Уолтер Кеннон, ввел в 1929 году в физиологию понятие гомеостаз считая, что постоянство внутренней среды организма поддерживается благодаря симпатической нервной системе и гормонам.
Категория поведения как особой формы саморегуляции жизни складывалась у И. П. Павлова (по образцу учения К. Бернара о гомеостазе), то есть об основных константах (давлении, температуре и др.), которые удерживают эту систему в стабильном состоянии в крайне неустойчивой внутренней среде. И. П. Павлов применил эту идею к взаимоотношениям организма с ещё более неустойчивой, непредсказуемой средой — внешней. Открытые им условно-рефлекторные механизмы служили достижению «уравновешивания» организма с этой средой.
Павловская трактовка рефлекса также имела четко выраженную гомеостатическую направленность и во многом базировалась на биодетерминизме К. Бернара, но не ограничивалась идеями гомеостаза. Известный американский физиолог Ч. Брукс отмечал: «Понятие об условном рефлексе предполагает появление длительных адаптивных изменений и даже включенность вегетативной системы» [7].
Связь с условным рефлексом
Условный рефлекс в трактовке И. П. Павлова, в первую очередь, — реакция адаптивного характера. И. П. Павлов уже в 1903 году в своей программной речи «Экспериментальная психология и психопатология на животных», на Международном медицинском конгрессе в Мадриде, впервые оповестил об открытии условных рефлексов, представив их как воплощение общебиологического закона приспособления организма к среде. В этом феномене, подчеркивал он, нет «ничего… кроме точной связи элементов сложной системы между собой и всего их комплекса с окружающей обстановкой». Он говорил тогда же: «К сожалению, мы не имеем до сих пор чисто научного термина для обозначения этого основного принципа организма — внутренней и внешней уравновешенности его» [8]. Через несколько десятков лет У. Кеннон изобрел этот термин, назвав «уравновешенность» гомеостазом. «Как часть природы, каждый животный организм представляет собою сложную обособленную систему, внутренние силы которой каждый момент, покуда она существует, как таковая, уравновешиваются с внешними силами окружающей среды. Чем сложнее организм, тем тоньше, многочисленнее и разнообразнее элементы уравновешивания. Для этого служат анализаторы и механизмы как постоянных, так и временных связей, устанавливающие точнейшие соотношения между мельчайшими элементами внешнего мира и тончайшими реакциями животного организма. Таким образом, вся жизнь от простейших до сложнейших организмов, включая, конечно, и человека, есть длинный ряд все усложняющихся до высочайшей степени уравновешиваний внешней среды» [9].
Чем сложнее организм, тем тоньше, многочисленнее и разнообразнее элементы уравновешивания. Для этого служат анализаторы и механизмы как постоянных, так и временных связей, устанавливающие точнейшие соотношения между мельчайшими элементами внешнего мира и тончайшими реакциями животного организма. Таким образом, вся жизнь от простейших до сложнейших организмов, включая, конечно, и человека, есть длинный ряд все усложняющихся до высочайшей степени уравновешиваний внешней среды» [9].
И. П. Павлов и У. Кеннон, в своих работах переходили к поведению целостного организма, то есть к особому типу его взаимодействия со средой. Ведь и условные рефлексы, и эмоции страха и ярости, ставшие (наряду с болью и голодом) главным объектом направления кенноновских исследований, выступают в качестве реакций, которые носят особый интегральный характер. Они даны в системе «организм — среда» и не могут быть объяснены вне её. Это не значит, что великий принцип единства организма и среды действует в полную силу лишь с переходом к условнорефлекторным и эмоциональным регуляциям. Речь идет о различных формах реализации этого принципа. Он выступает на многих уровнях: молекулярном, энергетическом, химическом, а также на уровнях функционирования различных физиологических и психологических систем. В условном рефлексе как поведенческом акте задействован физиологический механизм (по И. П. Павлову — кора и ближайшая к ней подкорка), но он становится поведенческим только тогда, когда в нём представлены условия среды в виде различаемых мозгом внешних (средовых) раздражителей, играющих роль сигналов.
Речь идет о различных формах реализации этого принципа. Он выступает на многих уровнях: молекулярном, энергетическом, химическом, а также на уровнях функционирования различных физиологических и психологических систем. В условном рефлексе как поведенческом акте задействован физиологический механизм (по И. П. Павлову — кора и ближайшая к ней подкорка), но он становится поведенческим только тогда, когда в нём представлены условия среды в виде различаемых мозгом внешних (средовых) раздражителей, играющих роль сигналов.
Множество операций, проведенных в кенноновской школе над животными, доказали, что внешне наблюдаемые признаки поведения, которые можно назвать эмоциональными, порождаются глубинными сдвигами в нейрогуморальных процессах. Эти сдвиги готовят организм к критическим ситуациям, которые требуют повышенной траты энергии, снятия усталости, предотвращения кровопотери и т. п. На одном из докладов о своих открытиях У. Кеннон сообщил, что благодаря выбрасываемому в кровь при сильных эмоциях адреналину в числе других его «мобилизационных» эффектов происходит увеличение в крови поступающего к мышцам сахара.
У. Кеннон в поисках нового языка для обсуждения эмоционального поведения нашёл его у И. П. Павлова. Поведение трактуется им в терминах рефлексов — безусловных и условных. Главная эмоция для У. Кеннона отныне не что иное, как безусловный рефлекс. Картина поведения таламического, лишенного высших нервных центров, животного свидетельствует, согласно У. Кеннону, в пользу положения о том, что эмоция в её внешнем выражении является сложным безусловным рефлексом. Кеннон писал: «Жизнь избегает грубых аритмичных, неуправляемых взрывов. Небольшие изменения, маленькие химические реакции при умеренной температуре дают организму сопротивляемость крепче стали и направляются с точностью и тонкостью, не свойственной технике термитов. В этом «великая мудрость организма» [10].
Саморегуляция
И. П. Павлов в общей форме выдвинул принцип саморегуляции при исследовании деятельности сердца и кровообращения, а позднее распространил его на деятельность организма в целом. В работе «Ответ физиолога психологам» (1951) он отметил: «Человек есть, конечно, система (грубо говоря, — машина), как и всякая другая в природе, подчиняющаяся неизбежным и единым для всей природы законам, но система, в горизонте нашего современного видения, единственная по высочайшему саморегулированию…». Помимо учения о высшей нервной деятельности, определившего «принцип нервизма» Павлов обратил внимание на ферменты назвав их «возбудителями жизни». По представлению И. П. Павлова (1949) «… животный организм как система существует среди окружающей природы только благодаря непрерывному уравновешиванию этой системы на падающие на нее из вне раздражения» [11].
Помимо учения о высшей нервной деятельности, определившего «принцип нервизма» Павлов обратил внимание на ферменты назвав их «возбудителями жизни». По представлению И. П. Павлова (1949) «… животный организм как система существует среди окружающей природы только благодаря непрерывному уравновешиванию этой системы на падающие на нее из вне раздражения» [11].
Таким образом, английская мысль внесла идею адаптации к среде внешней, как задаче, непрерывно решаемой организмом, французская мысль — идею саморегуляции процессов в этом организме, а русская — категорию поведения как особой формы саморегуляции жизни, что прекратило длившиеся многие годы прошлого столетия споры между «нервистами» и «гуморалистами», представляющие ныне лишь исторический интерес.
Адаптация и стресс
Ганс Селье вводит в медицинскую практику термин стресс. Он писал: «я впервые „наткнулся“ на идею стресса и общего адаптационного синдрома в 1925 году» [12]. Каждую составляющую своего определения Селье пояснил так: общий — потому что к стрессу приводят факторы, которые, воздействуя на разные области организма, в итоге способны вызвать общую системную защиту; адаптационный — потому что это явление как бы закрепляется, приобретает характер привычки; синдром — потому что его отдельные проявления частично взаимозависимы [13]. Позже (в 1931-1932 г.) он назвал стресс неспецифической реакцией организма на любые раздражители. Представление о стрессе касается всех людей, больных и здоровых, преуспевающих и неудачливых, и всех сторон жизни. «Удалось показать, что стресс представляет собой скорость изнашивания человеческого организма, сопровождает любую жизнедеятельность и соответствует в определённом смысле интенсивности жизни. Он увеличивается при нервном напряжении, телесных повреждениях, инфекциях, мышечной работе или любой другой напряженной деятельности и связан с неспецифическим защитным механизмом, увеличивающим сопротивляемость к стрессовым факторам, или „стрессорам“. Важной частью этого защитного механизма является повышенное выделение гипофизом так называемого адренокортикотропного гормона (АКТГ), который, в свою очередь, стимулирует выработку кортикоидов корой надпочечников.
Позже (в 1931-1932 г.) он назвал стресс неспецифической реакцией организма на любые раздражители. Представление о стрессе касается всех людей, больных и здоровых, преуспевающих и неудачливых, и всех сторон жизни. «Удалось показать, что стресс представляет собой скорость изнашивания человеческого организма, сопровождает любую жизнедеятельность и соответствует в определённом смысле интенсивности жизни. Он увеличивается при нервном напряжении, телесных повреждениях, инфекциях, мышечной работе или любой другой напряженной деятельности и связан с неспецифическим защитным механизмом, увеличивающим сопротивляемость к стрессовым факторам, или „стрессорам“. Важной частью этого защитного механизма является повышенное выделение гипофизом так называемого адренокортикотропного гормона (АКТГ), который, в свою очередь, стимулирует выработку кортикоидов корой надпочечников.
Cиндром стресса, или, иначе, общий адаптационный синдром (ОАС), проходит три стадии:
- „реакция тревоги“, во время которой мобилизуются защитные силы. Её биологический смысл заключается в том, что организм в кратчайшие сроки должен получить дополнительную, „аварийную“ энергию для того, чтобы максимально обеспечить условия для быстрого спасения от грозящей беды или даже гибели. Да, для организма это, безусловно, реакция энергозатратная — что для него в перспективе, конечно, плохо, — но иного выхода в данный момент нет. Ведь речь идет о спасении в целом.
- „стадия устойчивости“ отражающая полную адаптацию к стрессору. Крайне важно и то, что на этой стадии общие энергозатраты организма становятся меньше, чем на первой стадии: организм частично уже приспособился жить под давлением стрессирующего фактора — как бы отследил его. Если стрессирующий фактор действует сильно и длительно, то постепенно развивается следующая, третья стадия.
- „стадия истощения“, которая неумолимо наступает, если стрессор оказывается достаточно силён и действует достаточно долгое время, поскольку „адаптационная энергия“, или приспособляемость живого вещества всегда конечна» [14]. Результат на этой стадии — устойчивость организма к внешним воздействиям резко снижается. То есть база для возможного развития патологии на клиническом уровне уже создана.
 Её биологический смысл заключается в том, что организм в кратчайшие сроки должен получить дополнительную, „аварийную“ энергию для того, чтобы максимально обеспечить условия для быстрого спасения от грозящей беды или даже гибели. Да, для организма это, безусловно, реакция энергозатратная — что для него в перспективе, конечно, плохо, — но иного выхода в данный момент нет. Ведь речь идет о спасении в целом.
Её биологический смысл заключается в том, что организм в кратчайшие сроки должен получить дополнительную, „аварийную“ энергию для того, чтобы максимально обеспечить условия для быстрого спасения от грозящей беды или даже гибели. Да, для организма это, безусловно, реакция энергозатратная — что для него в перспективе, конечно, плохо, — но иного выхода в данный момент нет. Ведь речь идет о спасении в целом. Результат на этой стадии — устойчивость организма к внешним воздействиям резко снижается. То есть база для возможного развития патологии на клиническом уровне уже создана.
Результат на этой стадии — устойчивость организма к внешним воздействиям резко снижается. То есть база для возможного развития патологии на клиническом уровне уже создана.По мере формирования устойчивой адаптации нарушения гомеостаза, составляющие стимул стресс-симптома постепенно исчезают, как и сам стресс-симптом, сыграв свою важную роль в становлении адаптации. Это состояние между стрессом (агрессией) и адаптацией служит доказательством того, что стресс сложился в процессе эволюции как необходимое неспецифическое звено более сложного целостного механизма адаптации. Полная свобода от стресса, как считал Селье, означала бы смерть. В 1982 году Селье определил стресс как совокупность стереотипных филогенетических запрограммированных реакций организма, которые вызываются любыми сильными, сверхсильными, экстремальными воздействиями и сопровождаются перестройкой адаптивных сил организма.[15].
По признанию Г. Селье, он заимствовал у И. П. Павлова очень многое.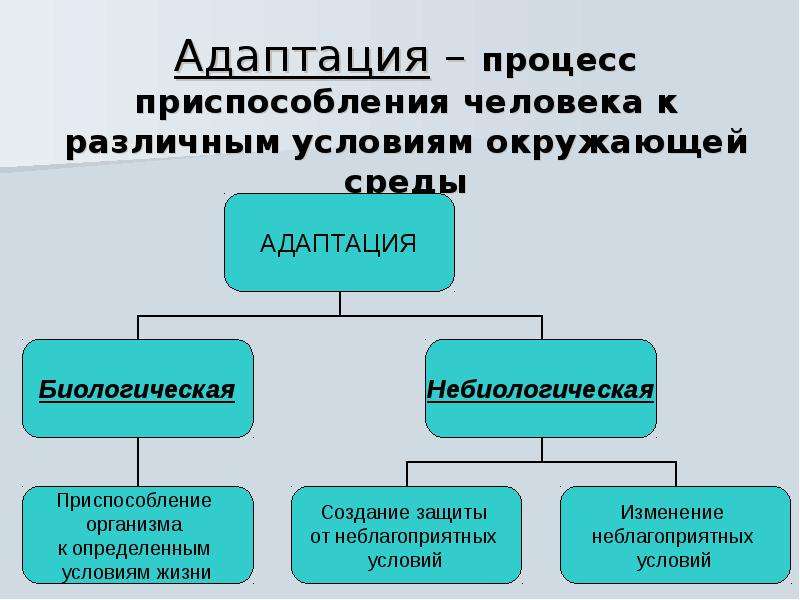 То, что Павлов трактовал с точки зрения нервной системы, он перевел на язык и термины гуморальной системы. Павлов указывал, что организм как целое непрерывно адаптируется к окружающей среде. Эту идею целостности и адаптации Селье почерпнул у Павлова, и это было положено в основу теории стресса. То, что внешние воздействия приводят только к расходу и исчерпанию адаптационных возможностей, в течение многих лет было убеждением Г. Селье, которое он лаконично формулировал как «wear and tear». Это убеждение основывалось на том, что он использовал преимущественно сильные патогенные воздействия.
То, что Павлов трактовал с точки зрения нервной системы, он перевел на язык и термины гуморальной системы. Павлов указывал, что организм как целое непрерывно адаптируется к окружающей среде. Эту идею целостности и адаптации Селье почерпнул у Павлова, и это было положено в основу теории стресса. То, что внешние воздействия приводят только к расходу и исчерпанию адаптационных возможностей, в течение многих лет было убеждением Г. Селье, которое он лаконично формулировал как «wear and tear». Это убеждение основывалось на том, что он использовал преимущественно сильные патогенные воздействия.
Концепции о существовании генетически предопределённого потенциала роста берут начало от А. Вейсмана, который считал, что зигота обладает способностью осуществлять определённое и ограниченное число делений клеток, после чего развивающийся организма обрекается на старение и смерть. Немецкий физиолог и гигиенист Макс Рубнер (М. Rubner) (1854—1932) в связи с представлениями Вейсмана обосновал понятие о генетически предопределённом энергетическом фонде, который у всех видов млекопитающих, за исключением человека почти одинаков. Концепцию Рубнера и вытекающее из неё представление об онтогенезе, как о раскручивании пружины в заведенных часах, и развил «Селье в учении о стрессе и о предопределённом фонде „адаптивной энергии“, сравнивая его с банковым вложением, которое можно расходовать, но нельзя увеличивать». Во всех этих теориях речь идет о постепенном исчерпании некоего исходного потенциала (жизненной субстанции), приводящих живые организмы к состоянию равновесия. Развивая эти представления Ричард Перл пришёл к мысли, что старение и смерть — „расплата за преимущества“ какие дают специализация и дифференциация клеток у многоклеточных организмов, в отличие от практически бессмертных одноклеточных этими качествами не обладающими.[16].
Концепцию Рубнера и вытекающее из неё представление об онтогенезе, как о раскручивании пружины в заведенных часах, и развил «Селье в учении о стрессе и о предопределённом фонде „адаптивной энергии“, сравнивая его с банковым вложением, которое можно расходовать, но нельзя увеличивать». Во всех этих теориях речь идет о постепенном исчерпании некоего исходного потенциала (жизненной субстанции), приводящих живые организмы к состоянию равновесия. Развивая эти представления Ричард Перл пришёл к мысли, что старение и смерть — „расплата за преимущества“ какие дают специализация и дифференциация клеток у многоклеточных организмов, в отличие от практически бессмертных одноклеточных этими качествами не обладающими.[16].
«Стресс есть неспецифический ответ организма на любое предъявление ему требования. … С точки зрения стрессовой реакции не имеет значения, приятна или неприятна ситуация, с которой мы столкнулись. Имеет значение лишь интенсивность потребности в перестройке или в адаптации. » [17]
» [17]
Измерение адаптированности
В результате обработки многолетних наблюдений при сравнительном анализе популяций и групп, находящихся в различных экологических условиях (на Крайнем Севере и в средних широтах Сибири), А. Н. Горбанем и Е. И. Смирновой под руководством ак. АМН СССР К. Р. Седова получен вывод: наибольшую информацию о степени адаптированности популяции к экстремальным или просто изменившимся условиям несут корреляции между физиологическими параметрами.
В типичной ситуации при увеличении адаптационной нагрузки уровень корреляций повышается, а в результате успешной адаптации — снижается. Показано, что этот эффект группового стресса связан с организацией системы факторов, действующих на группу.
Если система факторов организована в соответствии с принципом Либиха (монолимитирование) то с ростом адаптационного напряжения размерность облака точек, представляющего группу в пространстве физиологических параметров, снижается, растет сумма абсолютных величин коэффициентов корреляции и, в то же время, растет разброс этого облака — возрастают дисперсии.
Если происходит взаимное усиление давления различных факторов (синергичные системы), то, напротив, с ростом адаптационного напряжения размерность облака точек, представляющего группу в пространстве физиологических параметров, растет, падает сумма коэффициентов корреляции. Дисперсии в то же время возрастают — так же, как и в предыдущем случае.
Литература
- Агаджанян Н. А., Труханов А. И., Шендеров Б. А. Этюды об адаптации и путях сохранения здоровья. — М.: изд-во «Сирин», — 2002. — 156 с.
- Ader Robert, Cohen Nicolas, Felten David, Psychoneuroimmunology: interactions between the nervous system and the immune system, Lancet 1995; 345:99-103
- Акофф Р., Эмерли Ф. О целеустремленных системах. — М.: Советское радио, 1974. — 272 с.
- Анохин П. К. Очерки по физиологии функциональных систем. — М.: Медицина, −1975.- 477 с.
- Анохин П. К.Узловые вопросы теории функциональной системы. — М.: Наука, −1980. — 197 с.
- Аршавский И. А.Физиологические механизмы и закономерности индивидуального развития. М.: Наука, — 1982. 270 с.
- Аршавский И. А. Некоторые методологические и теоретические аспекты анализа закономерностей индивидуального развития организмов//Вопросы философии. — № 11. — 1986. — С. 95-104.
- Бернар Кл. Лекции по экспериментальной патологии // Пер. Д. Е. Жуковского. М.; Л., 1937, с. 312—374.
- Бородюк Н. Р. Адаптация. Новое в приспособлении к окружающей среде. М.: «Глобус», 1998. — 88 с.
- Воробьёв Р. И. Медицина по Дарвину// Химия и жизнь. — № 2. — 2001. — с.44-48.
- Гартман Франц Жизнь Парацельса и сущность его учения/Пер. с англ. — М.: Алетейа, 2002. — 272с
- Дербенёва Л. М. Интимная жизнь стресса// Химия и жизнь. — № 7. — 1999 г. — С.54-57.
- Калайков И. Цивилизация и адаптация, М.: «Прогресс», 1984.
- Кассиль Г. Н. Внутренняя среда организма М.: Наука, 1983. — 227 с.
- Козинец Г. И., Погорелов В. М. Консерватизм биологических процессов — стабильность кроветворения Клиническая лабораторная диагностика № 12, 1988.-С.21-32.
- Малов Ю. С. Адаптация и здоровье. Клиническая медицина № 12. — 2001.- С. 61-63.
- Меерсон Ф. З., Пшенникова М. Г. Адаптация к стрессовым ситуациям и физическим нагрузкам. — М.: Медицина, 1988. — 256 с.
- Монтень Мишель, Опыты. Книга третья. Изд-во «Наука». — Москва. — 1981. — 535 с.
- Сапов И. А., Новиков В. С. Неспецифические механизмы адаптации человека. — Л.: Наука, 1984. — 146 с.
- Светуньков С. Г. Количественные методы прогнозирования эволюционных составляющих экономической динамики. — Ульяновск: Изд-во Ульяновского государственного университета, 1999. —177 с.
- Северцов А. С. Теория эволюции. — М.: Гуманитар. изд. центр ВЛАДОС, 2005. — 380с
- Скулачев В. П. Эволюция биологических механизмов запасания энергии // Соросовский Образовательный Журнал. 1997. № 5. С. 11-19.
- Урманцев Ю. А. Природа адаптации (системная экспликация)//«Вопросы философии». — 1998. — № 12.
- Философские проблемы адаптации Под ред. Г. И. Царегородцева. М., «Мысль», 1975 277 с.
- Hensel H. Grundbegriffe und neuere Aspekte der physiologischen Adaptation,

 1997. № 5. С. 11-19.
1997. № 5. С. 11-19.См. также
Примечания
- ↑ (Ю. А. Урманцев — 1998 г.)
- ↑ Акофф Р., Эмерли Ф. — 1974 г.
- ↑ (Hensel H. — 1974 г.)
- ↑ (Пилат Т. Л., Иванов А. А. — 2002 г.)
- ↑ (Малов Ю. С.- 2001 г.)
- ↑ (цит. по С. 621. — Монтень М. — 1981 г.)
- ↑ (цит. по С.57. — Ярошевский М. Г. — 1995 г.)
- ↑ (цит. по С.57.Ярошевский М. Г. — 1995 г.)
- ↑ (цит. по С.22. — «Химия и жизнь». — № 3. — 1990г)
- ↑ (цит. по С.13 Залманов А. С. — 1991 г.)
- ↑ (цит. по С. 8. — Агаджанян Н. А., Труханов А. И., Шендеров Б. А. — 2002 г. )
- ↑ (цит. по С.68. — Селье Г. — 1987 г.)
- ↑ (Дербенёва Л. М. — 1999 г.)
- ↑ (цит. по С.71-72 Селье Г. — 1987 г.)
- ↑ (Никонов В. В. — 2002 г.)
- ↑ цит. по С.15. Аршавский И. А. −1982., С. 95 Аршавский И. А.- 1986 г.
- ↑ Селье Г., Стресс жизни.
 )
)Адаптация (биология) — это… Что такое Адаптация (биология)?
У этого термина существуют и другие значения, см. Адаптация.Биологическая адаптация (от лат. adaptatio — приспособление) — приспособление организма к внешним условиям в процессе эволюции, включая морфофизиологическую и поведенческую составляющие. Адаптация может обеспечивать выживаемость в условиях конкретного местообитания, устойчивость к воздействию факторов абиотического и биологического характера, а также успех в конкуренции с другими видами, популяциями, особями. Каждый вид имеет собственную способность к адаптации, ограниченную физиологией (индивидуальная адаптация), пределами проявления материнского эффекта и модификаций, эпигенетическим разнообразием, внутривидовой изменчивостью, мутационными возможностями, коадаптационными характеристиками внутренних органов и другими видовыми особенностями.
Приспособленность живых существ к естественным условиям внешней среды была осознана людьми ещё в античные времена. Вплоть до середины XIX века это объяснялось изначальной целесообразностью природы. В теории эволюции Чарльза Дарвина было предложено научное объяснение адаптационного процесса на основе естественного отбора.
Адаптации видов в рамках одного биоценоза зачастую тесно связаны друг с другом (одним из наиболее поразительных примеров межвидовой коадаптации является жёсткая привязка строения органов некоторых видов цветковых растений и насекомых друг к другу с целью опыления и питания). Если адаптационный процесс у какого-либо вида не находится в равновесном состоянии, то эволюционировать может весь биоценоз (иногда — с негативными последствиями) даже в стабильных условиях окружающей среды.
Проблема определения адаптации
Философ Ю. Урманцев указывает на то, что по объёму и содержанию «адаптация» — понятие не только междисциплинарное, но и весьма сложное. Он обращает внимание на недостатки определений, предлагаемые энциклопедиями, а именно [1]:
Он обращает внимание на недостатки определений, предлагаемые энциклопедиями, а именно [1]:
- тавтологичность и отсутствие указаний на существенные и специфические признаки, которые бы выделяли именно адаптацию ;
- охватывание определением «адаптация», как правило, только одного (обычно медико-биологического) вида адаптации из, во много раз большего, числа существующих;
- вызванное этим отсутствие корректного определения вообще, а тем самым и методологических указаний о возможных путях развития учения об адаптации.
Процесс развития жизни на земле предполагает наличие адаптации у организмов. Начинается эта адаптация с самых примитивных видов — приспособления к окружающей среде и к существующим условиям. Возникновение и выживание организмов возможно только при соответствии организмов окружающей среде. Выживают те организмы, которые вырабатывают лучшие формы своего сохранения. Их развитие, переход организмов на более высокую ступень обусловлены необходимостью адаптации. Таким образом, эволюция и адаптация суть процессы, неотделимые друг от друга.
Таким образом, эволюция и адаптация суть процессы, неотделимые друг от друга.
Под адаптацией можно понимать так же способность любой системы получать новую информацию для приближения своего поведения и структуры к оптимальным. Системы адаптивны, если при изменении в их окружении или внутреннем состоянии, снижающем их эффективность в выполнении своих функций, они реагируют или откликаются, изменяя свое собственное состояние или состояние окружающей среды так, чтобы их эффективность увеличилась [2]. Термин адаптация можно рассматривать в трех аспектах:
- адаптация как свойство системы приспосабливаться к возможным изменениям функционирования — система адаптации;
- адаптация как сам процесс приспособления адаптивной системы — собственно, адаптация;
- адаптация как метод, основанный на обработке поступающей информации и приспособленный для достижения некоторого критерия оптимизации — адаптационные алгоритмы.
В более широком смысле, адаптациями в биологии называют возникновение и развитие определенных, конкретных морфофизиологических свойств, значения которых для организма связаны с теми или иными общими или частными условиями его абиотической и биотической среды.
Адаптация, как адаптационный ответ
Адаптация, как адаптационный ответ, может осуществляться на различных уровнях:
- на уровне клетки в виде функциональных или морфологических изменений;
- на уровне органа или группы клеток, имеющих одинаковую функцию;
- на уровне организма как морфологического так и функционального целого, представляющего собой совокупность всех физиологических функций, направленных на сохранение витальных функций и самой жизни.
С учетом этого H. Hensel выделяет различные уровни адаптационных процессов [3]:
- привыкание — начальный процесс адаптации под влиянием кратковременного воздействия стрессора,
- функциональную адаптацию — продолжительное состояние, возникающее под влиянием определенных раздражителей, приводящих к физиологическим изменениям гомеостаза человека,
- трофо-пластическую адаптацию — дальнейшая ступень адаптационных процессов, не принадлежит к терапевтической области реабилитационной медицины, так как при ней наступают морфологические изменения органов и систем человеческого организма.
Главное содержание адаптации, по мнению Т. Пилат, — это внутренние процессы в системе, которые обеспечивают сохранение её внешних функций по отношению к среде. Если структура системы обеспечивает ей нормальное функционирование в данных условиях среды, то такую систему следует считать адаптированной к этим условиям. На этой стадии устанавливается динамическое равновесие, при котором происходит изменение физиологических показателей в границах нормы. Организм клинически здорового человека с высокими адаптивными способностями обладает значительным потенциалом саморегуляции и самовосстановления. Для поддержания здоровья ему достаточно вести здоровый образ жизни, соблюдать принципы сбалансированного питания [4].
При этом, по мнению Ю. Малова, здоровье или норма — состояние относительно стабильное, но оно не может сохраняться в течение всей жизни, ибо в природе не существует абсолютно адаптированных к данной среде индивидов. Длительное пребывание в одних и тех же условиях всё же рано или поздно приведет к развитию какой-либо болезни, которая является механизмом элиминации индивида [5].
История представлений об адаптации в биологии и медицине
«Биологическая адаптивность и есть жизнь». Ганс Селье
В философских предпосылках
В античности Фалес Милетский, Парменид, Демокрит, Гиппократ, Платон, Аристотель, Плутарх, Гален, Флавий Ренат Вегеций, а также позже Авиценна в том или ином виде замечали и указывали, что заболевания, поведение и некоторые другие виды психической деятельности зависят от внешних факторов, внешней среды окружающей человека.
Парацельс — считал, что все органические функции вызваны действием единого универсального «жизненного принципа». Если деятельность «жизненного принципа» внутри формы происходит нормальным и регулярным образом, который не нарушают никакие препятствия (в том числе и внешние), такое состояние называется здоровьем. Если же его деятельность по какой-либо причине нарушается или если он действует ненормально или нерегулярно, такое состояние называется болезнью.
Мишель Монтень писал: «Мы знаем по опыту, что на нас оказывает влияние воздух, климат, земля того места, где мы родились; причем они влияют не только на цвет нашей кожи, на наш рост, телосложение и осанку, но и на наши душевные качества. Люди меняют свой нрав, если их переселить в другое место, совершенно так же, как и деревья» [6].
В теориях эволюции
Основная статья: Адаптация (теории эволюции)
Тем не менее, научное обоснование идей адаптации разрабатывалось медленнее, чем философские воззрения и эмпирические знания на этот счёт и кардинально не менялось практически до 1809 года, когда французский естествоиспытатель Жан Батист Ламарк создал первое целостное учение об эволюции живой природы, основные идеи которого были изложены в «Философии зоологии».
На основе законов Ламарка сложилось направление, обычно называемое ламаркизмом или неоламаркизмом, а более точно эктогенезом — эктос — внешний (греч.), эктогенез — эволюция под действием внешних сил. В рамках этого направления эволюция рассматривалась как адаптивный процесс, основанный на всеобщем свойстве живых существ — «наследовании благоприобретённых свойств» (признаков). Следовательно, приспособленность рассматривалась как причина, а не результат эволюции. Единицей эволюции в рамках эктогенеза, является потомство родителей, которые передали своим потомкам результаты «упражнения или неупражнения органов», то есть череда поколений.
Дальнейшее развитие вопросы влияния внешней среды на человека получили в трудах английского естествоиспытателя, создателя теории эволюции Чарльза Роберта Дарвина. В 1859 году он выпустил книгу «Происхождение видов путём естественного отбора или сохранение благоприятствуемых пород в борьбе за жизнь», в которой было показано, как развивались все виды живого, согласно выдвинутому им положению о «выживании наиболее приспособленных».
Дарвин рассматривал приспособление (адаптацию) лишь в качестве средства для выживания. Согласно дарвинизму, эволюция представляет собой процесс адаптации организмов к условиям окружающей среды, причем приспособленность организмов является результатом эволюции.
В физиологии
Основная статья: Адаптация (физиология)
Во многом идеи К. Бернара получили продолжение в работах физиолога Ивана Петровича Павлова и американского ученого Уолтера Брейдфорда Кеннона.
Уолтер Кеннон, ввел в 1929 году в физиологию понятие гомеостаз считая, что постоянство внутренней среды организма поддерживается благодаря симпатической нервной системе и гормонам.
Категория поведения как особой формы саморегуляции жизни складывалась у И. П. Павлова (по образцу учения К. Бернара о гомеостазе), то есть об основных константах (давлении, температуре и др.), которые удерживают эту систему в стабильном состоянии в крайне неустойчивой внутренней среде. И. П. Павлов применил эту идею к взаимоотношениям организма с ещё более неустойчивой, непредсказуемой средой — внешней. Открытые им условно-рефлекторные механизмы служили достижению «уравновешивания» организма с этой средой.
Павловская трактовка рефлекса также имела четко выраженную гомеостатическую направленность и во многом базировалась на биодетерминизме К. Бернара, но не ограничивалась идеями гомеостаза. Известный американский физиолог Ч. Брукс отмечал: «Понятие об условном рефлексе предполагает появление длительных адаптивных изменений и даже включенность вегетативной системы» [7].
Связь с условным рефлексом
Условный рефлекс в трактовке И. П. Павлова, в первую очередь, — реакция адаптивного характера. И. П. Павлов уже в 1903 году в своей программной речи «Экспериментальная психология и психопатология на животных», на Международном медицинском конгрессе в Мадриде, впервые оповестил об открытии условных рефлексов, представив их как воплощение общебиологического закона приспособления организма к среде. В этом феномене, подчеркивал он, нет «ничего… кроме точной связи элементов сложной системы между собой и всего их комплекса с окружающей обстановкой». Он говорил тогда же: «К сожалению, мы не имеем до сих пор чисто научного термина для обозначения этого основного принципа организма — внутренней и внешней уравновешенности его» [8]. Через несколько десятков лет У. Кеннон изобрел этот термин, назвав «уравновешенность» гомеостазом. «Как часть природы, каждый животный организм представляет собою сложную обособленную систему, внутренние силы которой каждый момент, покуда она существует, как таковая, уравновешиваются с внешними силами окружающей среды. Чем сложнее организм, тем тоньше, многочисленнее и разнообразнее элементы уравновешивания. Для этого служат анализаторы и механизмы как постоянных, так и временных связей, устанавливающие точнейшие соотношения между мельчайшими элементами внешнего мира и тончайшими реакциями животного организма. Таким образом, вся жизнь от простейших до сложнейших организмов, включая, конечно, и человека, есть длинный ряд все усложняющихся до высочайшей степени уравновешиваний внешней среды» [9].
И. П. Павлов и У. Кеннон, в своих работах переходили к поведению целостного организма, то есть к особому типу его взаимодействия со средой. Ведь и условные рефлексы, и эмоции страха и ярости, ставшие (наряду с болью и голодом) главным объектом направления кенноновских исследований, выступают в качестве реакций, которые носят особый интегральный характер. Они даны в системе «организм — среда» и не могут быть объяснены вне её. Это не значит, что великий принцип единства организма и среды действует в полную силу лишь с переходом к условнорефлекторным и эмоциональным регуляциям. Речь идет о различных формах реализации этого принципа. Он выступает на многих уровнях: молекулярном, энергетическом, химическом, а также на уровнях функционирования различных физиологических и психологических систем. В условном рефлексе как поведенческом акте задействован физиологический механизм (по И. П. Павлову — кора и ближайшая к ней подкорка), но он становится поведенческим только тогда, когда в нём представлены условия среды в виде различаемых мозгом внешних (средовых) раздражителей, играющих роль сигналов.
Множество операций, проведенных в кенноновской школе над животными, доказали, что внешне наблюдаемые признаки поведения, которые можно назвать эмоциональными, порождаются глубинными сдвигами в нейрогуморальных процессах. Эти сдвиги готовят организм к критическим ситуациям, которые требуют повышенной траты энергии, снятия усталости, предотвращения кровопотери и т. п. На одном из докладов о своих открытиях У. Кеннон сообщил, что благодаря выбрасываемому в кровь при сильных эмоциях адреналину в числе других его «мобилизационных» эффектов происходит увеличение в крови поступающего к мышцам сахара.
У. Кеннон в поисках нового языка для обсуждения эмоционального поведения нашёл его у И. П. Павлова. Поведение трактуется им в терминах рефлексов — безусловных и условных. Главная эмоция для У. Кеннона отныне не что иное, как безусловный рефлекс. Картина поведения таламического, лишенного высших нервных центров, животного свидетельствует, согласно У. Кеннону, в пользу положения о том, что эмоция в её внешнем выражении является сложным безусловным рефлексом. Кеннон писал: «Жизнь избегает грубых аритмичных, неуправляемых взрывов. Небольшие изменения, маленькие химические реакции при умеренной температуре дают организму сопротивляемость крепче стали и направляются с точностью и тонкостью, не свойственной технике термитов. В этом «великая мудрость организма» [10].
Саморегуляция
И. П. Павлов в общей форме выдвинул принцип саморегуляции при исследовании деятельности сердца и кровообращения, а позднее распространил его на деятельность организма в целом. В работе «Ответ физиолога психологам» (1951) он отметил: «Человек есть, конечно, система (грубо говоря, — машина), как и всякая другая в природе, подчиняющаяся неизбежным и единым для всей природы законам, но система, в горизонте нашего современного видения, единственная по высочайшему саморегулированию…». Помимо учения о высшей нервной деятельности, определившего «принцип нервизма» Павлов обратил внимание на ферменты назвав их «возбудителями жизни». По представлению И. П. Павлова (1949) «… животный организм как система существует среди окружающей природы только благодаря непрерывному уравновешиванию этой системы на падающие на нее из вне раздражения» [11].
Таким образом, английская мысль внесла идею адаптации к среде внешней, как задаче, непрерывно решаемой организмом, французская мысль — идею саморегуляции процессов в этом организме, а русская — категорию поведения как особой формы саморегуляции жизни, что прекратило длившиеся многие годы прошлого столетия споры между «нервистами» и «гуморалистами», представляющие ныне лишь исторический интерес.
Адаптация и стресс
Ганс Селье вводит в медицинскую практику термин стресс. Он писал: «я впервые „наткнулся“ на идею стресса и общего адаптационного синдрома в 1925 году» [12]. Каждую составляющую своего определения Селье пояснил так: общий — потому что к стрессу приводят факторы, которые, воздействуя на разные области организма, в итоге способны вызвать общую системную защиту; адаптационный — потому что это явление как бы закрепляется, приобретает характер привычки; синдром — потому что его отдельные проявления частично взаимозависимы [13]. Позже (в 1931-1932 г.) он назвал стресс неспецифической реакцией организма на любые раздражители. Представление о стрессе касается всех людей, больных и здоровых, преуспевающих и неудачливых, и всех сторон жизни. «Удалось показать, что стресс представляет собой скорость изнашивания человеческого организма, сопровождает любую жизнедеятельность и соответствует в определённом смысле интенсивности жизни. Он увеличивается при нервном напряжении, телесных повреждениях, инфекциях, мышечной работе или любой другой напряженной деятельности и связан с неспецифическим защитным механизмом, увеличивающим сопротивляемость к стрессовым факторам, или „стрессорам“. Важной частью этого защитного механизма является повышенное выделение гипофизом так называемого адренокортикотропного гормона (АКТГ), который, в свою очередь, стимулирует выработку кортикоидов корой надпочечников.
Cиндром стресса, или, иначе, общий адаптационный синдром (ОАС), проходит три стадии:
- „реакция тревоги“, во время которой мобилизуются защитные силы. Её биологический смысл заключается в том, что организм в кратчайшие сроки должен получить дополнительную, „аварийную“ энергию для того, чтобы максимально обеспечить условия для быстрого спасения от грозящей беды или даже гибели. Да, для организма это, безусловно, реакция энергозатратная — что для него в перспективе, конечно, плохо, — но иного выхода в данный момент нет. Ведь речь идет о спасении в целом.
- „стадия устойчивости“ отражающая полную адаптацию к стрессору. Крайне важно и то, что на этой стадии общие энергозатраты организма становятся меньше, чем на первой стадии: организм частично уже приспособился жить под давлением стрессирующего фактора — как бы отследил его. Если стрессирующий фактор действует сильно и длительно, то постепенно развивается следующая, третья стадия.
- „стадия истощения“, которая неумолимо наступает, если стрессор оказывается достаточно силён и действует достаточно долгое время, поскольку „адаптационная энергия“, или приспособляемость живого вещества всегда конечна» [14]. Результат на этой стадии — устойчивость организма к внешним воздействиям резко снижается. То есть база для возможного развития патологии на клиническом уровне уже создана.
По мере формирования устойчивой адаптации нарушения гомеостаза, составляющие стимул стресс-симптома постепенно исчезают, как и сам стресс-симптом, сыграв свою важную роль в становлении адаптации. Это состояние между стрессом (агрессией) и адаптацией служит доказательством того, что стресс сложился в процессе эволюции как необходимое неспецифическое звено более сложного целостного механизма адаптации. Полная свобода от стресса, как считал Селье, означала бы смерть. В 1982 году Селье определил стресс как совокупность стереотипных филогенетических запрограммированных реакций организма, которые вызываются любыми сильными, сверхсильными, экстремальными воздействиями и сопровождаются перестройкой адаптивных сил организма.[15].
По признанию Г. Селье, он заимствовал у И. П. Павлова очень многое. То, что Павлов трактовал с точки зрения нервной системы, он перевел на язык и термины гуморальной системы. Павлов указывал, что организм как целое непрерывно адаптируется к окружающей среде. Эту идею целостности и адаптации Селье почерпнул у Павлова, и это было положено в основу теории стресса. То, что внешние воздействия приводят только к расходу и исчерпанию адаптационных возможностей, в течение многих лет было убеждением Г. Селье, которое он лаконично формулировал как «wear and tear». Это убеждение основывалось на том, что он использовал преимущественно сильные патогенные воздействия.
Концепции о существовании генетически предопределённого потенциала роста берут начало от А. Вейсмана, который считал, что зигота обладает способностью осуществлять определённое и ограниченное число делений клеток, после чего развивающийся организма обрекается на старение и смерть. Немецкий физиолог и гигиенист Макс Рубнер (М. Rubner) (1854—1932) в связи с представлениями Вейсмана обосновал понятие о генетически предопределённом энергетическом фонде, который у всех видов млекопитающих, за исключением человека почти одинаков. Концепцию Рубнера и вытекающее из неё представление об онтогенезе, как о раскручивании пружины в заведенных часах, и развил «Селье в учении о стрессе и о предопределённом фонде „адаптивной энергии“, сравнивая его с банковым вложением, которое можно расходовать, но нельзя увеличивать». Во всех этих теориях речь идет о постепенном исчерпании некоего исходного потенциала (жизненной субстанции), приводящих живые организмы к состоянию равновесия. Развивая эти представления Ричард Перл пришёл к мысли, что старение и смерть — „расплата за преимущества“ какие дают специализация и дифференциация клеток у многоклеточных организмов, в отличие от практически бессмертных одноклеточных этими качествами не обладающими.[16].
«Стресс есть неспецифический ответ организма на любое предъявление ему требования. … С точки зрения стрессовой реакции не имеет значения, приятна или неприятна ситуация, с которой мы столкнулись. Имеет значение лишь интенсивность потребности в перестройке или в адаптации.» [17]
Измерение адаптированности
В результате обработки многолетних наблюдений при сравнительном анализе популяций и групп, находящихся в различных экологических условиях (на Крайнем Севере и в средних широтах Сибири), А. Н. Горбанем и Е. И. Смирновой под руководством ак. АМН СССР К. Р. Седова получен вывод: наибольшую информацию о степени адаптированности популяции к экстремальным или просто изменившимся условиям несут корреляции между физиологическими параметрами.
В типичной ситуации при увеличении адаптационной нагрузки уровень корреляций повышается, а в результате успешной адаптации — снижается. Показано, что этот эффект группового стресса связан с организацией системы факторов, действующих на группу.
Если система факторов организована в соответствии с принципом Либиха (монолимитирование) то с ростом адаптационного напряжения размерность облака точек, представляющего группу в пространстве физиологических параметров, снижается, растет сумма абсолютных величин коэффициентов корреляции и, в то же время, растет разброс этого облака — возрастают дисперсии.
Если происходит взаимное усиление давления различных факторов (синергичные системы), то, напротив, с ростом адаптационного напряжения размерность облака точек, представляющего группу в пространстве физиологических параметров, растет, падает сумма коэффициентов корреляции. Дисперсии в то же время возрастают — так же, как и в предыдущем случае.
Литература
- Агаджанян Н. А., Труханов А. И., Шендеров Б. А. Этюды об адаптации и путях сохранения здоровья. — М.: изд-во «Сирин», — 2002. — 156 с.
- Ader Robert, Cohen Nicolas, Felten David, Psychoneuroimmunology: interactions between the nervous system and the immune system, Lancet 1995; 345:99-103
- Акофф Р., Эмерли Ф. О целеустремленных системах. — М.: Советское радио, 1974. — 272 с.
- Анохин П. К. Очерки по физиологии функциональных систем. — М.: Медицина, −1975.- 477 с.
- Анохин П. К.Узловые вопросы теории функциональной системы. — М.: Наука, −1980. — 197 с.
- Аршавский И. А.Физиологические механизмы и закономерности индивидуального развития. М.: Наука, — 1982. 270 с.
- Аршавский И. А. Некоторые методологические и теоретические аспекты анализа закономерностей индивидуального развития организмов//Вопросы философии. — № 11. — 1986. — С. 95-104.
- Бернар Кл. Лекции по экспериментальной патологии // Пер. Д. Е. Жуковского. М.; Л., 1937, с. 312—374.
- Бородюк Н. Р. Адаптация. Новое в приспособлении к окружающей среде. М.: «Глобус», 1998. — 88 с.
- Воробьёв Р. И. Медицина по Дарвину// Химия и жизнь. — № 2. — 2001. — с.44-48.
- Гартман Франц Жизнь Парацельса и сущность его учения/Пер. с англ. — М.: Алетейа, 2002. — 272с
- Дербенёва Л. М. Интимная жизнь стресса// Химия и жизнь. — № 7. — 1999 г. — С.54-57.
- Калайков И. Цивилизация и адаптация, М.: «Прогресс», 1984.
- Кассиль Г. Н. Внутренняя среда организма М.: Наука, 1983. — 227 с.
- Козинец Г. И., Погорелов В. М. Консерватизм биологических процессов — стабильность кроветворения Клиническая лабораторная диагностика № 12, 1988.-С.21-32.
- Малов Ю. С. Адаптация и здоровье. Клиническая медицина № 12. — 2001.- С. 61-63.
- Меерсон Ф. З., Пшенникова М. Г. Адаптация к стрессовым ситуациям и физическим нагрузкам. — М.: Медицина, 1988. — 256 с.
- Монтень Мишель, Опыты. Книга третья. Изд-во «Наука». — Москва. — 1981. — 535 с.
- Сапов И. А., Новиков В. С. Неспецифические механизмы адаптации человека. — Л.: Наука, 1984. — 146 с.
- Светуньков С. Г. Количественные методы прогнозирования эволюционных составляющих экономической динамики. — Ульяновск: Изд-во Ульяновского государственного университета, 1999. —177 с.
- Северцов А. С. Теория эволюции. — М.: Гуманитар. изд. центр ВЛАДОС, 2005. — 380с
- Скулачев В. П. Эволюция биологических механизмов запасания энергии // Соросовский Образовательный Журнал. 1997. № 5. С. 11-19.
- Урманцев Ю. А. Природа адаптации (системная экспликация)//«Вопросы философии». — 1998. — № 12.
- Философские проблемы адаптации Под ред. Г. И. Царегородцева. М., «Мысль», 1975 277 с.
- Hensel H. Grundbegriffe und neuere Aspekte der physiologischen Adaptation,
См. также
Примечания
- ↑ (Ю. А. Урманцев — 1998 г.)
- ↑ Акофф Р., Эмерли Ф. — 1974 г.
- ↑ (Hensel H. — 1974 г.)
- ↑ (Пилат Т. Л., Иванов А. А. — 2002 г.)
- ↑ (Малов Ю. С.- 2001 г.)
- ↑ (цит. по С. 621. — Монтень М. — 1981 г.)
- ↑ (цит. по С.57. — Ярошевский М. Г. — 1995 г.)
- ↑ (цит. по С.57.Ярошевский М. Г. — 1995 г.)
- ↑ (цит. по С.22. — «Химия и жизнь». — № 3. — 1990г)
- ↑ (цит. по С.13 Залманов А. С. — 1991 г.)
- ↑ (цит. по С. 8. — Агаджанян Н. А., Труханов А. И., Шендеров Б. А. — 2002 г.)
- ↑ (цит. по С.68. — Селье Г. — 1987 г.)
- ↑ (Дербенёва Л. М. — 1999 г.)
- ↑ (цит. по С.71-72 Селье Г. — 1987 г.)
- ↑ (Никонов В. В. — 2002 г.)
- ↑ цит. по С.15. Аршавский И. А. −1982., С. 95 Аршавский И. А.- 1986 г.
- ↑ Селье Г., Стресс жизни.
Адаптация (биология) — это… Что такое Адаптация (биология)?
У этого термина существуют и другие значения, см. Адаптация.Биологическая адаптация (от лат. adaptatio — приспособление) — приспособление организма к внешним условиям в процессе эволюции, включая морфофизиологическую и поведенческую составляющие. Адаптация может обеспечивать выживаемость в условиях конкретного местообитания, устойчивость к воздействию факторов абиотического и биологического характера, а также успех в конкуренции с другими видами, популяциями, особями. Каждый вид имеет собственную способность к адаптации, ограниченную физиологией (индивидуальная адаптация), пределами проявления материнского эффекта и модификаций, эпигенетическим разнообразием, внутривидовой изменчивостью, мутационными возможностями, коадаптационными характеристиками внутренних органов и другими видовыми особенностями.
Приспособленность живых существ к естественным условиям внешней среды была осознана людьми ещё в античные времена. Вплоть до середины XIX века это объяснялось изначальной целесообразностью природы. В теории эволюции Чарльза Дарвина было предложено научное объяснение адаптационного процесса на основе естественного отбора.
Адаптации видов в рамках одного биоценоза зачастую тесно связаны друг с другом (одним из наиболее поразительных примеров межвидовой коадаптации является жёсткая привязка строения органов некоторых видов цветковых растений и насекомых друг к другу с целью опыления и питания). Если адаптационный процесс у какого-либо вида не находится в равновесном состоянии, то эволюционировать может весь биоценоз (иногда — с негативными последствиями) даже в стабильных условиях окружающей среды.
Проблема определения адаптации
Философ Ю. Урманцев указывает на то, что по объёму и содержанию «адаптация» — понятие не только междисциплинарное, но и весьма сложное. Он обращает внимание на недостатки определений, предлагаемые энциклопедиями, а именно [1]:
- тавтологичность и отсутствие указаний на существенные и специфические признаки, которые бы выделяли именно адаптацию ;
- охватывание определением «адаптация», как правило, только одного (обычно медико-биологического) вида адаптации из, во много раз большего, числа существующих;
- вызванное этим отсутствие корректного определения вообще, а тем самым и методологических указаний о возможных путях развития учения об адаптации.
Процесс развития жизни на земле предполагает наличие адаптации у организмов. Начинается эта адаптация с самых примитивных видов — приспособления к окружающей среде и к существующим условиям. Возникновение и выживание организмов возможно только при соответствии организмов окружающей среде. Выживают те организмы, которые вырабатывают лучшие формы своего сохранения. Их развитие, переход организмов на более высокую ступень обусловлены необходимостью адаптации. Таким образом, эволюция и адаптация суть процессы, неотделимые друг от друга.
Под адаптацией можно понимать так же способность любой системы получать новую информацию для приближения своего поведения и структуры к оптимальным. Системы адаптивны, если при изменении в их окружении или внутреннем состоянии, снижающем их эффективность в выполнении своих функций, они реагируют или откликаются, изменяя свое собственное состояние или состояние окружающей среды так, чтобы их эффективность увеличилась [2]. Термин адаптация можно рассматривать в трех аспектах:
- адаптация как свойство системы приспосабливаться к возможным изменениям функционирования — система адаптации;
- адаптация как сам процесс приспособления адаптивной системы — собственно, адаптация;
- адаптация как метод, основанный на обработке поступающей информации и приспособленный для достижения некоторого критерия оптимизации — адаптационные алгоритмы.
В более широком смысле, адаптациями в биологии называют возникновение и развитие определенных, конкретных морфофизиологических свойств, значения которых для организма связаны с теми или иными общими или частными условиями его абиотической и биотической среды.
Адаптация, как адаптационный ответ
Адаптация, как адаптационный ответ, может осуществляться на различных уровнях:
- на уровне клетки в виде функциональных или морфологических изменений;
- на уровне органа или группы клеток, имеющих одинаковую функцию;
- на уровне организма как морфологического так и функционального целого, представляющего собой совокупность всех физиологических функций, направленных на сохранение витальных функций и самой жизни.
С учетом этого H. Hensel выделяет различные уровни адаптационных процессов [3]:
- привыкание — начальный процесс адаптации под влиянием кратковременного воздействия стрессора,
- функциональную адаптацию — продолжительное состояние, возникающее под влиянием определенных раздражителей, приводящих к физиологическим изменениям гомеостаза человека,
- трофо-пластическую адаптацию — дальнейшая ступень адаптационных процессов, не принадлежит к терапевтической области реабилитационной медицины, так как при ней наступают морфологические изменения органов и систем человеческого организма.
Главное содержание адаптации, по мнению Т. Пилат, — это внутренние процессы в системе, которые обеспечивают сохранение её внешних функций по отношению к среде. Если структура системы обеспечивает ей нормальное функционирование в данных условиях среды, то такую систему следует считать адаптированной к этим условиям. На этой стадии устанавливается динамическое равновесие, при котором происходит изменение физиологических показателей в границах нормы. Организм клинически здорового человека с высокими адаптивными способностями обладает значительным потенциалом саморегуляции и самовосстановления. Для поддержания здоровья ему достаточно вести здоровый образ жизни, соблюдать принципы сбалансированного питания [4].
При этом, по мнению Ю. Малова, здоровье или норма — состояние относительно стабильное, но оно не может сохраняться в течение всей жизни, ибо в природе не существует абсолютно адаптированных к данной среде индивидов. Длительное пребывание в одних и тех же условиях всё же рано или поздно приведет к развитию какой-либо болезни, которая является механизмом элиминации индивида [5].
История представлений об адаптации в биологии и медицине
«Биологическая адаптивность и есть жизнь». Ганс Селье
В философских предпосылках
В античности Фалес Милетский, Парменид, Демокрит, Гиппократ, Платон, Аристотель, Плутарх, Гален, Флавий Ренат Вегеций, а также позже Авиценна в том или ином виде замечали и указывали, что заболевания, поведение и некоторые другие виды психической деятельности зависят от внешних факторов, внешней среды окружающей человека.
Парацельс — считал, что все органические функции вызваны действием единого универсального «жизненного принципа». Если деятельность «жизненного принципа» внутри формы происходит нормальным и регулярным образом, который не нарушают никакие препятствия (в том числе и внешние), такое состояние называется здоровьем. Если же его деятельность по какой-либо причине нарушается или если он действует ненормально или нерегулярно, такое состояние называется болезнью.
Мишель Монтень писал: «Мы знаем по опыту, что на нас оказывает влияние воздух, климат, земля того места, где мы родились; причем они влияют не только на цвет нашей кожи, на наш рост, телосложение и осанку, но и на наши душевные качества. Люди меняют свой нрав, если их переселить в другое место, совершенно так же, как и деревья» [6].
В теориях эволюции
Основная статья: Адаптация (теории эволюции)
Тем не менее, научное обоснование идей адаптации разрабатывалось медленнее, чем философские воззрения и эмпирические знания на этот счёт и кардинально не менялось практически до 1809 года, когда французский естествоиспытатель Жан Батист Ламарк создал первое целостное учение об эволюции живой природы, основные идеи которого были изложены в «Философии зоологии».
На основе законов Ламарка сложилось направление, обычно называемое ламаркизмом или неоламаркизмом, а более точно эктогенезом — эктос — внешний (греч.), эктогенез — эволюция под действием внешних сил. В рамках этого направления эволюция рассматривалась как адаптивный процесс, основанный на всеобщем свойстве живых существ — «наследовании благоприобретённых свойств» (признаков). Следовательно, приспособленность рассматривалась как причина, а не результат эволюции. Единицей эволюции в рамках эктогенеза, является потомство родителей, которые передали своим потомкам результаты «упражнения или неупражнения органов», то есть череда поколений.
Дальнейшее развитие вопросы влияния внешней среды на человека получили в трудах английского естествоиспытателя, создателя теории эволюции Чарльза Роберта Дарвина. В 1859 году он выпустил книгу «Происхождение видов путём естественного отбора или сохранение благоприятствуемых пород в борьбе за жизнь», в которой было показано, как развивались все виды живого, согласно выдвинутому им положению о «выживании наиболее приспособленных».
Дарвин рассматривал приспособление (адаптацию) лишь в качестве средства для выживания. Согласно дарвинизму, эволюция представляет собой процесс адаптации организмов к условиям окружающей среды, причем приспособленность организмов является результатом эволюции.
В физиологии
Основная статья: Адаптация (физиология)
Во многом идеи К. Бернара получили продолжение в работах физиолога Ивана Петровича Павлова и американского ученого Уолтера Брейдфорда Кеннона.
Уолтер Кеннон, ввел в 1929 году в физиологию понятие гомеостаз считая, что постоянство внутренней среды организма поддерживается благодаря симпатической нервной системе и гормонам.
Категория поведения как особой формы саморегуляции жизни складывалась у И. П. Павлова (по образцу учения К. Бернара о гомеостазе), то есть об основных константах (давлении, температуре и др.), которые удерживают эту систему в стабильном состоянии в крайне неустойчивой внутренней среде. И. П. Павлов применил эту идею к взаимоотношениям организма с ещё более неустойчивой, непредсказуемой средой — внешней. Открытые им условно-рефлекторные механизмы служили достижению «уравновешивания» организма с этой средой.
Павловская трактовка рефлекса также имела четко выраженную гомеостатическую направленность и во многом базировалась на биодетерминизме К. Бернара, но не ограничивалась идеями гомеостаза. Известный американский физиолог Ч. Брукс отмечал: «Понятие об условном рефлексе предполагает появление длительных адаптивных изменений и даже включенность вегетативной системы» [7].
Связь с условным рефлексом
Условный рефлекс в трактовке И. П. Павлова, в первую очередь, — реакция адаптивного характера. И. П. Павлов уже в 1903 году в своей программной речи «Экспериментальная психология и психопатология на животных», на Международном медицинском конгрессе в Мадриде, впервые оповестил об открытии условных рефлексов, представив их как воплощение общебиологического закона приспособления организма к среде. В этом феномене, подчеркивал он, нет «ничего… кроме точной связи элементов сложной системы между собой и всего их комплекса с окружающей обстановкой». Он говорил тогда же: «К сожалению, мы не имеем до сих пор чисто научного термина для обозначения этого основного принципа организма — внутренней и внешней уравновешенности его» [8]. Через несколько десятков лет У. Кеннон изобрел этот термин, назвав «уравновешенность» гомеостазом. «Как часть природы, каждый животный организм представляет собою сложную обособленную систему, внутренние силы которой каждый момент, покуда она существует, как таковая, уравновешиваются с внешними силами окружающей среды. Чем сложнее организм, тем тоньше, многочисленнее и разнообразнее элементы уравновешивания. Для этого служат анализаторы и механизмы как постоянных, так и временных связей, устанавливающие точнейшие соотношения между мельчайшими элементами внешнего мира и тончайшими реакциями животного организма. Таким образом, вся жизнь от простейших до сложнейших организмов, включая, конечно, и человека, есть длинный ряд все усложняющихся до высочайшей степени уравновешиваний внешней среды» [9].
И. П. Павлов и У. Кеннон, в своих работах переходили к поведению целостного организма, то есть к особому типу его взаимодействия со средой. Ведь и условные рефлексы, и эмоции страха и ярости, ставшие (наряду с болью и голодом) главным объектом направления кенноновских исследований, выступают в качестве реакций, которые носят особый интегральный характер. Они даны в системе «организм — среда» и не могут быть объяснены вне её. Это не значит, что великий принцип единства организма и среды действует в полную силу лишь с переходом к условнорефлекторным и эмоциональным регуляциям. Речь идет о различных формах реализации этого принципа. Он выступает на многих уровнях: молекулярном, энергетическом, химическом, а также на уровнях функционирования различных физиологических и психологических систем. В условном рефлексе как поведенческом акте задействован физиологический механизм (по И. П. Павлову — кора и ближайшая к ней подкорка), но он становится поведенческим только тогда, когда в нём представлены условия среды в виде различаемых мозгом внешних (средовых) раздражителей, играющих роль сигналов.
Множество операций, проведенных в кенноновской школе над животными, доказали, что внешне наблюдаемые признаки поведения, которые можно назвать эмоциональными, порождаются глубинными сдвигами в нейрогуморальных процессах. Эти сдвиги готовят организм к критическим ситуациям, которые требуют повышенной траты энергии, снятия усталости, предотвращения кровопотери и т. п. На одном из докладов о своих открытиях У. Кеннон сообщил, что благодаря выбрасываемому в кровь при сильных эмоциях адреналину в числе других его «мобилизационных» эффектов происходит увеличение в крови поступающего к мышцам сахара.
У. Кеннон в поисках нового языка для обсуждения эмоционального поведения нашёл его у И. П. Павлова. Поведение трактуется им в терминах рефлексов — безусловных и условных. Главная эмоция для У. Кеннона отныне не что иное, как безусловный рефлекс. Картина поведения таламического, лишенного высших нервных центров, животного свидетельствует, согласно У. Кеннону, в пользу положения о том, что эмоция в её внешнем выражении является сложным безусловным рефлексом. Кеннон писал: «Жизнь избегает грубых аритмичных, неуправляемых взрывов. Небольшие изменения, маленькие химические реакции при умеренной температуре дают организму сопротивляемость крепче стали и направляются с точностью и тонкостью, не свойственной технике термитов. В этом «великая мудрость организма» [10].
Саморегуляция
И. П. Павлов в общей форме выдвинул принцип саморегуляции при исследовании деятельности сердца и кровообращения, а позднее распространил его на деятельность организма в целом. В работе «Ответ физиолога психологам» (1951) он отметил: «Человек есть, конечно, система (грубо говоря, — машина), как и всякая другая в природе, подчиняющаяся неизбежным и единым для всей природы законам, но система, в горизонте нашего современного видения, единственная по высочайшему саморегулированию…». Помимо учения о высшей нервной деятельности, определившего «принцип нервизма» Павлов обратил внимание на ферменты назвав их «возбудителями жизни». По представлению И. П. Павлова (1949) «… животный организм как система существует среди окружающей природы только благодаря непрерывному уравновешиванию этой системы на падающие на нее из вне раздражения» [11].
Таким образом, английская мысль внесла идею адаптации к среде внешней, как задаче, непрерывно решаемой организмом, французская мысль — идею саморегуляции процессов в этом организме, а русская — категорию поведения как особой формы саморегуляции жизни, что прекратило длившиеся многие годы прошлого столетия споры между «нервистами» и «гуморалистами», представляющие ныне лишь исторический интерес.
Адаптация и стресс
Ганс Селье вводит в медицинскую практику термин стресс. Он писал: «я впервые „наткнулся“ на идею стресса и общего адаптационного синдрома в 1925 году» [12]. Каждую составляющую своего определения Селье пояснил так: общий — потому что к стрессу приводят факторы, которые, воздействуя на разные области организма, в итоге способны вызвать общую системную защиту; адаптационный — потому что это явление как бы закрепляется, приобретает характер привычки; синдром — потому что его отдельные проявления частично взаимозависимы [13]. Позже (в 1931-1932 г.) он назвал стресс неспецифической реакцией организма на любые раздражители. Представление о стрессе касается всех людей, больных и здоровых, преуспевающих и неудачливых, и всех сторон жизни. «Удалось показать, что стресс представляет собой скорость изнашивания человеческого организма, сопровождает любую жизнедеятельность и соответствует в определённом смысле интенсивности жизни. Он увеличивается при нервном напряжении, телесных повреждениях, инфекциях, мышечной работе или любой другой напряженной деятельности и связан с неспецифическим защитным механизмом, увеличивающим сопротивляемость к стрессовым факторам, или „стрессорам“. Важной частью этого защитного механизма является повышенное выделение гипофизом так называемого адренокортикотропного гормона (АКТГ), который, в свою очередь, стимулирует выработку кортикоидов корой надпочечников.
Cиндром стресса, или, иначе, общий адаптационный синдром (ОАС), проходит три стадии:
- „реакция тревоги“, во время которой мобилизуются защитные силы. Её биологический смысл заключается в том, что организм в кратчайшие сроки должен получить дополнительную, „аварийную“ энергию для того, чтобы максимально обеспечить условия для быстрого спасения от грозящей беды или даже гибели. Да, для организма это, безусловно, реакция энергозатратная — что для него в перспективе, конечно, плохо, — но иного выхода в данный момент нет. Ведь речь идет о спасении в целом.
- „стадия устойчивости“ отражающая полную адаптацию к стрессору. Крайне важно и то, что на этой стадии общие энергозатраты организма становятся меньше, чем на первой стадии: организм частично уже приспособился жить под давлением стрессирующего фактора — как бы отследил его. Если стрессирующий фактор действует сильно и длительно, то постепенно развивается следующая, третья стадия.
- „стадия истощения“, которая неумолимо наступает, если стрессор оказывается достаточно силён и действует достаточно долгое время, поскольку „адаптационная энергия“, или приспособляемость живого вещества всегда конечна» [14]. Результат на этой стадии — устойчивость организма к внешним воздействиям резко снижается. То есть база для возможного развития патологии на клиническом уровне уже создана.
По мере формирования устойчивой адаптации нарушения гомеостаза, составляющие стимул стресс-симптома постепенно исчезают, как и сам стресс-симптом, сыграв свою важную роль в становлении адаптации. Это состояние между стрессом (агрессией) и адаптацией служит доказательством того, что стресс сложился в процессе эволюции как необходимое неспецифическое звено более сложного целостного механизма адаптации. Полная свобода от стресса, как считал Селье, означала бы смерть. В 1982 году Селье определил стресс как совокупность стереотипных филогенетических запрограммированных реакций организма, которые вызываются любыми сильными, сверхсильными, экстремальными воздействиями и сопровождаются перестройкой адаптивных сил организма.[15].
По признанию Г. Селье, он заимствовал у И. П. Павлова очень многое. То, что Павлов трактовал с точки зрения нервной системы, он перевел на язык и термины гуморальной системы. Павлов указывал, что организм как целое непрерывно адаптируется к окружающей среде. Эту идею целостности и адаптации Селье почерпнул у Павлова, и это было положено в основу теории стресса. То, что внешние воздействия приводят только к расходу и исчерпанию адаптационных возможностей, в течение многих лет было убеждением Г. Селье, которое он лаконично формулировал как «wear and tear». Это убеждение основывалось на том, что он использовал преимущественно сильные патогенные воздействия.
Концепции о существовании генетически предопределённого потенциала роста берут начало от А. Вейсмана, который считал, что зигота обладает способностью осуществлять определённое и ограниченное число делений клеток, после чего развивающийся организма обрекается на старение и смерть. Немецкий физиолог и гигиенист Макс Рубнер (М. Rubner) (1854—1932) в связи с представлениями Вейсмана обосновал понятие о генетически предопределённом энергетическом фонде, который у всех видов млекопитающих, за исключением человека почти одинаков. Концепцию Рубнера и вытекающее из неё представление об онтогенезе, как о раскручивании пружины в заведенных часах, и развил «Селье в учении о стрессе и о предопределённом фонде „адаптивной энергии“, сравнивая его с банковым вложением, которое можно расходовать, но нельзя увеличивать». Во всех этих теориях речь идет о постепенном исчерпании некоего исходного потенциала (жизненной субстанции), приводящих живые организмы к состоянию равновесия. Развивая эти представления Ричард Перл пришёл к мысли, что старение и смерть — „расплата за преимущества“ какие дают специализация и дифференциация клеток у многоклеточных организмов, в отличие от практически бессмертных одноклеточных этими качествами не обладающими.[16].
«Стресс есть неспецифический ответ организма на любое предъявление ему требования. … С точки зрения стрессовой реакции не имеет значения, приятна или неприятна ситуация, с которой мы столкнулись. Имеет значение лишь интенсивность потребности в перестройке или в адаптации.» [17]
Измерение адаптированности
В результате обработки многолетних наблюдений при сравнительном анализе популяций и групп, находящихся в различных экологических условиях (на Крайнем Севере и в средних широтах Сибири), А. Н. Горбанем и Е. И. Смирновой под руководством ак. АМН СССР К. Р. Седова получен вывод: наибольшую информацию о степени адаптированности популяции к экстремальным или просто изменившимся условиям несут корреляции между физиологическими параметрами.
В типичной ситуации при увеличении адаптационной нагрузки уровень корреляций повышается, а в результате успешной адаптации — снижается. Показано, что этот эффект группового стресса связан с организацией системы факторов, действующих на группу.
Если система факторов организована в соответствии с принципом Либиха (монолимитирование) то с ростом адаптационного напряжения размерность облака точек, представляющего группу в пространстве физиологических параметров, снижается, растет сумма абсолютных величин коэффициентов корреляции и, в то же время, растет разброс этого облака — возрастают дисперсии.
Если происходит взаимное усиление давления различных факторов (синергичные системы), то, напротив, с ростом адаптационного напряжения размерность облака точек, представляющего группу в пространстве физиологических параметров, растет, падает сумма коэффициентов корреляции. Дисперсии в то же время возрастают — так же, как и в предыдущем случае.
Литература
- Агаджанян Н. А., Труханов А. И., Шендеров Б. А. Этюды об адаптации и путях сохранения здоровья. — М.: изд-во «Сирин», — 2002. — 156 с.
- Ader Robert, Cohen Nicolas, Felten David, Psychoneuroimmunology: interactions between the nervous system and the immune system, Lancet 1995; 345:99-103
- Акофф Р., Эмерли Ф. О целеустремленных системах. — М.: Советское радио, 1974. — 272 с.
- Анохин П. К. Очерки по физиологии функциональных систем. — М.: Медицина, −1975.- 477 с.
- Анохин П. К.Узловые вопросы теории функциональной системы. — М.: Наука, −1980. — 197 с.
- Аршавский И. А.Физиологические механизмы и закономерности индивидуального развития. М.: Наука, — 1982. 270 с.
- Аршавский И. А. Некоторые методологические и теоретические аспекты анализа закономерностей индивидуального развития организмов//Вопросы философии. — № 11. — 1986. — С. 95-104.
- Бернар Кл. Лекции по экспериментальной патологии // Пер. Д. Е. Жуковского. М.; Л., 1937, с. 312—374.
- Бородюк Н. Р. Адаптация. Новое в приспособлении к окружающей среде. М.: «Глобус», 1998. — 88 с.
- Воробьёв Р. И. Медицина по Дарвину// Химия и жизнь. — № 2. — 2001. — с.44-48.
- Гартман Франц Жизнь Парацельса и сущность его учения/Пер. с англ. — М.: Алетейа, 2002. — 272с
- Дербенёва Л. М. Интимная жизнь стресса// Химия и жизнь. — № 7. — 1999 г. — С.54-57.
- Калайков И. Цивилизация и адаптация, М.: «Прогресс», 1984.
- Кассиль Г. Н. Внутренняя среда организма М.: Наука, 1983. — 227 с.
- Козинец Г. И., Погорелов В. М. Консерватизм биологических процессов — стабильность кроветворения Клиническая лабораторная диагностика № 12, 1988.-С.21-32.
- Малов Ю. С. Адаптация и здоровье. Клиническая медицина № 12. — 2001.- С. 61-63.
- Меерсон Ф. З., Пшенникова М. Г. Адаптация к стрессовым ситуациям и физическим нагрузкам. — М.: Медицина, 1988. — 256 с.
- Монтень Мишель, Опыты. Книга третья. Изд-во «Наука». — Москва. — 1981. — 535 с.
- Сапов И. А., Новиков В. С. Неспецифические механизмы адаптации человека. — Л.: Наука, 1984. — 146 с.
- Светуньков С. Г. Количественные методы прогнозирования эволюционных составляющих экономической динамики. — Ульяновск: Изд-во Ульяновского государственного университета, 1999. —177 с.
- Северцов А. С. Теория эволюции. — М.: Гуманитар. изд. центр ВЛАДОС, 2005. — 380с
- Скулачев В. П. Эволюция биологических механизмов запасания энергии // Соросовский Образовательный Журнал. 1997. № 5. С. 11-19.
- Урманцев Ю. А. Природа адаптации (системная экспликация)//«Вопросы философии». — 1998. — № 12.
- Философские проблемы адаптации Под ред. Г. И. Царегородцева. М., «Мысль», 1975 277 с.
- Hensel H. Grundbegriffe und neuere Aspekte der physiologischen Adaptation,
См. также
Примечания
- ↑ (Ю. А. Урманцев — 1998 г.)
- ↑ Акофф Р., Эмерли Ф. — 1974 г.
- ↑ (Hensel H. — 1974 г.)
- ↑ (Пилат Т. Л., Иванов А. А. — 2002 г.)
- ↑ (Малов Ю. С.- 2001 г.)
- ↑ (цит. по С. 621. — Монтень М. — 1981 г.)
- ↑ (цит. по С.57. — Ярошевский М. Г. — 1995 г.)
- ↑ (цит. по С.57.Ярошевский М. Г. — 1995 г.)
- ↑ (цит. по С.22. — «Химия и жизнь». — № 3. — 1990г)
- ↑ (цит. по С.13 Залманов А. С. — 1991 г.)
- ↑ (цит. по С. 8. — Агаджанян Н. А., Труханов А. И., Шендеров Б. А. — 2002 г.)
- ↑ (цит. по С.68. — Селье Г. — 1987 г.)
- ↑ (Дербенёва Л. М. — 1999 г.)
- ↑ (цит. по С.71-72 Селье Г. — 1987 г.)
- ↑ (Никонов В. В. — 2002 г.)
- ↑ цит. по С.15. Аршавский И. А. −1982., С. 95 Аршавский И. А.- 1986 г.
- ↑ Селье Г., Стресс жизни.
Адаптация (биология) — это… Что такое Адаптация (биология)?
У этого термина существуют и другие значения, см. Адаптация.Биологическая адаптация (от лат. adaptatio — приспособление) — приспособление организма к внешним условиям в процессе эволюции, включая морфофизиологическую и поведенческую составляющие. Адаптация может обеспечивать выживаемость в условиях конкретного местообитания, устойчивость к воздействию факторов абиотического и биологического характера, а также успех в конкуренции с другими видами, популяциями, особями. Каждый вид имеет собственную способность к адаптации, ограниченную физиологией (индивидуальная адаптация), пределами проявления материнского эффекта и модификаций, эпигенетическим разнообразием, внутривидовой изменчивостью, мутационными возможностями, коадаптационными характеристиками внутренних органов и другими видовыми особенностями.
Приспособленность живых существ к естественным условиям внешней среды была осознана людьми ещё в античные времена. Вплоть до середины XIX века это объяснялось изначальной целесообразностью природы. В теории эволюции Чарльза Дарвина было предложено научное объяснение адаптационного процесса на основе естественного отбора.
Адаптации видов в рамках одного биоценоза зачастую тесно связаны друг с другом (одним из наиболее поразительных примеров межвидовой коадаптации является жёсткая привязка строения органов некоторых видов цветковых растений и насекомых друг к другу с целью опыления и питания). Если адаптационный процесс у какого-либо вида не находится в равновесном состоянии, то эволюционировать может весь биоценоз (иногда — с негативными последствиями) даже в стабильных условиях окружающей среды.
Проблема определения адаптации
Философ Ю. Урманцев указывает на то, что по объёму и содержанию «адаптация» — понятие не только междисциплинарное, но и весьма сложное. Он обращает внимание на недостатки определений, предлагаемые энциклопедиями, а именно [1]:
- тавтологичность и отсутствие указаний на существенные и специфические признаки, которые бы выделяли именно адаптацию ;
- охватывание определением «адаптация», как правило, только одного (обычно медико-биологического) вида адаптации из, во много раз большего, числа существующих;
- вызванное этим отсутствие корректного определения вообще, а тем самым и методологических указаний о возможных путях развития учения об адаптации.
Процесс развития жизни на земле предполагает наличие адаптации у организмов. Начинается эта адаптация с самых примитивных видов — приспособления к окружающей среде и к существующим условиям. Возникновение и выживание организмов возможно только при соответствии организмов окружающей среде. Выживают те организмы, которые вырабатывают лучшие формы своего сохранения. Их развитие, переход организмов на более высокую ступень обусловлены необходимостью адаптации. Таким образом, эволюция и адаптация суть процессы, неотделимые друг от друга.
Под адаптацией можно понимать так же способность любой системы получать новую информацию для приближения своего поведения и структуры к оптимальным. Системы адаптивны, если при изменении в их окружении или внутреннем состоянии, снижающем их эффективность в выполнении своих функций, они реагируют или откликаются, изменяя свое собственное состояние или состояние окружающей среды так, чтобы их эффективность увеличилась [2]. Термин адаптация можно рассматривать в трех аспектах:
- адаптация как свойство системы приспосабливаться к возможным изменениям функционирования — система адаптации;
- адаптация как сам процесс приспособления адаптивной системы — собственно, адаптация;
- адаптация как метод, основанный на обработке поступающей информации и приспособленный для достижения некоторого критерия оптимизации — адаптационные алгоритмы.
В более широком смысле, адаптациями в биологии называют возникновение и развитие определенных, конкретных морфофизиологических свойств, значения которых для организма связаны с теми или иными общими или частными условиями его абиотической и биотической среды.
Адаптация, как адаптационный ответ
Адаптация, как адаптационный ответ, может осуществляться на различных уровнях:
- на уровне клетки в виде функциональных или морфологических изменений;
- на уровне органа или группы клеток, имеющих одинаковую функцию;
- на уровне организма как морфологического так и функционального целого, представляющего собой совокупность всех физиологических функций, направленных на сохранение витальных функций и самой жизни.
С учетом этого H. Hensel выделяет различные уровни адаптационных процессов [3]:
- привыкание — начальный процесс адаптации под влиянием кратковременного воздействия стрессора,
- функциональную адаптацию — продолжительное состояние, возникающее под влиянием определенных раздражителей, приводящих к физиологическим изменениям гомеостаза человека,
- трофо-пластическую адаптацию — дальнейшая ступень адаптационных процессов, не принадлежит к терапевтической области реабилитационной медицины, так как при ней наступают морфологические изменения органов и систем человеческого организма.
Главное содержание адаптации, по мнению Т. Пилат, — это внутренние процессы в системе, которые обеспечивают сохранение её внешних функций по отношению к среде. Если структура системы обеспечивает ей нормальное функционирование в данных условиях среды, то такую систему следует считать адаптированной к этим условиям. На этой стадии устанавливается динамическое равновесие, при котором происходит изменение физиологических показателей в границах нормы. Организм клинически здорового человека с высокими адаптивными способностями обладает значительным потенциалом саморегуляции и самовосстановления. Для поддержания здоровья ему достаточно вести здоровый образ жизни, соблюдать принципы сбалансированного питания [4].
При этом, по мнению Ю. Малова, здоровье или норма — состояние относительно стабильное, но оно не может сохраняться в течение всей жизни, ибо в природе не существует абсолютно адаптированных к данной среде индивидов. Длительное пребывание в одних и тех же условиях всё же рано или поздно приведет к развитию какой-либо болезни, которая является механизмом элиминации индивида [5].
История представлений об адаптации в биологии и медицине
«Биологическая адаптивность и есть жизнь». Ганс Селье
В философских предпосылках
В античности Фалес Милетский, Парменид, Демокрит, Гиппократ, Платон, Аристотель, Плутарх, Гален, Флавий Ренат Вегеций, а также позже Авиценна в том или ином виде замечали и указывали, что заболевания, поведение и некоторые другие виды психической деятельности зависят от внешних факторов, внешней среды окружающей человека.
Парацельс — считал, что все органические функции вызваны действием единого универсального «жизненного принципа». Если деятельность «жизненного принципа» внутри формы происходит нормальным и регулярным образом, который не нарушают никакие препятствия (в том числе и внешние), такое состояние называется здоровьем. Если же его деятельность по какой-либо причине нарушается или если он действует ненормально или нерегулярно, такое состояние называется болезнью.
Мишель Монтень писал: «Мы знаем по опыту, что на нас оказывает влияние воздух, климат, земля того места, где мы родились; причем они влияют не только на цвет нашей кожи, на наш рост, телосложение и осанку, но и на наши душевные качества. Люди меняют свой нрав, если их переселить в другое место, совершенно так же, как и деревья» [6].
В теориях эволюции
Основная статья: Адаптация (теории эволюции)
Тем не менее, научное обоснование идей адаптации разрабатывалось медленнее, чем философские воззрения и эмпирические знания на этот счёт и кардинально не менялось практически до 1809 года, когда французский естествоиспытатель Жан Батист Ламарк создал первое целостное учение об эволюции живой природы, основные идеи которого были изложены в «Философии зоологии».
На основе законов Ламарка сложилось направление, обычно называемое ламаркизмом или неоламаркизмом, а более точно эктогенезом — эктос — внешний (греч.), эктогенез — эволюция под действием внешних сил. В рамках этого направления эволюция рассматривалась как адаптивный процесс, основанный на всеобщем свойстве живых существ — «наследовании благоприобретённых свойств» (признаков). Следовательно, приспособленность рассматривалась как причина, а не результат эволюции. Единицей эволюции в рамках эктогенеза, является потомство родителей, которые передали своим потомкам результаты «упражнения или неупражнения органов», то есть череда поколений.
Дальнейшее развитие вопросы влияния внешней среды на человека получили в трудах английского естествоиспытателя, создателя теории эволюции Чарльза Роберта Дарвина. В 1859 году он выпустил книгу «Происхождение видов путём естественного отбора или сохранение благоприятствуемых пород в борьбе за жизнь», в которой было показано, как развивались все виды живого, согласно выдвинутому им положению о «выживании наиболее приспособленных».
Дарвин рассматривал приспособление (адаптацию) лишь в качестве средства для выживания. Согласно дарвинизму, эволюция представляет собой процесс адаптации организмов к условиям окружающей среды, причем приспособленность организмов является результатом эволюции.
В физиологии
Основная статья: Адаптация (физиология)
Во многом идеи К. Бернара получили продолжение в работах физиолога Ивана Петровича Павлова и американского ученого Уолтера Брейдфорда Кеннона.
Уолтер Кеннон, ввел в 1929 году в физиологию понятие гомеостаз считая, что постоянство внутренней среды организма поддерживается благодаря симпатической нервной системе и гормонам.
Категория поведения как особой формы саморегуляции жизни складывалась у И. П. Павлова (по образцу учения К. Бернара о гомеостазе), то есть об основных константах (давлении, температуре и др.), которые удерживают эту систему в стабильном состоянии в крайне неустойчивой внутренней среде. И. П. Павлов применил эту идею к взаимоотношениям организма с ещё более неустойчивой, непредсказуемой средой — внешней. Открытые им условно-рефлекторные механизмы служили достижению «уравновешивания» организма с этой средой.
Павловская трактовка рефлекса также имела четко выраженную гомеостатическую направленность и во многом базировалась на биодетерминизме К. Бернара, но не ограничивалась идеями гомеостаза. Известный американский физиолог Ч. Брукс отмечал: «Понятие об условном рефлексе предполагает появление длительных адаптивных изменений и даже включенность вегетативной системы» [7].
Связь с условным рефлексом
Условный рефлекс в трактовке И. П. Павлова, в первую очередь, — реакция адаптивного характера. И. П. Павлов уже в 1903 году в своей программной речи «Экспериментальная психология и психопатология на животных», на Международном медицинском конгрессе в Мадриде, впервые оповестил об открытии условных рефлексов, представив их как воплощение общебиологического закона приспособления организма к среде. В этом феномене, подчеркивал он, нет «ничего… кроме точной связи элементов сложной системы между собой и всего их комплекса с окружающей обстановкой». Он говорил тогда же: «К сожалению, мы не имеем до сих пор чисто научного термина для обозначения этого основного принципа организма — внутренней и внешней уравновешенности его» [8]. Через несколько десятков лет У. Кеннон изобрел этот термин, назвав «уравновешенность» гомеостазом. «Как часть природы, каждый животный организм представляет собою сложную обособленную систему, внутренние силы которой каждый момент, покуда она существует, как таковая, уравновешиваются с внешними силами окружающей среды. Чем сложнее организм, тем тоньше, многочисленнее и разнообразнее элементы уравновешивания. Для этого служат анализаторы и механизмы как постоянных, так и временных связей, устанавливающие точнейшие соотношения между мельчайшими элементами внешнего мира и тончайшими реакциями животного организма. Таким образом, вся жизнь от простейших до сложнейших организмов, включая, конечно, и человека, есть длинный ряд все усложняющихся до высочайшей степени уравновешиваний внешней среды» [9].
И. П. Павлов и У. Кеннон, в своих работах переходили к поведению целостного организма, то есть к особому типу его взаимодействия со средой. Ведь и условные рефлексы, и эмоции страха и ярости, ставшие (наряду с болью и голодом) главным объектом направления кенноновских исследований, выступают в качестве реакций, которые носят особый интегральный характер. Они даны в системе «организм — среда» и не могут быть объяснены вне её. Это не значит, что великий принцип единства организма и среды действует в полную силу лишь с переходом к условнорефлекторным и эмоциональным регуляциям. Речь идет о различных формах реализации этого принципа. Он выступает на многих уровнях: молекулярном, энергетическом, химическом, а также на уровнях функционирования различных физиологических и психологических систем. В условном рефлексе как поведенческом акте задействован физиологический механизм (по И. П. Павлову — кора и ближайшая к ней подкорка), но он становится поведенческим только тогда, когда в нём представлены условия среды в виде различаемых мозгом внешних (средовых) раздражителей, играющих роль сигналов.
Множество операций, проведенных в кенноновской школе над животными, доказали, что внешне наблюдаемые признаки поведения, которые можно назвать эмоциональными, порождаются глубинными сдвигами в нейрогуморальных процессах. Эти сдвиги готовят организм к критическим ситуациям, которые требуют повышенной траты энергии, снятия усталости, предотвращения кровопотери и т. п. На одном из докладов о своих открытиях У. Кеннон сообщил, что благодаря выбрасываемому в кровь при сильных эмоциях адреналину в числе других его «мобилизационных» эффектов происходит увеличение в крови поступающего к мышцам сахара.
У. Кеннон в поисках нового языка для обсуждения эмоционального поведения нашёл его у И. П. Павлова. Поведение трактуется им в терминах рефлексов — безусловных и условных. Главная эмоция для У. Кеннона отныне не что иное, как безусловный рефлекс. Картина поведения таламического, лишенного высших нервных центров, животного свидетельствует, согласно У. Кеннону, в пользу положения о том, что эмоция в её внешнем выражении является сложным безусловным рефлексом. Кеннон писал: «Жизнь избегает грубых аритмичных, неуправляемых взрывов. Небольшие изменения, маленькие химические реакции при умеренной температуре дают организму сопротивляемость крепче стали и направляются с точностью и тонкостью, не свойственной технике термитов. В этом «великая мудрость организма» [10].
Саморегуляция
И. П. Павлов в общей форме выдвинул принцип саморегуляции при исследовании деятельности сердца и кровообращения, а позднее распространил его на деятельность организма в целом. В работе «Ответ физиолога психологам» (1951) он отметил: «Человек есть, конечно, система (грубо говоря, — машина), как и всякая другая в природе, подчиняющаяся неизбежным и единым для всей природы законам, но система, в горизонте нашего современного видения, единственная по высочайшему саморегулированию…». Помимо учения о высшей нервной деятельности, определившего «принцип нервизма» Павлов обратил внимание на ферменты назвав их «возбудителями жизни». По представлению И. П. Павлова (1949) «… животный организм как система существует среди окружающей природы только благодаря непрерывному уравновешиванию этой системы на падающие на нее из вне раздражения» [11].
Таким образом, английская мысль внесла идею адаптации к среде внешней, как задаче, непрерывно решаемой организмом, французская мысль — идею саморегуляции процессов в этом организме, а русская — категорию поведения как особой формы саморегуляции жизни, что прекратило длившиеся многие годы прошлого столетия споры между «нервистами» и «гуморалистами», представляющие ныне лишь исторический интерес.
Адаптация и стресс
Ганс Селье вводит в медицинскую практику термин стресс. Он писал: «я впервые „наткнулся“ на идею стресса и общего адаптационного синдрома в 1925 году» [12]. Каждую составляющую своего определения Селье пояснил так: общий — потому что к стрессу приводят факторы, которые, воздействуя на разные области организма, в итоге способны вызвать общую системную защиту; адаптационный — потому что это явление как бы закрепляется, приобретает характер привычки; синдром — потому что его отдельные проявления частично взаимозависимы [13]. Позже (в 1931-1932 г.) он назвал стресс неспецифической реакцией организма на любые раздражители. Представление о стрессе касается всех людей, больных и здоровых, преуспевающих и неудачливых, и всех сторон жизни. «Удалось показать, что стресс представляет собой скорость изнашивания человеческого организма, сопровождает любую жизнедеятельность и соответствует в определённом смысле интенсивности жизни. Он увеличивается при нервном напряжении, телесных повреждениях, инфекциях, мышечной работе или любой другой напряженной деятельности и связан с неспецифическим защитным механизмом, увеличивающим сопротивляемость к стрессовым факторам, или „стрессорам“. Важной частью этого защитного механизма является повышенное выделение гипофизом так называемого адренокортикотропного гормона (АКТГ), который, в свою очередь, стимулирует выработку кортикоидов корой надпочечников.
Cиндром стресса, или, иначе, общий адаптационный синдром (ОАС), проходит три стадии:
- „реакция тревоги“, во время которой мобилизуются защитные силы. Её биологический смысл заключается в том, что организм в кратчайшие сроки должен получить дополнительную, „аварийную“ энергию для того, чтобы максимально обеспечить условия для быстрого спасения от грозящей беды или даже гибели. Да, для организма это, безусловно, реакция энергозатратная — что для него в перспективе, конечно, плохо, — но иного выхода в данный момент нет. Ведь речь идет о спасении в целом.
- „стадия устойчивости“ отражающая полную адаптацию к стрессору. Крайне важно и то, что на этой стадии общие энергозатраты организма становятся меньше, чем на первой стадии: организм частично уже приспособился жить под давлением стрессирующего фактора — как бы отследил его. Если стрессирующий фактор действует сильно и длительно, то постепенно развивается следующая, третья стадия.
- „стадия истощения“, которая неумолимо наступает, если стрессор оказывается достаточно силён и действует достаточно долгое время, поскольку „адаптационная энергия“, или приспособляемость живого вещества всегда конечна» [14]. Результат на этой стадии — устойчивость организма к внешним воздействиям резко снижается. То есть база для возможного развития патологии на клиническом уровне уже создана.
По мере формирования устойчивой адаптации нарушения гомеостаза, составляющие стимул стресс-симптома постепенно исчезают, как и сам стресс-симптом, сыграв свою важную роль в становлении адаптации. Это состояние между стрессом (агрессией) и адаптацией служит доказательством того, что стресс сложился в процессе эволюции как необходимое неспецифическое звено более сложного целостного механизма адаптации. Полная свобода от стресса, как считал Селье, означала бы смерть. В 1982 году Селье определил стресс как совокупность стереотипных филогенетических запрограммированных реакций организма, которые вызываются любыми сильными, сверхсильными, экстремальными воздействиями и сопровождаются перестройкой адаптивных сил организма.[15].
По признанию Г. Селье, он заимствовал у И. П. Павлова очень многое. То, что Павлов трактовал с точки зрения нервной системы, он перевел на язык и термины гуморальной системы. Павлов указывал, что организм как целое непрерывно адаптируется к окружающей среде. Эту идею целостности и адаптации Селье почерпнул у Павлова, и это было положено в основу теории стресса. То, что внешние воздействия приводят только к расходу и исчерпанию адаптационных возможностей, в течение многих лет было убеждением Г. Селье, которое он лаконично формулировал как «wear and tear». Это убеждение основывалось на том, что он использовал преимущественно сильные патогенные воздействия.
Концепции о существовании генетически предопределённого потенциала роста берут начало от А. Вейсмана, который считал, что зигота обладает способностью осуществлять определённое и ограниченное число делений клеток, после чего развивающийся организма обрекается на старение и смерть. Немецкий физиолог и гигиенист Макс Рубнер (М. Rubner) (1854—1932) в связи с представлениями Вейсмана обосновал понятие о генетически предопределённом энергетическом фонде, который у всех видов млекопитающих, за исключением человека почти одинаков. Концепцию Рубнера и вытекающее из неё представление об онтогенезе, как о раскручивании пружины в заведенных часах, и развил «Селье в учении о стрессе и о предопределённом фонде „адаптивной энергии“, сравнивая его с банковым вложением, которое можно расходовать, но нельзя увеличивать». Во всех этих теориях речь идет о постепенном исчерпании некоего исходного потенциала (жизненной субстанции), приводящих живые организмы к состоянию равновесия. Развивая эти представления Ричард Перл пришёл к мысли, что старение и смерть — „расплата за преимущества“ какие дают специализация и дифференциация клеток у многоклеточных организмов, в отличие от практически бессмертных одноклеточных этими качествами не обладающими.[16].
«Стресс есть неспецифический ответ организма на любое предъявление ему требования. … С точки зрения стрессовой реакции не имеет значения, приятна или неприятна ситуация, с которой мы столкнулись. Имеет значение лишь интенсивность потребности в перестройке или в адаптации.» [17]
Измерение адаптированности
В результате обработки многолетних наблюдений при сравнительном анализе популяций и групп, находящихся в различных экологических условиях (на Крайнем Севере и в средних широтах Сибири), А. Н. Горбанем и Е. И. Смирновой под руководством ак. АМН СССР К. Р. Седова получен вывод: наибольшую информацию о степени адаптированности популяции к экстремальным или просто изменившимся условиям несут корреляции между физиологическими параметрами.
В типичной ситуации при увеличении адаптационной нагрузки уровень корреляций повышается, а в результате успешной адаптации — снижается. Показано, что этот эффект группового стресса связан с организацией системы факторов, действующих на группу.
Если система факторов организована в соответствии с принципом Либиха (монолимитирование) то с ростом адаптационного напряжения размерность облака точек, представляющего группу в пространстве физиологических параметров, снижается, растет сумма абсолютных величин коэффициентов корреляции и, в то же время, растет разброс этого облака — возрастают дисперсии.
Если происходит взаимное усиление давления различных факторов (синергичные системы), то, напротив, с ростом адаптационного напряжения размерность облака точек, представляющего группу в пространстве физиологических параметров, растет, падает сумма коэффициентов корреляции. Дисперсии в то же время возрастают — так же, как и в предыдущем случае.
Литература
- Агаджанян Н. А., Труханов А. И., Шендеров Б. А. Этюды об адаптации и путях сохранения здоровья. — М.: изд-во «Сирин», — 2002. — 156 с.
- Ader Robert, Cohen Nicolas, Felten David, Psychoneuroimmunology: interactions between the nervous system and the immune system, Lancet 1995; 345:99-103
- Акофф Р., Эмерли Ф. О целеустремленных системах. — М.: Советское радио, 1974. — 272 с.
- Анохин П. К. Очерки по физиологии функциональных систем. — М.: Медицина, −1975.- 477 с.
- Анохин П. К.Узловые вопросы теории функциональной системы. — М.: Наука, −1980. — 197 с.
- Аршавский И. А.Физиологические механизмы и закономерности индивидуального развития. М.: Наука, — 1982. 270 с.
- Аршавский И. А. Некоторые методологические и теоретические аспекты анализа закономерностей индивидуального развития организмов//Вопросы философии. — № 11. — 1986. — С. 95-104.
- Бернар Кл. Лекции по экспериментальной патологии // Пер. Д. Е. Жуковского. М.; Л., 1937, с. 312—374.
- Бородюк Н. Р. Адаптация. Новое в приспособлении к окружающей среде. М.: «Глобус», 1998. — 88 с.
- Воробьёв Р. И. Медицина по Дарвину// Химия и жизнь. — № 2. — 2001. — с.44-48.
- Гартман Франц Жизнь Парацельса и сущность его учения/Пер. с англ. — М.: Алетейа, 2002. — 272с
- Дербенёва Л. М. Интимная жизнь стресса// Химия и жизнь. — № 7. — 1999 г. — С.54-57.
- Калайков И. Цивилизация и адаптация, М.: «Прогресс», 1984.
- Кассиль Г. Н. Внутренняя среда организма М.: Наука, 1983. — 227 с.
- Козинец Г. И., Погорелов В. М. Консерватизм биологических процессов — стабильность кроветворения Клиническая лабораторная диагностика № 12, 1988.-С.21-32.
- Малов Ю. С. Адаптация и здоровье. Клиническая медицина № 12. — 2001.- С. 61-63.
- Меерсон Ф. З., Пшенникова М. Г. Адаптация к стрессовым ситуациям и физическим нагрузкам. — М.: Медицина, 1988. — 256 с.
- Монтень Мишель, Опыты. Книга третья. Изд-во «Наука». — Москва. — 1981. — 535 с.
- Сапов И. А., Новиков В. С. Неспецифические механизмы адаптации человека. — Л.: Наука, 1984. — 146 с.
- Светуньков С. Г. Количественные методы прогнозирования эволюционных составляющих экономической динамики. — Ульяновск: Изд-во Ульяновского государственного университета, 1999. —177 с.
- Северцов А. С. Теория эволюции. — М.: Гуманитар. изд. центр ВЛАДОС, 2005. — 380с
- Скулачев В. П. Эволюция биологических механизмов запасания энергии // Соросовский Образовательный Журнал. 1997. № 5. С. 11-19.
- Урманцев Ю. А. Природа адаптации (системная экспликация)//«Вопросы философии». — 1998. — № 12.
- Философские проблемы адаптации Под ред. Г. И. Царегородцева. М., «Мысль», 1975 277 с.
- Hensel H. Grundbegriffe und neuere Aspekte der physiologischen Adaptation,
См. также
Примечания
- ↑ (Ю. А. Урманцев — 1998 г.)
- ↑ Акофф Р., Эмерли Ф. — 1974 г.
- ↑ (Hensel H. — 1974 г.)
- ↑ (Пилат Т. Л., Иванов А. А. — 2002 г.)
- ↑ (Малов Ю. С.- 2001 г.)
- ↑ (цит. по С. 621. — Монтень М. — 1981 г.)
- ↑ (цит. по С.57. — Ярошевский М. Г. — 1995 г.)
- ↑ (цит. по С.57.Ярошевский М. Г. — 1995 г.)
- ↑ (цит. по С.22. — «Химия и жизнь». — № 3. — 1990г)
- ↑ (цит. по С.13 Залманов А. С. — 1991 г.)
- ↑ (цит. по С. 8. — Агаджанян Н. А., Труханов А. И., Шендеров Б. А. — 2002 г.)
- ↑ (цит. по С.68. — Селье Г. — 1987 г.)
- ↑ (Дербенёва Л. М. — 1999 г.)
- ↑ (цит. по С.71-72 Селье Г. — 1987 г.)
- ↑ (Никонов В. В. — 2002 г.)
- ↑ цит. по С.15. Аршавский И. А. −1982., С. 95 Аршавский И. А.- 1986 г.
- ↑ Селье Г., Стресс жизни.
| 1. |
Обитатели различных сред
Сложность: лёгкое |
2 |
| 2. |
Фотопериод
Сложность: лёгкое |
1 |
| 3. |
Жизненные формы растений
Сложность: лёгкое |
1 |
| 4. |
Организм как среда
Сложность: среднее |
7 |
| 5. |
Адаптации организмов к водной среде
Сложность: среднее |
1 |
| 6. |
Факторы среды
Сложность: среднее |
1 |
| 7. |
Характеристика сред жизни
Сложность: среднее |
3 |
| 8. |
Определи среду обитания
Сложность: среднее |
1 |
| 9. |
Лимитирующие факторы
Сложность: среднее |
1 |
| 10. |
Условия среды
Сложность: сложное |
1 |
Адаптация человека к среде обитания
Адаптация человека к новой для него среде – сложный социально-биологический процесс, в основе которого лежит изменение систем и функций организма, а также привычного поведения. Под адаптацией человека понимаются приспособительные реакции его организма на изменяющиеся факторы среды. Адаптация проявляется на разных уровнях организации живой материи: от молекулярного до биоценотического. Адаптация развивается под воздействием трёх факторов: наследственность, изменчивость, естественный/искусственный отбор. Существует три основных пути приспособления организмов к среде обитания: активный путь, пассивный путь и избегание неблагоприятных воздействий.
Активный путь – усиление сопротивляемости, развитие регуляторных процессов, позволяющих осуществлять все жизненные функции организма, несмотря на отклонение фактора среды от оптимума. Например, поддержание постоянной температуры тела у теплокровных (птиц, млекопитающих, человека), оптимальной для протекания биохимических процессов в клетках.
Пассивный путь – подчинение жизненных функций организма изменению факторов среды. Например, перехлд при неблагоприятных условиях среды в состояние анабиоза (скрытой жизни), когда обмен веществ в организме практически полностью останавливается (зимний покой растений, сохранение семян и спор в почве, оцепенение насекомых, спячка позвоночных животных и т.д.).
Избегание неблагоприятных условий – выработка организмом таких жизненных циклов и поведения, которые позволяют избежать неблагоприятных воздействий. Например, сезонные миграции животных.
Обычно приспособление вида к среде проходит тем или иным сочетанием всех трёх возможных путей адаптации.
Адаптации можно разделить на три основных ипа: морфологические, физиологические, этологические.
Морфологические адаптации – изменения в строении организма(например, видоизменение листа в колючку у кактусов для снижения потерь воды, яркая окраска цветов для привлечения опылителей и др.). Морфологические адаптации у животных приводят к образованию определённых жизненных форм.
Физиологические адаптации – изменения в физиологии организма (например, способность верблюда обеспечивать организм влагой путём окисления запасов жира, наличие целлюлозоразрушающих ферментов у целлюлозоразрушающих бактерий и др.).
Этологические (поведенческие) адаптации – изменения в поведении (например, сезонные миграции млекопитающих и птиц, впадение в спячку в зимний период, брачные игры у птиц и млекопитающих в период размножения и др.). Этологические адаптации характерны для животных.
Живые организмы хорошо адаптированы к периодическим факторам. Непериодические факторы могут вызвать болезни и даже смерть живого организма. Человек использует это, применяя пестициды, антибиотики и другие непериодические факторы. Однако длительность их воздействия также может вызвать к ним адаптацию.
Среда обитания оказывает огромное воздействие на человека. В связи с этим все большую актуальность приобретает проблема адаптации человека к своей среде. В социальной экологии этой проблеме придается первостепенное значение. В то же время адаптация – это лишь начальный этап, на котором преобладают реактивные формы поведения человека. Человек не останавливается на этом этапе. Он проявляет физическую, интеллектуальную, нравственную, духовную активность, преобразует (в худшую или худшую сторону) свою среду.
Адаптация человека подразделяется на генотипическую и фенотипическую. Генотипическая адаптация: человек вне своего сознания может приспособиться к изменившимся условиям среды (перепадам температуры, вкусу пищи и т. д.), то есть, если механизмы адаптации заложены уже в генах. Под фенотипической адаптацией понимается включение сознания, своих личностных качеств человека, чтобы приспособится организму к новой среде, сохранить в новых условиях равновесие.
К основным видам адаптации относят физиологическую, адаптацию к деятельности, адаптацию к социуму. Остановимся на физиологической адаптации. Под физиологической адаптацией человека понимается процесс поддержания функционального состояния организма в целом, обеспечивая его сохранение, развитие, работоспособность, максимальную продолжительность жизни. Большое значение в физиологической адаптации придается акклимации и акклиматизации. Понятно, что жизнь человека на Крайнем Севере отличается от его жизни на экваторе, так как это разные климатические зоны. Причем южанин, прожив определенное время на севере, адаптируется к нему и может жить там постоянно и, наоборот. Акклимация – это начальный, срочный этап акклиматизации при изменении климато-географических условий. В некоторых случаях синонимом физиологической адаптации является акклиматизация, то есть приспособление растений, животных и человека к новым для них климатическим условиям. Физиологическая акклиматизация наступает когда у человека с помощью приспособительных реакций повышается работоспособность, улучшается самочувствие, которое может резко ухудшиться в период акклимации. При смене новых условий старыми организм может возвратиться к прежнему состоянию. Подобные изменения и называются акклиматизацией. Те же изменения, которые в процессе приспособления к новой среде перешли в генотип и передаются по наследству, называются адаптивными.
Адаптация организма к условиям проживания (городу, селу, другой местности). Среда обитания человека не ограничивается лишь климатическими условиями. Человек может жить в городе и в селе. Очень многие предпочитают мегаполис с его шумом, загрязненностью, бешенным ритмом жизни. Объективно жить в селе, где чистый воздух, спокойный размеренный ритм, более благоприятно для людей.
К этой же сфере адаптации относиться переезд, например, в другую страну. Одни быстро адаптируются, преодолевают языковый барьер, находят себе работу, другие – с большим трудом, третьи, внешне адаптировавшись, испытывают чувство, которое называется ностальгией.
Можно особо выделить адаптацию к деятельности. Различные виды деятельности человека предъявляют различные требования к личности (одни требуют усидчивости, исполнительности, пунктуальности, другие – быстроты реакции, умения самостоятельно принимать решения и т. п.). Однако с теми и другими видами деятельности человек может справиться достаточно успешно. Есть деятельность, которая противопоказана человеку, но он ее может выполнять, так как срабатывают механизмы адаптации, который называется выработкой индивидуального стиля деятельности.
Особо следует остановиться на адаптации к социуму, другим людям, коллективу. Человек может приспособиться к группе, усвоив ее нормы, правила поведения, ценности и т. д. В качестве механизмов адаптации здесь выступают внушаемость, толерантность, конформность как формы подчинительного поведения, а с другой – умение найти свое место, обрести лицо, проявить решительность.
Можно говорить об адаптации к духовным ценностям, к вещам, к состояниям, например, к стрессовым, и ко многому другому. В 1936 году канадский физиолог Ганс Селье опубликовал сообщение «Синдром, вызванный разными повреждающими элементами», в котором описал явление стресса – общей неспецифической реакции организма, направленной на мобилизацию его защитных сил при действии раздражающих факторов. В развитии стресса были выделены 3 стадии: 1. стадия тревоги, 2. стадия сопротивления, 3. стадия истощения. Г. Селье была сформулирована теория «Общего адаптационного синдрома» (ОАС) и адаптивных болезней, как следствия адаптационной реакции, согласно которой ОАС проявляется всякий раз, когда человек чувствует опасность для себя. Видимыми причинами стресса могут быть травмы, послеоперационные состояния и т. д., изменения абиотических и биотических факторов среды. В последние десятилетия значительно возросло число антропогенных факторов среды, обладающих высоким стрессогенным эффектом (химическое загрязнение, радиация, воздействие компьютеров при систематической работе с ними и т. д.). К стрессорным факторам среды следует отнести и негативные изменения в современном обществе: повышение плотности населения, изменение соотношения городского и сельского населения, рост безработицы, преступность.
Биологическая адаптация — обзор
Естественный и половой отбор
Идея о том, что люди, наряду со всеми другими организмами, являются продуктом долгой истории эволюции (в терминах Дарвина: «происхождение с модификацией»), во всех смыслах является и цели, правда (Coyne, 2009; Dawkins, 2009). Действительно, существование биологической эволюции было признано многими мыслителями еще до публикации знаменательной книги Дарвина (1859 г.) О происхождении видов . Однако в этих ранних описаниях эволюции отсутствовало правдоподобное описание механизмов , ответственных за эволюционные изменения.Любой взгляд на современный учебник по эволюции (например, Futuyma, 2009) обнаруживает большое количество взаимосвязанных теорий, которые в совокупности могут помочь объяснить характеристики организмов, изменения в популяциях с течением времени, географическое распределение видов, происхождение. новых видов и так далее. Однако центральным элементом современной эволюционной мысли является описание Дарвина ключевого механизма, ответственного за эволюционные изменения: естественного отбора.
Естественный отбор с учетом времени, необходимого для работы, является чрезвычайно мощным процессом, но его можно аккуратно резюмировать в терминах трех ключевых принципов: вариативность, дифференциальная пригодность и наследование.Во-первых, представители вида различаются по различным характеристикам, которыми они обладают. Например, некоторые импалы могут бегать быстрее других, некоторые обитающие в пустыне жабы могут лучше сохранять влагу, чем их собратья-жабы, а некоторые самцы павианов могут быть более доминирующими физически, чем другие. Во-вторых, некоторые из этих различий приведут к различиям в физической форме. Другими словами, некоторые представители вида могут иметь больше шансов выжить и размножаться в результате специфических характеристик, которыми они обладают: например, чем более быстроногие импалы с большей вероятностью будут уклоняться от хищников, тем больше будут жабы, сохраняющие влагу. с большей вероятностью выживут в продолжительной засухе, и более доминирующие павианы могут получить преимущественный доступ к пище и спариванию.В-третьих, что очень важно для эволюции, эти различия должны быть наследуемыми. Другими словами, они должны надежно передаваться от одного поколения к другому (чаще всего от родителей к потомству через общие гены, но см. Обсуждение ниже). Со временем, учитывая эти три процесса, характеристики популяции организмов будут меняться, поскольку более предпочтительные характеристики сохраняются за счет менее выгодных вариантов.
Таким образом, естественный отбор является основным процессом эволюционных изменений.Медленно отсеивая менее благоприятные черты и отдавая предпочтение тем, которые надежно повышают выживаемость и репродуктивный успех, характеристики организмов формируются таким образом, чтобы способствовать «соответствию» между организмами и окружающей средой. Эти характеристики известны как адаптации. Сложный нейронный механизм, лежащий в основе эхолокации у летучих мышей, способность хамелеонов изменять цвет своей кожи в зависимости от местных условий и двуногая походка современных людей — все это примеры биологических адаптаций: черт, которые обязаны своим существованием их историческому успеху в обеспечении выживания и жизни. репродуктивный успех.Здесь стоит отметить, что естественный отбор — не единственный фактор эволюционных изменений, и эволюционные биологи также признают важность случайных факторов (таких как «генетический дрейф») в формировании характеристик популяций (Futuyma, 2009). Более того, процесс естественного отбора не только порождает адаптации, но и приводит к побочным продуктам адаптации, наряду с остаточным биологическим «шумом» (Buss, Haselton, Shackelford, Bleske, & Wakefield, 1998; Travis & Reznick, 2009). ).Понятие побочных продуктов широко используется, но охватывает разнородный класс объектов. Знакомыми примерами являются звук, который издают сердце при биении (побочный продукт отбора для механизма перекачивания сердец), белый цвет костей (побочный продукт солей кальция, придающих костям твердость и жесткость) и человеческая способность к письму (отчасти побочный продукт эволюционировавших механизмов, лежащих в основе языка). Идея биологического «шума» просто улавливает идею о том, что вариация признаков, если она избирательно нейтральна, будет сохраняться в организмах, не влияя на биологическую приспособленность.
Проблема четкого разграничения адаптаций от неадаптаций в биологическом мире является важной. Не существует надежного метода выявления биологических адаптаций; однако характеристики, сформированные естественным отбором, часто демонстрируют особенности «особого дизайна», такие как функциональность, экономичность, эффективность, точность, специализация, надежность и адаптивная сложность (Andrews, Gangestad, & Matthews, 2002; Williams, 1966). Человеческий глаз, если использовать часто цитируемый пример, имеет все признаки биологической адаптации: несколько частей (сетчатка, хрусталик, роговица, радужная оболочка) скоординированы для эффективного и надежного выполнения единой функции (зрения) с очевидными эволюционными преимуществами. (зрение явно выгодно и много раз развивалось независимо).Эволюция человеческого глаза также иллюстрирует другой важный аспект биологической адаптации — хотя глаза могут быть специализированы для выполнения конкретной функциональной задачи, их конструкция не всегда может быть оптимальной. Поскольку эволюция, как отметил Джейкоб (1977), представляет собой процесс «возведения в порядок», в котором диапазон возможных «решений» адаптивных проблем ограничен тем, что было ранее (с точки зрения эволюции), мы должны ожидать, что этот «неоптимальный» дизайн, который несет на себе отпечаток эволюционной истории рассматриваемой характеристики.Например, поскольку глаз позвоночных изначально развился как устройство для обнаружения света (а не для создания изображений), у людей есть «слепое пятно», вызванное тем, как зрительный нерв расположен в сетчатке (Travis & Reznick, 2009).
Это означает, что в наших усилиях по выявлению адаптаций мы должны учитывать историю рассматриваемой характеристики. История также важна, потому что разные виды имеют общие предки. Например, люди и шимпанзе имеют общего предка, который жил около пяти-восьми миллионов лет назад; все приматы (включая людей) имеют общего предка, который жил примерно 63 млн лет назад; все позвоночные произошли от предка, жившего более 500 млн лет назад, и так далее (см. наглядное описание этой истории в Докинз, 2004 г.).Следовательно, чтобы дать четкое представление об адаптивной эволюции конкретных характеристик, нам необходимо установить, какие черты являются «наследственными» (они присутствовали у общего предка определенной эволюционной линии), а какие — «производными» (они уникальны для конкретной эволюционной линии и, следовательно, отражают процессы отбора, происходящие после отделения от общего предка). Эти соображения составляют часть так называемого «сравнительного метода» в эволюционной биологии для выявления адаптаций и влекут за собой подробный филогенетический (эволюционно-исторический) анализ конкретных характеристик (Griffiths, 1996; Harvey & Pagel, 1991).Детали этого подхода не должны здесь нас интересовать, но с практической точки зрения это означает, что при разработке гипотез относительно эволюции человеческих характеристик мы должны учитывать их «более глубокую» эволюционную историю, а исследования шимпанзе и других приматов, в частности, могут пролить свет на этот процесс, как мы проиллюстрируем в этой книге (см. Mitani, Call, Kappeler, Palombit, & Silk, 2012).
Концепция адаптации, как заметил Джордж Уильямс (1966), обременительна, и нам следует с осторожностью заявлять, что определенные черты являются адаптациями, при отсутствии соответствующих доступных доказательств.Этот вопрос может иметь особое значение при изучении человеческих характеристик и черт. Например, было много жарких споров о том, можно ли изнасилование человека рассматривать как биологическую адаптацию или является побочным продуктом других адаптаций (Thornhill & Palmer, 2000), и сохраняются разногласия относительно того, есть ли у людей особые приспособления к убийству ( Дантли и Басс, 2011; Даррант, 2009). Как мы обсудим в нашем обзоре эволюционной психологии, определение адаптаций — гораздо более простая задача, чем их идентификация, особенно когда они предполагаются в связи с психологическими и поведенческими чертами человека, а иногда эволюционные психологи были несколько чрезмерно нетерпеливы, чтобы объявить какие-то человеческие характеристики как адаптация при отсутствии продуманной оценки альтернативных гипотез.Однако разумная стратегия — разработать предварительные гипотезы относительно адаптивных функций конкретных характеристик и принять лучшее из текущих объяснений, признавая при этом, что это признание всегда может изменяться (Durrant & Haig, 2001).
Хотя довольно легко распознать, как скорость бега импал и механизмы сохранения влаги у обитающих в пустынях жаб способствуют выживанию, и, следовательно, они, вероятно, являются результатом естественного отбора, многие характеристики организмов, по-видимому, не предполагают столь очевидного утилитарные функции.Рассмотрим довольно удачно названную длиннохвостую птицу-вдову. Самцы этого вида размером с воробья имеют хвост длиной 2 м, который явно ограничивает способность птицы летать. Как признал Дарвин (1871), многие характеристики организмов не увеличивают выживаемость, а являются продуктами процесса, который он назвал половой отбор . Половой отбор, вероятно, лучше всего охарактеризовать как частный случай естественного отбора, который связан с выбором партнера и спариванием (Andersson, 1994; Clutton-Brock, 2004).Дарвин (1871) выделил два основных типа полового отбора. Первый тип относится к выбору партнера и называется интерсексуальным отбором. Короче говоря, можно выбрать конкретные характеристики, если представители одного пола отдают предпочтение определенным характеристикам, которыми обладает другой. Вероятно, например, что удлиненный хвост самца длиннохвостой птицы-вдовы является продуктом долгой истории самок, преимущественно спаривающихся с самцами, имеющими самый длинный хвост (Andersson, 1982).Другая форма полового отбора, известная как интрасексуальный отбор, связана с конкуренцией между представителями одного пола за доступ к спариванию с представителями другого пола. Эволюция мужского оружия у многих видов животных в виде клыков, рогов, больших клыков и т. возможности для спаривания и улучшение репродуктивной способности (Emlen, 2008).
Определение и примеры адаптации — Биологический онлайн-словарь
Адаптация
n., ˌÆdæpˈteɪʃən
Процесс или состояние адаптации или изменения, чтобы стать более подходящим для окружающей среды; характеристика как результат процесса
Определение адаптации
В биологии и экологии, адаптация относится к процессу адаптации поведения, физиологии или структуры, чтобы стать более приспособленными к окружающей среде. Это также может быть определено как состояние, в котором биологическая популяция претерпевает корректировки или изменения.Это также может относиться к признаку , который сделал вид лучше приспособленным к окружающей среде. Эта черта называется адаптивной чертой .
Информация
Термин адаптация произошел от латинского «адаптироваться», означающего «Я подхожу» или «Я приспосабливаюсь к». Это обычное слово в экологии и эволюционной биологии. Организм имеет тенденцию претерпевать изменения, чтобы стать более приспособленным к окружающей среде. Адаптация отличается от акклиматизации. Оба условия касаются изменений.Однако акклиматизация — это физиологическая адаптация к новым условиям, но она не влечет за собой увеличения видового разнообразия, как адаптация. Чтобы признак можно было рассматривать как адаптацию, он должен быть наследуемым, функциональным и повышать приспособленность. (Ссылка 1)
Согласно теории эволюции путем естественного отбора Чарльза Дарвина , организмы приспосабливаются к окружающей среде, чтобы они могли сохраняться и передавать свои гены следующему поколению. В экосистеме несколько видов совместно адаптируются, и при этом они также совместно эволюционируют.Возьмем, к примеру, симбионтов в ассоциации лишайников. Компоненты водорослей и грибов имеют тенденцию эволюционировать вместе, поскольку отсутствие одного из них означает смерть. Зависимость от существования друг друга означает, что они должны вместе адаптироваться, вместе выживать.
Адаптации необходимы для выживания вида. Адаптивные черты, которые вид приобретет с течением времени, могут быть структурными (то есть физическими адаптивными чертами), поведенческими (например, вокализациями, ритуалами ухаживания, гнездования и спаривания) или физиологическими (т.е.грамм. развитие устойчивости к болезням или к токсичным химическим веществам). (Ссылка 1, 2)
Другие определения
В неврологии адаптация означает снижение частоты возбуждения нейрона. В офтальмологии это способность глаза приспосабливаться к различной интенсивности света путем регулирования зрачком количества света, попадающего в глаз через зрачок.
ПРОЧИТАЙТЕ: Учебное пособие по адаптации
Посмотрите это видео об адаптации у животных.
Это видео об адаптации на заводах
Ссылки
- Определение адаптации.(2020). Berkeley.Edu. https://evolution.berkeley.edu/evolibrary/article/0_0_0/misconcep_06
- Адаптация. (2020). Berkeley.Edu. https://evolution.berkeley.edu/evolibrary/article/evo_31
© BiologyOnline. Контент предоставлен и модерируется редакторами BiologyOnline.
СледующийОпределение и признание биологической адаптации в JSTOR
АбстрактныйБиологическая адаптация — это свойство фенотипических особенностей организмов по отношению к требованиям отбора окружающей среды.Адаптивные свойства — это те свойства, которые обладают свойствами формы и функции, которые позволяют организму успешно поддерживать синергизм между биологической ролью этого свойства и заявленной силой отбора. Степень положительности адаптации можно измерить с помощью количества энергии, необходимого для поддержания синергизма, при этом меньшая энергия указывает на лучшую адаптацию. Адаптация не применяется к точному совпадению между различными элементами или между формой и функцией элемента или к взаимным взаимодействиям между элементами структурной сети.Состояние адаптации не зависит от процесса адаптации, поскольку адаптации не обязательно должны развиваться под контролем сил отбора, к которым они теперь адаптированы. Отбор здесь рассматривается как требования, предъявляемые к организму окружающей средой, с которой организм должен справляться, чтобы продолжать выживать как индивидуум. Адаптированные характеристики оценивались с использованием трех методов — сравнительного, коррелятивного и синтетического, из которых действителен только последний. Синтетический метод требует изучения форм и функций признака в лаборатории, а также биологических ролей признака и требований к отбору окружающей среды в полевых условиях.Предполагается, что лучший подход к изучению адаптаций — это командные усилия с постоянной обратной связью между лабораторным и полевым этапами исследования. Настоятельно рекомендуется уделять внимание развитию экологической морфологии, чтобы дополнить функциональную морфологию и обеспечить необходимую основу для правильного выяснения биологических адаптаций.
Информация об издателеOxford University Press — это отделение Оксфордского университета.Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP — крупнейшая в мире университетская пресса с самым широким глобальным присутствием. В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5 500 сотрудников по всему миру. Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
Адаптации | BioNinja
Понимание:
• Адаптации — это характеристики, которые делают человека приспособленным к окружающей среде и образу жизни
• Лица, передающие характеристики своему потомству
Адаптации — это особенности организмов, которые помогают им выживать, позволяя им лучше адаптироваться к окружающей среде
Эти адаптации можно классифицировать по-разному:
- Структурные: Физические различия в биологической структуре (например,грамм. длина шеи жирафа)
- Поведенческие: Различия в образцах активности (например, опоссумы симулируют смерть при угрозе)
- Физиологические: Вариации в обнаружении и реакции жизненно важных органов (например, гомеотермия, цветовосприятие)
- Биохимические : Различия в молекулярном составе клеток и функциях ферментов (например, группы крови, толерантность к лактозе)
- Развитие: Переменные изменения, которые происходят на протяжении жизни организма (например,грамм. закономерности старения / старения)
Биологические адаптации имеют генетическую основу (т.е. кодируются генами) и могут передаваться потомству при воспроизводстве родителей
- Организмов с полезными адаптациями будет более вероятно, что выживут долго достаточно, чтобы воспроизводить и передавать эти гены. возможность естественного отбора
Адаптации ехидныЧто такое адаптация? — Определение, причины и типы адаптации
Теория адаптации
Теория адаптации также называется теорией выживания наиболее приспособленных, которая представляет собой способность организма адаптироваться к окружающей среде и приспосабливаться со временем.Ученый, который выступил с его теорией, — Чарльз Дарвин, проводивший эксперименты на Галапагосском острове в 1830-х годах. до Дарвина было много других ученых, таких как Эмпедокл, Аристотель, Уильям Пейли, Ламарк и Бюффон, которые признавали тот факт, что виды меняются со временем, но не понимали основную причину этого изменения или что адаптация была непрерывным процессом, который не имеет окончательная форма.
Теория адаптации предполагает, что с изменением среды обитания происходят еще три изменения: отслеживание среды обитания, генетическое изменение и вымирание.
Отслеживание среды обитания: организмы, которые раньше жили в окружающей среде, ищут такую среду, в которой они жили раньше.
Вымирание: когда виды не могут адаптироваться к новой среде или покинуть ее и найти новую среду для жизни, в конечном итоге вымрут и вымрут.
Генетические изменения: Генетические изменения — естественный отбор, позволяющий организму с небольшими мутациями лучше выживать в окружающей среде по сравнению с другими организмами в данной области.
Пример. Дарвин наблюдал за черепахами, живущими на двух островах. Черепахи на одном острове ели с низа, из-за чего у них были более короткие ноги и прямой панцирь. Когда эти черепахи мигрировали на другой остров, где пищевые ресурсы были на высоте, черепахи с более длинными ногами смогли есть, выжить и размножаться. Со временем их шеи также удлинились, а панцири стали круглыми, и эти приспособления передавались из поколения в поколение черепахам, живущим на этом острове.
Что вызывает адаптацию?
Адаптация могла произойти в результате мутации или естественного отбора. Мутация — это внезапное генетическое изменение, которое может быть случайным или случайным. Мутация возникает из-за любого изменения нуклеотидной последовательности ДНК и замены одной пары нуклеотидов. Определенные мутации вредны для человека, в то время как определенные мутации могут быть полезны для выживания организма в борьбе за существование.
Пример. Птица рождается с длинным клювом, что помогает ей захватывать пищу и, следовательно, лучше выжить.Таким образом, эта птица может лучше выжить и больше размножаться. Этот ген продолжает передаваться по наследству из поколения в поколение.
Естественный отбор — это процесс, в котором люди с преимуществом в выживании будут выжить в окружающей среде, а люди с менее благоприятными чертами умрут из-за давления окружающей среды. В области ресурсы фиксированы или ограничены. Среди видов этой области существует конкуренция в борьбе за доступные ресурсы. В популяции некоторые особи обладают чертами, которые могут быть структурными или поведенческими, что позволяет им лучше выживать и воспроизводиться.В борьбе за существование некоторые особи демонстрируют различные вариации, которые имеют репродуктивное преимущество и дают больше потомства, чем другие виды. Любая неблагоприятная вариация отбирается от их присутствия, создавая селективный недостаток для этого организма.
Пример. На острове Галапагос существовали разные виды зябликов, различия между которыми заключались в характеристиках клюва. Природа выбрала наиболее подходящий клюв из менее полезных.
Какие бывают типы адаптации?
Адаптации бывают разных типов:
1.структурные адаптации
Структурные адаптации — это изменения в структуре живого организма с целью лучшей адаптации к окружающей среде.
Пример. Как пустынные растения адаптировались к условиям пустыни, где очень мало воды и высокая температура. Растения, называемые суккулентами, адаптировались к условиям пустыни, накапливая воду внутри себя, чтобы компенсировать более низкую доступность воды.
2.Поведенческая адаптация
Поведенческая адаптация — это изменение поведения организма с целью лучшего выживания в окружающей среде.
Пример — миграция — птица мигрирует на юг зимой, так как там больше пищи.
3. Физиологические адаптации
Физиологические адаптации — телесные процессы, которые помогают организму выжить / лучше размножаться в окружающей среде. Эти адаптации могут быть разными способами, которыми организм реагирует на стимулы из окружающей среды.
Пример. Животное, живущее в холодных регионах, будет иметь такие черты, как густой мех и короткие уши, чтобы уменьшить потерю тепла. Физиологическая адаптация здесь — это дрожь, чтобы генерировать больше тепла, когда становится холодно.
4. Коадаптация
Коадаптация, когда два или более вида симбиотически связаны друг с другом для их выживания и совместной адаптации, это называется коадаптацией.
Пример. У колибри длинные клювы, которые помогают птице улавливать нектар с определенных растений, при этом он опрыскивается пыльцевыми зернами.Таким образом, пыльцевые зерна распределяются, и колибри получают пищу.
Почему адаптация важна для видов?
Каждый организм имеет естественную среду обитания, которая является его домом. Здесь удовлетворяются основные потребности организма, такие как пища, кров, вода и другие потребности. Всем организмам необходимо адаптироваться к своей среде обитания, чтобы выжить. Адаптация — это изменение структуры или поведения организма, которое помогает ему лучше выжить в среде обитания.Это означает, что организму придется адаптироваться к климатическим условиям, хищникам и конкурировать за те же ресурсы с другими организмами, живущими в этой среде обитания.
Животные живут в среде, где существует несколько факторов, таких как климат, виды растений и других животных, которые могут быть хищниками или могут конкурировать с ними за ресурсы. Чтобы выжить, животное должно адаптироваться ко всем этим факторам.
Адаптация защищает животных от хищников.Хороший пример — возможность маскировки. Камуфляж — это приспособление, которое помогает организму сливаться с окружающей средой, чтобы он был незаметен. Адаптации также помогают организму защититься от суровой погоды. На Аляске, где температура очень низкая, животные хранят пищу в своем теле и защищаются от холодного климата густыми мехами.
Эволюционные адаптации к новой среде, как правило, обращают вспять пластические фенотипические изменения
Распространенность реверсии экспрессии в экспериментальной эволюции
Мы идентифицировали пять исследований, в которых было проведено шесть различных экспериментов по адаптации, и собраны данные транскриптома, подходящие для нашего анализа.Эти шесть экспериментов включали 10 повторов E. coli , адаптирующихся к высокотемпературной среде 17 , 6 повторов другого штамма E. coli , адаптирующегося к высокотемпературной среде 18 , 7 повторов E .coli , адаптирующийся к глицериновой среде 16 , 7 реплик E. coli , адаптирующийся к лактатной среде 16 , 1 реплика каждого из 12 различных штаммов дрожжей ( Saccharomyces cerevisiae ), адаптирующихся к ксилулозной среде 19 и 2 копии гуппи ( Poecilia reticulata ), адаптирующиеся к среде с низким уровнем хищничества 15 .Всего проанализировано 44 случая адаптации.
В каждом случае данные транскриптома были соответственно собраны для организмов в исходной среде (o для исходной стадии), вскоре после их воздействия на новую среду (p для пластической стадии) и по завершении экспериментальной эволюции. в новой среде (а для адаптированного этапа; рис. 1а). Обратите внимание, что время между o и p настолько короткое, что ожидается, что ни один вновь возникший аллель не достигнет заметной частоты на стадии p, чтобы повлиять на средний фенотип популяции.Уровень экспрессии каждого гена рассматривается как признак. Пусть уровни экспрессии гена на стадиях o, p и a равны L o , L p и L a соответственно. В каждом эксперименте мы сначала идентифицировали гены с заметным уровнем экспрессии PC, требуя PC = | L p — L o | быть больше предустановленного порога. Мы также идентифицировали гены с заметным уровнем экспрессии GC, потребовав GC = | L a — L p | быть больше, чем то же предустановленное значение отсечки.Для тех генов, которые демонстрируют как заметные ПК, так и заметные GC, мы спрашиваем, имеют ли эти два изменения одно и то же направление (то есть подкрепление) или противоположные направления (то есть реверсию; Рис. 1b, c). Мы использовали 20% от исходного уровня экспрессии гена (т.е. 0,2 L o ) в качестве порогового значения в приведенном выше анализе. Доля генов, демонстрирующих усиление уровня экспрессии ( C RI ), меньше, чем доля генов, демонстрирующих реверсию ( C RV ) в 42 из 44 адаптаций, и разница между C RI и C RV имеет значение в 40 из этих 42 случаев (номинальное значение P <0.05; двусторонний биномиальный тест; Рис. 1г). Среди двух приспособлений с C RI > C RV их разница значима только в одном случае (рис. 1d). Общее преобладание реверсии уровня экспрессии (т.е. 42 случая из 44) в адаптации является статистически значимым ( P = 1,1 × 10 −10 , двусторонний биномиальный тест). Та же тенденция очевидна, когда порог изменяется на 0,05 L o (дополнительный рис.1a) или 0,5 L o (дополнительный рис. 2a), что позволяет предположить, что вышеприведенные результаты устойчивы к выбору отсечки. Ясно, что транскриптомные данные не подтверждают гипотезу о том, что пластичность обычно способствует генетической адаптации.
Реверсия метаболического потока в адаптации к окружающей среде
Чтобы оценить общность вышеуказанного открытия и понять его основную причину, мы расширили сравнение между фенотипическим усилением и возвращением к метаболическим потокам (см. Введение).Поскольку наш метаболический анализ не предназначен для моделирования вышеуказанной экспериментальной эволюции или изменений экспрессии, используемые параметры не связаны с экспериментальной эволюцией. В частности, мы с помощью расчетов предсказали изменения пластического и генетического потока во время адаптации к окружающей среде с использованием i AF1260, реконструированной метаболической сети E. coli 23 . Мы использовали анализ баланса потоков (FBA) для прогнозирования оптимизированных потоков полностью адаптированных организмов в исходной (стадия o) и новой (стадия a) окружающей среде, соответственно, в предположении, что скорость производства биомассы, показатель приспособленности, максимальна. естественным отбором 20 .Прогнозы FBA достаточно хорошо согласуются с экспериментальными показателями для организмов, адаптированных к окружающей среде 24,25,26,27,28,29 и обычно используются при изучении взаимосвязей генотип-среда-фенотип 22,27,29,30,31 , 32,33,34,35,36,37 . При прогнозировании изменений пластического потока при изменении окружающей среды (стадия p) мы использовали минимизацию метаболической адаптации (MOMA) вместо FBA, потому что MOMA лучше воспроизводит немедленную реакцию потока на возмущения 21 (см. Методы).Мы рассматривали поток каждой реакции в метаболической сети как черту и моделировали изменения окружающей среды, изменяя источник углерода, доступный для сети. В i AF1260 существует 258 различных обменных реакций, каждая из которых переносит свой источник углерода. Поэтому мы исследовали 258 различных сред с одним источником углерода.
Мы начали анализ с использования глюкозы в качестве источника углерода в исходной среде, потому что эта среда была эталоном в i AF1260 Construction 23 .Затем мы рассмотрели адаптации E. coli к 257 новым средам, каждая из которых имеет свой источник одного углерода. Мы обнаружили, что эти новые среды естественным образом разделяются на две группы в прогнозируемой MOMA скорости производства биомассы, что является показателем пригодности на стадии p ( f p ) (дополнительный рис. 3). Одна группа показывает f p <10 −4 , что позволяет предположить, что E. coli вряд ли сохранится в этих новых условиях.Поэтому мы сосредоточились на оставшихся 50 новых средах с f p > 10 −4 , к которым E. coli предположительно может адаптироваться (дополнительная таблица 1).
Определяя усиление и реверсию потока и используя порог 0,2 L o , как в анализе транскриптома, мы обнаружили, что C RV значительно больше, чем C RI (номинал P < 10 −10 , двусторонний биномиальный тест) в каждой адаптации.Вероятность того, что все 50 адаптаций покажут C RV > C RI , составляет 1,8 × 10 −15 (двусторонний биномиальный тест; рис. 2a), что свидетельствует об общем преобладании реверсии потока. Среднее значение и медиана C RV составляют 30,2% и 30,5% соответственно, а для C RI — только 1,0% и 0,8% соответственно. Вышеупомянутая тенденция сохраняется, когда мы изменяем порог на 0,05 L o (дополнительный рис.1b) или 0,5 L o (дополнительный рис. 2b). Поскольку проблема FBA или MOMA может иметь несколько решений, порядок реакций в стехиометрической матрице может повлиять на конкретное решение, предоставляемое решателем. Тем не менее, когда мы случайным образом перетасовали порядок реакции в i AF1260, общая картина C RV > C RI не изменилась (дополнительный рисунок 4a). Поскольку квадратичное программирование, требуемое MOMA, труднее решить, чем линейное программирование, используемое в FBA, C RV могло быть завышено по сравнению с C RI .Чтобы устранить эту потенциальную проблему, мы разработали MOMA на основе квадратичного программирования под названием «MOMA-b» и использовали его вместо FBA для прогнозирования потоков на этапе a (см. Методы), но обнаружили, что C RV по-прежнему превышает C. RI (дополнительный рис. 4б). Таким образом, эта тенденция не является техническим артефактом различия решателей между MOMA и FBA.
Рис. 2Преобладание реверсии потока в экологических адаптациях E. coli . a Фракции реакций с усиливающим ( C RI ) и реверсивным ( C RV ) изменениями потока, соответственно, при адаптации от среды глюкозы к каждой из 50 новых сред. Каждая полоса представляет собой адаптацию к новой среде. Равенство долей усиливающих и реверсивных реакций проверяется двусторонним биномиальным тестом. Когда C RV > C RI , P -значения отображаются следующим образом: * P <0.05; ** P <10 −10 ; *** P <10 −100 ; когда C RV < C RI , P -значения отображаются следующим образом: o P <0,05; oo P <10 −10 ; ООО П <10 −100 . b Классификация реверсии по трем категориям на основе того, является ли фенотипическая ценность в исходной среде недостаточно восстановленной, восстановленной или чрезмерно восстановленной. c Доли трех категорий реверсии в каждой из 50 адаптаций. d Доля усиливающих реакций по сравнению с реверсивными реакциями ( C RI / C RV ) в адаптациях E. coli по крайней мере к 20 новым средам из каждой из 41 исследованной исходной среды. Коэффициенты C RI / C RV для всех адаптаций из каждой исходной среды представлены в виде прямоугольной диаграммы, где нижний и верхний края прямоугольника представляют собой первый (qu 1 ) и третий ( qu 3 ) квартилей, соответственно, горизонтальная линия внутри прямоугольника указывает медианное значение (md), усы простираются до самых крайних значений внутри внутренних ограждений, md ± 1.5 (qu 3 — qu 1 ), а кружки представляют значения за пределами внутренних ограждений (выбросы)
Реверсия потока в значительной степени восстанавливает исходные потоки
Чтобы проверить, восстанавливает ли реверсия потока во время генетической адаптации потоки на стадии o мы сравнили полное изменение TC = | L a — L o | с 0,2 л o для каждой реакции, показывающей обращение потока, в каждой адаптации. При TC <0,2 L o поток считается восстановленным (рис.2б). В противном случае мы дополнительно сравниваем ПК с GC. Если GC> PC, поток восстанавливается сверх нормы; в противном случае он недовосстановлен (рис. 2б). По 50 адаптациям средние доли реакций, показывающих «восстановленную», «чрезмерно восстановленную» и «недостаточно восстановленную» реверсию потока, составляют 26,4%, 3,1% и 0,7% соответственно, а медианы — 30,2%, 0,3 % и 0,1% соответственно (рис. 2в). Ясно, что реверсия потока в значительной степени восстанавливает потоки на стадии o.
Преобладание реверсии потока независимо от исходной среды
Чтобы исследовать общность нашего открытия преобладания реверсии потока, мы также исследовали адаптации с исходной средой, не содержащей глюкозы.Однако для многих исходных сред только несколько новых сред могут быть адаптированы метаболической сетью E. coli . Таким образом, мы сосредоточились на 41 исходной среде (включая ранее использованную среду глюкозы), каждая из которых имеет более 20 адаптируемых (т. Е. f p > 10 −4 ) новых сред (дополнительная таблица 2). Для каждой из этих исходных сред мы вычислили коэффициент C RI / C RV для каждой адаптируемой новой среды и обнаружили, что оно обычно ниже 0.1 (рис. 2г). Затем мы вычислили медианное значение C RI / C RV для всех адаптируемых новых сред из каждой исходной среды. В 41 исходной среде наибольшая медиана C RI / C RV составляет 0,11, а медиана медианы C RI / C RV составляет всего 0,02. Следовательно, независимо от исходной среды, реверсия потока гораздо более распространена, чем подкрепление во время генетической адаптации к новой среде.
Почему фенотипическая реверсия встречается чаще, чем подкрепление
Наше открытие, что фенотипическое подкрепление не только не больше, но и гораздо реже, чем реверсия, является неожиданным и, следовательно, требует объяснения. Наблюдение за этой тенденцией как в транскриптомном, так и в флуксомном анализах указывает на общий лежащий в основе механизм, который, как мы предполагаем, заключается в возникновении PC> TC. Геометрически очевидно, что когда PC> TC, GC должен перевернуть PC (левая и средняя диаграммы в верхнем ряду на рис.3а). Напротив, когда PC
Рис. 3Причина преобладания фенотипической реверсии в адаптации. a Схема, иллюстрирующая модель. Верхняя часть показывает, что если пластическое изменение (PC) больше, чем общее изменение (TC), генетическое изменение (GC) должно полностью изменить PC (левая и средняя диаграммы). Одна из причин PC> TC заключается в том, что разница в приспособленности между организмами на стадиях o и p больше, чем между стадиями o и a (правая диаграмма). В нижней части показано, что если PC
Для поиска эмпирических доказательств приведенного выше объяснения для каждого из 44 случаев Экспериментальной эволюции мы рассчитали долю генов, изменения экспрессии которых удовлетворяют PC> TC (рис.3б). Средняя и медианная доли составляют 0,51 и 0,48 соответственно. Более того, после того, как мы удалим все гены, для которых PC> TC, больше не будет избытка реверсии (дополнительный рис. 5a), что указывает на достаточность нашего объяснения. Точно так же мы вычислили долю метаболических реакций, показывающих PC> TC при адаптации метаболической сети E. coli от глюкозной среды к каждой из 50 новых сред (рис. 3c). Среднее и медианное значение составляют 0,85 и 0.93 соответственно. Точно так же после удаления реакций, показывающих PC> TC, нет общей тенденции к большему возврату, чем подкреплению в 50 адаптациях (дополнительный рис. 5b). Эти результаты транскриптома и флуксома подтверждают, что избыток реверсии по сравнению с подкреплением объясняется возникновением PC> TC для значимых фракций признаков.
Почему ПК превосходит TC по многим характеристикам? Вероятная причина в том, что ПК позволяют организмам выживать при внезапном изменении окружающей среды, но их приспособленность значительно снижается по сравнению с исходной средой, а также после адаптации к новой среде.Таким образом, общее физиологическое состояние организмов может быть весьма схожим между адаптированными стадиями в исходной и новой окружающей среде, но сильно отличаться на стадии пластической низкой приспособленности сразу после изменения окружающей среды. Это может объяснить, почему PC превышает TC по многим признакам, независимо от того, являются ли значения признаков причинами или следствиями приспособленности и физиологии организма.
Мы нашли убедительные доказательства вышеупомянутой модели с помощью анализа метаболической сети. Во-первых, используя прогнозируемую производительность биомассы как показатель пригодности, мы сравнили E.coli на пластической стадии ( f p ) и после адаптации к новой среде ( f a ) относительно исходной среды глюкозы для каждой из адаптаций к 50 новым среды. Во всех случаях f p <1 (рис. 3d), подтверждая, что сдвиги окружающей среды вызывают снижение приспособленности до генетической адаптации. Мы обнаружили, что f a обычно близко к 1, хотя в некоторых новых средах оно намного больше 1 (рис.3d). В логарифмической шкале 10 f p больше отличается от 1, чем f a в 43 из 50 адаптаций ( P = 1.0 × 10 −7 ; односторонний бином тестовое задание). Во-вторых, наша модель предполагает связь между изменениями потока и изменениями приспособленности 22 . Между 50 адаптациями глюкозной среды существует сильная отрицательная корреляция между f p и средним PC (Спирмен ρ = -0.98, P <10 −300 ; Рис. 3д). Противоположная корреляция существует между f a и средним TC ( ρ = 0,57, P = 1,1 × 10 −5 ; рис. 3f). Вместе наш анализ показывает, что основная причина более высокой частоты фенотипической реверсии, чем подкрепление во время адаптации, заключается в том, что с точки зрения приспособленности и связанных фенотипов организмы на стадии p более отличаются от организмов на стадии a, по сравнению с организмами на стадии o. .
Фенотипическая реверсия в случайных метаболических сетях
PC и GC в уровне экспрессии генов и метаболическом потоке во время адаптации зависят, соответственно, от регуляторной сети и метаболической сети соответствующих видов. Поскольку эти сети являются результатом миллиардов лет эволюции, возникает вопрос, связано ли преобладание фенотипической реверсии с эволюционной историей изучаемых видов, особенно с окружающей средой, в которой вид и его предки были выбраны в прошлом, или с присущей им природой. свойство любой функциональной системы.Чтобы ответить на этот вопрос, мы применили тот же анализ к 500 функциональным случайным метаболическим сетям, ранее созданным 22 . Эти сети были построены из i AF1260 путем замены его реакций произвольно выбранными реакциями из вселенной всех метаболических реакций в Киотской энциклопедии генов и геномов 38 до тех пор, пока сеть имеет ненулевую пригодность, предсказанную FBA в среда глюкозы при каждой реакции обмена 39 .
Только 20 новых сред, к которым i AF1260 может адаптироваться (из среды глюкозы), адаптируются как минимум в 20 из 500 случайных сетей.Таким образом, мы проанализировали адаптацию случайных сетей к каждой из этих 20 новых сред, при этом среда глюкозы была исходной средой. Для каждой новой среды медиана C RV всех случайных сетей, которые могут адаптироваться к этой среде, обычно составляет около 0,1 (прямоугольные диаграммы на рис. 4a), при этом медиана медианы C RV составляет 0,11. Напротив, медиана C RI в случайных сетях для новой среды обычно ниже 0.01 (прямоугольные диаграммы на рис. 4b), при этом медиана медианы C RI составляет 0,0033. Медиана C RI / C Отношение RV в случайных сетях для новой среды обычно ниже 0,05 (прямоугольная диаграмма на рис. 4c), при этом медиана медианы C RI / C RV , равное 0,0033. Ясно, что преобладание реверсии потока также очевидно в функциональных случайных сетях, предполагая, что это свойство присуще любой функциональной метаболической сети, а не продукту конкретных эволюционных историй.Действительно, механистическое объяснение этого свойства в реальных организмах (рис. 3) справедливо в случайных метаболических сетях, рассмотренных здесь. В частности, доля реакций, демонстрирующих PC> TC, значительна (рис. 4d), а f p в основном меньше 1 (рис. 4e). Кроме того, f p обычно больше отличается от 1, чем f a в масштабе журнала 10 , потому что | log 10 f p | — | log 10 f a | в основном положительный (рис.4е).
Рис. 4Преобладание реверсии потока в случайных метаболических сетях. Фракции реакций, показывающих реверсию потока ( C RV ) ( a ), доли реакций, показывающих усиление флюса ( C RI ) ( b ), C RI / C отношения RV ( c ), доля реакций, показывающих PC> TC ( d ), f p ( e ) и | log 10 f p | — | журнал 10 f a | ( f ) в адаптации случайных сетей из среды глюкозы к каждой из 20 исследованных новых сред.Для каждой новой среды значения, оцененные из различных случайных сетей, показаны в виде прямоугольной диаграммы с символами, поясненными в легенде на рис. 2d. Соответствующие значения для сети E. coli i AF1260 показаны красными ромбами.
Интересно, однако, что для 19 из 20 новых сред C RV в метаболической сети E. coli превышает медианное значение. C RV в случайных сетях (рис. 4а). Аналогичная, но менее очевидная тенденция сохраняется для C RI (рис.4б). Для 16 из 20 новых сред C RI / C RV из E. coli меньше медианы C RI / C RV случайных сетей ( P = 0,012, двусторонний биномиальный тест; рис. 4в). Следовательно, хотя метаболическая сеть E. coli и случайные сети демонстрируют преобладание реверсии потока, это явление более выражено у E. coli . Механически это несоответствие, по крайней мере качественно, объясняется нашей моделью в предыдущем разделе.В частности, для 15 из 20 новых сред доля реакций E. coli с PC> TC превышает соответствующую медианную долю в случайных сетях ( P = 0,021, односторонний биномиальный тест; рис. 4d). Для всех 20 новых сред f p из E. coli ниже медианы f p случайных сетей ( P = 9,5 × 10 −7 , односторонний биномиальный тест ; Рис. 4д). Для 19 из 20 новых сред | log 10 f p | — | журнал 10 f a | для E.coli больше, чем соответствующее медианное значение для случайных сетей ( P = 2,0 × 10 −5 , односторонний биномиальный тест; рис. 4f). Но почему f p из E. coli ниже, чем у случайных сетей? Одно из возможных объяснений состоит в том, что состав и структура метаболической сети E. coli были эволюционно оптимизированы для роста в среде глюкозы и / или связанных средах, в то время как то же самое не верно для случайных сетей, которые были необходимы только для быть жизнеспособным в среде глюкозы.В результате, когда глюкоза заменяется новым источником углерода в новой среде, приспособленность E. coli существенно падает, но приспособленность случайных сетей может снизиться лишь незначительно. Хотя абсолютная пригодность на пластической стадии может быть выше для E. coli , чем для случайных сетей, относительная пригодность, равная f p , ожидается, будет ниже для E. coli , чем случайная. сети. Таким образом, более высокая распространенность реверсии потока по сравнению с армированием в E.coli , чем случайные сети, вероятно, является побочным продуктом более сильного отбора E. coli по сравнению со случайными сетями в исходной среде, использованной в нашем анализе адаптации.
Реверсия встречается по крайней мере так же часто, как усиление, даже для признаков с заметным TC
В предшествующем анализе транскриптомов (рис. 1d) и флуксомов (рис. 2a) мы рассмотрели все признаки, демонстрирующие заметные PC и GC. Однако в сравнительных и эволюционных исследованиях фенотипы на стадии p обычно недоступны.В результате биологи-сравнители и эволюционисты обычно сосредотачиваются на чертах, фенотипические значения которых различаются между стадиями o и a, несмотря на то, что другие черты также могли претерпевать адаптивные изменения (от значений на стадии p до значений на стадии a). Чтобы выяснить, применимы ли наши предыдущие результаты к чертам, которые являются предметом изучения большинства сравнительных и эволюционных биологов, мы сосредоточимся на подмножестве проанализированных выше черт, которые удовлетворяют условию TC> 0,2 L o .Из 44 случаев экспериментальной эволюции 33 показали C RV > C RI ( P = 0,0013, двусторонний биномиальный тест), в 30 из которых C RV значительно превышает C RI (номинал P <0,05; двусторонний биномиальный тест; рис. 5а). Из 50 экологических адаптаций метаболической сети E. coli , происходящих из среды глюкозы, три случая имели равное количество реверсии потока и подкрепления.Из оставшихся 47 случаев в 22 было больше реверсии, чем подкрепления, а в 25 — обратное ( P = 0,77, двусторонний биномиальный тест; рис. 5b). Когда C RI значительно отличается от C RV , 15 случаев показали C RV < C RI , в то время как 11 показали противоположное ( P = 0,70, двусторонний биномиальный тест; рис. 5б). Следовательно, даже среди признаков с TC> 0,2 L o нет свидетельств значительно большего подкрепления, чем реверсия.Следует отметить, что в приведенном выше метаболическом анализе в среднем 139 реакций удовлетворяли TC> 0,2 L o на адаптацию. Поскольку все изменения потока, наблюдаемые при максимизации приспособленности, необходимы и, следовательно, по определению являются полезными, даже адаптация к простому изменению источника углерода, очевидно, включает в себя гораздо больше, чем несколько реакций.
Рис. 5Доля усиливающих признаков ( C RI ) не больше, чем доля реверсивных признаков ( C RV ) в адаптациях, даже если общее изменение превышает заданное пороговое значение.Черты удовлетворяющие | L a — L o | > 0,2 L o , | L p — L o | > 0,2 L o , и | L a — L p | > 0,2 L o классифицируются на усиливающие и меняющие признаки в зависимости от того, имеют ли генетические и пластические изменения одинаковое или противоположное направление. a Фракции генов с усилением и обращением изменений экспрессии, соответственно, в экспериментальной эволюции.Указаны организмы, а также новая среда, к которой организмы приспосабливались. Каждая полоса представляет собой адаптацию. b Фракции реакций с прогнозируемыми усиливающими и обратными изменениями потока, соответственно, в адаптации E. coli к 50 новым средам из среды глюкозы. На обеих панелях равенство долей усиливающих и реверсивных реакций проверяется двусторонним биномиальным тестом. Когда C RV > C RI , P -значения отображаются следующим образом: * P <0.05; ** P <10 −10 ; *** P <10 −100 ; когда C RV < C RI , P -значения отображаются следующим образом: o P <0,05; oo P <10 −10 ; ooo P <10 −100
Адаптация и эволюция биологических материалов | Интегративная и сравнительная биология
Аннотация
Исследования биологических материалов часто сосредотачиваются на впечатляющих свойствах материалов, созданных в природе.В процессе, однако, это исследование часто игнорирует экологию материалов, контексты организма, связанные с тем, как на самом деле используется биологический материал. В биологии материалы жизненно важны для взаимодействия организмов с окружающей средой и их физиологией, а также обеспечивают записи об их филогенетических отношениях и давлениях отбора, которые движут биологическими новинками. В статьях этого симпозиума мы даем обзор передовых разработок в области биологического материаловедения. Собранные исследования открывают новые взгляды на фундаментальные концепции материалов, предлагают удивительное понимание биологических инноваций и оспаривают границы методов определения характеристик материалов.Охватываемые темы, системы и дисциплины дают представление о широком спектре современных биологических материалов. Они также демонстрируют необходимость прогрессивного «мышления всего организма» при характеристике биологических материалов и важность рассмотрения биологических материалов в соответствующих биологических контекстах.
Введение
Как биологи, мы заинтересованы в материалах в контексте. Материалы, которые строят и используют организмы, не являются результатом внезапного вдохновения de novo , они являются продуктом накопленной истории взаимодействий между организмом, окружающей средой и филогенезом (Knoll 2003; рис.1). В отличие от искусственно созданных материалов, биологические материалы ограничены гораздо более ограниченной палитрой уже существующих ингредиентов, и все же они достигают невероятных диапазонов функциональности, опосредуя отношения между организмом и окружающей средой, приспособляемостью и выживанием (Wegst and Ashby 2004; Fratzl and Barth 2009). . Существует значительный интерес к разработке биологических материалов, но попытки в этом направлении все еще не позволяют воспроизвести биологические ткани и процессы (Holland et al.2012; Eder et al.2018). В основе этой проблемы лежит проблема масштаба: характеристика материалов часто требует подходов, которые ограничивают внимание очень точным разрешением, что позволяет легко потерять из виду организм, производящий материалы. Хотя десятилетия исследований в области биофизики, биомеханики и биохимии доказали полезность использования различных дисциплин в биологических темах, иногда эти области забывают, что биология может предложить взамен.
Рис. 1
Схема факторов, влияющих на эволюцию и адаптацию биологических материалов, подчеркивая необходимость «мышления всего организма» в исследованиях биологических материалов.(Объяснение движется по часовой стрелке из верхнего левого угла, термины, используемые на рисунке, выделены жирным шрифтом. Цветную версию рисунка см. В онлайн-формате PDF.)
Биологические материалы сталкиваются с множеством ограничений , которые определяют их эволюцию и приспособление. В общем смысле это могут быть:
Филогенетический : аспекты истории эволюции, которые ограничивают пути будущего развития.
Окружающая среда : например, абиотические факторы, такие как доступность ресурсов, влажность или температура, связанные с окружающей средой организма (биотические факторы, такие как давление на уровне популяции, также попадают в эту категорию).
Physical : например, конструктивные ограничения, связанные с законами масштабирования материала или характеристиками.
Эти факторы взаимодействуют с генотипом организма, тем самым формируя и оказывая отбор давления на фенотип организма.
Понимание действия различных ограничений на конкретный биологический материал сложно, потому что фенотипы организма состоят из огромного количества взаимодействующих компонентов (например, материалов ).На организацию этих материалов — их состав , ультраструктуру , структурную иерархию — можно повлиять посредством упомянутых ограничений. Организационные аспекты биологического материала также могут определяться взаимодействиями с другими материалами в организме и взаимодействиями между уровнями организации внутри материала. Например, доступность химического вещества в окружающей среде может повлиять на химический состав биологического материала, который, в свою очередь, влияет на структуру ткани, частью которой он является.
Все эти взаимодействия влияют на функцию или характеристики биологического материала (например, жесткость панциря жука или его способность воспроизводить структурный цвет), что затем определяет, насколько хорошо этот материал может выполнять определенную роль в организме ( например, защита мягких внутренних компонентов, возможность маскировки). Взаимодействие многих функциональных материалов организма может наложить дополнительные ограничения на характеристики материала, так что оптимальные характеристики для организма могут отличаться от максимальных характеристик материала в массе.В качестве гипотетического примера, если структурные аспекты, определяющие жесткость и цвет, в некоторой степени противоречат друг другу и давление отбора для хорошей маскировки велико, панцирь жука может быть менее жестким, чем это возможно, с учетом составляющих его материалов. Фактически, мультиролевой оптимум может быть более уместным с экологической точки зрения, когда жесткость материала и цветовые характеристики могут быть «неоптимальными», но вместе они позволяют панцирю одновременно защищать и маскировать.
То, как организм и его материалы функционируют по отношению к остальной части населения и в контексте данных ограничений, будет определять его репродуктивный успех ( приспособленность ).Таким образом, организм, его материалы и его ограничения являются фильтрами для вариаций на уровне популяции, формируя, например, какие наследственные материальные особенности устанавливаются, изменяются и отбираются в процессе эволюции (даже если не все эти особенности обязательно выгодны). ).
Рис. 1
Схема факторов, влияющих на эволюцию и адаптацию биологических материалов, подчеркивая необходимость «мышления всего организма» в исследованиях биологических материалов. (Объяснение движется по часовой стрелке из верхнего левого угла, термины, используемые на рисунке, выделены жирным шрифтом.Цветную версию рисунка см. В онлайн-формате PDF.)
Биологические материалы сталкиваются с рядом ограничений , которые определяют их эволюцию и адаптацию. В общем смысле это могут быть:
Филогенетический : аспекты истории эволюции, которые ограничивают пути будущего развития.
Окружающая среда : например, абиотические факторы, такие как доступность ресурсов, влажность или температура, связанные с окружающей средой организма (биотические факторы, такие как давление на уровне популяции, также попадают в эту категорию).
Physical : например, конструктивные ограничения, связанные с законами масштабирования материала или характеристиками.
Эти факторы взаимодействуют с генотипом организма, тем самым формируя и оказывая отбор давления на фенотип организма.
Понимание действия различных ограничений на конкретный биологический материал сложно, потому что фенотипы организма состоят из огромного количества взаимодействующих компонентов (например, материалов ).На организацию этих материалов — их состав , ультраструктуру , структурную иерархию — можно повлиять посредством упомянутых ограничений. Организационные аспекты биологического материала также могут определяться взаимодействиями с другими материалами в организме и взаимодействиями между уровнями организации внутри материала. Например, доступность химического вещества в окружающей среде может повлиять на химический состав биологического материала, который, в свою очередь, влияет на структуру ткани, частью которой он является.
Все эти взаимодействия влияют на функцию или характеристики биологического материала (например, жесткость панциря жука или его способность воспроизводить структурный цвет), что затем определяет, насколько хорошо этот материал может выполнять определенную роль в организме ( например, защита мягких внутренних компонентов, возможность маскировки). Взаимодействие многих функциональных материалов организма может наложить дополнительные ограничения на характеристики материала, так что оптимальные характеристики для организма могут отличаться от максимальных характеристик материала в массе.В качестве гипотетического примера, если структурные аспекты, определяющие жесткость и цвет, в некоторой степени противоречат друг другу и давление отбора для хорошей маскировки велико, панцирь жука может быть менее жестким, чем это возможно, с учетом составляющих его материалов. Фактически, мультиролевой оптимум может быть более уместным с экологической точки зрения, когда жесткость материала и цветовые характеристики могут быть «неоптимальными», но вместе они позволяют панцирю одновременно защищать и маскировать.
То, как организм и его материалы функционируют по отношению к остальной части населения и в контексте данных ограничений, будет определять его репродуктивный успех ( приспособленность ).Таким образом, организм, его материалы и его ограничения являются фильтрами для вариаций на уровне популяции, формируя, например, какие наследственные материальные особенности устанавливаются, изменяются и отбираются в процессе эволюции (даже если не все эти особенности обязательно выгодны). ).
Статьи в этом сборнике симпозиума являются призывом к более организационному взгляду на исследования биологических материалов, который учитывает не только биологию, но также экологию и филогению представляющих интерес материалов.По словам Фратцля и Барта (2009, 422), «исследование биомиметических материалов невозможно без надлежащих биологических исследований, включая тщательный анализ того, для чего этот материал сделан в условиях видоспецифичного поведения организма и экологической ситуации». Хотя перспективы материалов в биологии организма все еще сравнительно редки, методы характеризации, которые десять лет назад были в основном известны только материаловедам (например, синхротронная микроКТ, рамановская спектроскопия, наноиндентирование, FIB-SEM и крио-ТЕМ), становятся все более доступными и востребованными. биологами.Перекрывающийся интерес к методам характеризации и системам биологических исследований создает готовые платформы для междисциплинарного взаимодействия между дисциплинами естествознания и физики.
Исследования, представленные в настоящем документе, сосредоточены на разнообразии систем и тем в различных организационных масштабах: от эволюции формы и функции скелета и раковины до опосредованного материалом восприятия вибрации, светопропускания и демпфирования нагрузки, цвета конструкции до закрепления растений и животных инструменты. Авторы происходят из самых разных дисциплин, включая эволюционную биологию, морфологию, биохимию, материалы, компьютерные и инженерные науки, демонстрируя широту современных исследований материалов.Во введении мы представляем три тематические области, которые предлагают возможности для более глубокой интеграции между биологией организма и материаловедением: структурные иерархии, мышление в целом организма, а также адаптация и эволюция. Мы надеемся, что работа, собранная в этом выпуске, поможет биологам познакомиться с новыми линзами для просмотра их работы, в то время как исследователи, ориентированные на материалы, рассмотрят, как материалы адаптируются и развиваются вместе с организмами (рис. 1). Эти взаимные подходы могут способствовать пониманию инноваций в биологических материалах в биологическом контексте, а также способствовать созданию эффективных методов биомиметической инженерии.
Структурные иерархии
Очень ограниченный диапазон веществ, доступных для создания биологических тканей, означает, что Природа, в отличие от инженерии, обычно больше играет со структурой, чем с составом для достижения различных функций (Wegst and Ashby 2004; Dunlop et al. 2011; Eder et al. 2018). Общая стратегия естественного дизайна включает использование структурной иерархии, где различные структурные и морфологические особенности существуют в различных масштабах (Lakes 1993; Wegst and Ashby 2004).Таким образом, структурные устройства в очень малых масштабах (например, расположение кристаллов и / или волокон) могут вносить вклад в объемные свойства в гораздо более крупных масштабах (например, на уровне ткани или органа). Кость (Резников и др., 2018), эмаль (Ван Кастерен и Крофтс, 2019), моллюсковый перламутр (перламутр) (Баум и др., 2019) и шелк (Споннер и др., 2007) — все это хорошо известные биологические примеры. из иерархических материалов, структурированных таким образом, чтобы добиться чрезвычайно высокой прочности. Иерархические структуры также могут выполнять другие функции по отдельности или в сочетании, например, цвет (Saranathan et al.2015), прозрачность (Багге и др., 2019), адгезия (Бродочану и др., 2016) и / или самоочищение (Бартлотт и др., 2016).
В основе понимания функции (функций) данной структурной иерархии лежит необходимость преодоления огромного диапазона масштабных масштабов, от молекулы до метра (Cranford and Buehler 2010). Это делает проблемы биологических материалов одновременно сложными и идеальными для междисциплинарного сотрудничества внутри и за пределами сообщества сравнительной биологии. Биологи приучены думать об иерархических проблемах, связывая различные уровни эволюционного давления, анатомию организма и экологию сообщества в единую биологическую картину.Доступность методов магнитно-резонансной (Hesse et al., 2019), рентгеновской (Knötel et al., 2017) и электронной (Weber et al., 2014) методов визуализации в современных морфологических исследованиях подтолкнула инструментарий морфолога к более тонким и тонким масштабам. пока в процессе увеличивается количество деталей и информации, доступной для описания. Для управления потоком данных инструменты анализа и моделирования все больше автоматизируются, что делает невозможным многомасштабный высокопроизводительный анализ в масштабах, которые ранее были невозможны.Баум и др. (2019), например, показывают, что применение единого метода преобразования данных к наборам данных микроКТ может обеспечить быстрый и полуавтоматический анализ огромного количества морфологических признаков даже для очень разных типов анатомических данных. Хотя такие современные методы визуального анализа данных в значительной степени опираются на информатику (например, подходы машинного обучения), они всегда будут требовать биологических знаний (например, чтобы различать релевантный и нерелевантный материал при сегментации; Baum et al.2019), что делает знания об организме жизненно важными для руководства и ограничения оценки данных.
Возможно, в дисциплинах органической биологии менее ценится то, что концепции структурного дизайна также играют роль в управлении, казалось бы, сложной сборкой тканей на химических уровнях. Например, некоторые животные, производящие слизь или нити (пауки, мидии, миксины, бархатные черви), хранят необходимые биомолекулы-предшественники в высококонцентрированном жидком состоянии, которое может удивительно быстро превращаться в твердое вещество (например,ж., клетчатка) через определенные триггеры (например, механический сдвиг, изменение pH, концентрация соли и потеря воды; Holland et al.2012; Kurut et al.2015; Baer et al. 2019a). Эта инициированная самосборка запрограммирована в биохимическую структуру строительных блоков белка и их организацию более высокого порядка. Специфика процессов самосборки различается для разных таксонов, но их общие черты позволяют определить универсальные принципы проектирования материалов для устойчивого производства (например,г., полимеров, биомедицинских каркасов). Многие из физических и химических особенностей сборки также находят аналогии в гораздо более крупных масштабах, где группы организмов образуют запутанные системы, как в каплях червя (Aydin et al., 2019), плотах или башнях огненных муравьев (Phonekeo et al., 2017). , и фонтаны личинок мух (Шишков и др., 2019б). Эти системы имитируют сборку молекул и физику мягкой материи для решения проблем популяционного уровня, таких как преодоление физических препятствий (Phonekeo et al., 2017) или расширение доступа людей к ресурсам во время кормления (Shishkov et al.2019а).
Мышление всего организма
Как и в физиологических системах и системах на уровне организма, биологические материалы подвергаются функциональному уравновешиванию. Кости — это механические устройства и хранилища кальция, а чешуя рыбы обеспечивает защиту и влияет на гидродинамику. Поэтому особенности биологического материала часто нельзя понять вне экологии материала, его особой роли во взаимодействии организма с окружающей средой (рис. 1). Например, шелк исторически изучался в первую очередь как структурный материал, но он также выполняет важные функции восприятия вибраций.Эта многофункциональность помогает объяснить, с одной стороны, разнообразие типов шелка в полотнах и форме полотна (Mortimer et al., 2016), а с другой стороны, диапазон чувствительности встроенных датчиков вибрации в пауках, прядущих паутину. ноги (Мортимер и др., 2019). Органический контекст также важен для понимания функции нановолоконного паучьего шелка, который взаимодействует с воском кутикулы насекомых для достижения более высокой адгезии к добыче (Bott et al., 2017). Точно так же механическое вытягивание, необходимое для быстрого превращения улавливающей жидкость слизи бархатных червей в жесткие липкие нити, рассматривается в контексте, когда учитывается распыление слизи червя и борьба с добычей (Baer et al.2019b). Такая функциональная зависимость от контекста может затруднить прогнозирование конкретных функций только на основе структуры материала. Джоэл и Вайсбах (2019) описывают, как различные типы поверхностных структур — микроканалы, повторяющиеся столбы или капиллярные сети — используются в самых разных экологических задачах у животных, включая сбор воды в пустынях, транспортировку нефти для защитных выделений, самоочищающиеся поверхности. которые уменьшают количество патогенов и задержку воздуха при нырянии. Эти структурные особенности предлагают неоднозначные ключи к разгадке их функции в организме, делая их биологические роли легко ложно интерпретируемыми и подчеркивая важность интеграции экологических и поведенческих исследований в изучение биологических материалов.
Так же, как история загрузки важна для понимания потенциала искусственного материала к механическому отказу (например, из-за многоцикловой усталости), история жизни организма обеспечивает важный контекст для характеристик биологического материала с течением времени и использования. Например, хотя эмаль млекопитающих и эмелоид акулы схожи по составу, первая покрывает зубы, которые обычно служат всю жизнь (van Casteren and Crofts, 2019), тогда как последняя покрывает зубы, которые постоянно заменяются (Corn et al.2016). Нити прикрепления мидий (биссальные нити) долговечны, тогда как нити улавливающей слизи бархатных червей постоянно перерабатываются и регенерируются, хотя оба являются материалами на основе белков со структурными мотивами бета-листов (разница в перекрестном связывании: ковалентные в первом случае нековалентный во втором) (Baer et al. 2019b). Кроме того, функциональные свойства данного материала могут изменяться с помощью определенных триггеров, таких как зависящее от гидратации обратимое разворачивание капсул семян ледяных растений (активируемое набухающим слоем целлюлозы) (Harrington et al.2011) и открытие семенных коробочек Banksia после многократного воздействия огня (Huss et al. 2018). Методы тестирования биологических материалов в таких активных условиях становятся все более доступными. Например, более быстрая визуализация и более настраиваемые лабораторные устройства теперь позволяют выполнять «4D» (с временным разрешением) МРТ и микроКТ, включая этапов механических испытаний in situ , что позволяет охарактеризовать реакцию морфологии на нагрузку (например, Gustafsson et al. 2018; Hesse et al.2019). Такие технологические достижения расширяют наши возможности задавать поддающиеся количественной оценке вопросы в состояниях, более близких к физиологическим.
Контекстуализация адаптации и эволюции
Биология не статична, поэтому биологические материалы также должны приспосабливаться к различным ситуациям и средам и могут со временем развиваться. Новый рубеж в материаловедении — выйти за рамки статических состояний материала и охарактеризовать адаптируемые активные состояния материалов в различных временных масштабах (Burla et al.2019; де Пабло и др. 2019). Краткосрочная адаптация свойств активного материала может быть динамической, например, в способности стаи огненных муравьев модулировать широкий диапазон текучести и вязкоупругих свойств (Vernerey et al.2018), или более пассивной, как при транспортировке жидкости, которая увеличивает адгезию шелка. во время поимки добычи (Ботт и др., 2017). Материальная восприимчивость также может быть ограничивающей с биологической точки зрения, например, внезапное помутнение креветок с прозрачным телом при физиологическом стрессе, что делает их видимыми для хищников (Bagge et al.2017). В более длительных временных масштабах (например, связанных с ростом организма) изменения в структуре могут диктовать более эффективное использование пространства в скелетных тканях (Baum et al., 2019) и лучшую несущую способность, как в случае адаптивного изменения сосудистой архитектуры у драконова дерева. ветвей (Hesse et al., 2016).
В эволюционной шкале времени знание филогенетического распределения материала жизненно важно для понимания давления, которое сформировало его структуру и функцию. Выявление схожих материалов в биологии — это начало, и контекстуализация разнообразия материалов в контексте эволюционных отношений может предоставить больше биологической информации о конвергенции, а не о конвергенции.гомология и кооптация существующих инструментов для новых функций. Биология предлагает множество возможностей для метаанализов таких материалов из-за чрезвычайно широко распространенной природы важных компонентов основы ткани (например, карбонат кальция, кератин) и структурных мотивов (например, скрученная фанерная архитектура, трабекуляция) (Knoll 2003; Eder et al. 2018). Обширные исследования материалов уже дают представление о биологических особенностях, которые определяют форму птичьего яйца (Stoddard et al., 2017), эволюционное происхождение трехмерных фотонных кристаллов (Seago et al.2019) и взаимосвязь между изменением эмали и диетической экологией (Lucas et al., 2016). Точно так же, как высокопроизводительное секвенирование произвело революцию в обширных филогенетических исследованиях, теперь высокопроизводительные инструменты визуализации и анализа вместе с общими онлайн-хранилищами данных (например, oVert, MorphoMuseum, iDigBio, MorphoSource) облегчают исследования функциональной морфологии в более широком масштабе, более филогенетически значимые шкалы (Wipfler et al., 2016).
На пути к пониманию инноваций
Работы, представленные в этом выпуске, представляют собой небольшую выборку передовых подходов и междисциплинарных тем, которые характеризуют растущую область интегративных исследований биологических материалов.Симпозиум и этот сборник объединяют множество различных систем, дисциплин и перспектив в общем стремлении понять биологические материалы в их организменных системах отсчета. Биологическое материаловедение вступает в захватывающий этап, выходя за рамки визуализации и описания статических материалов и переходя к изучению более активных, адаптируемых систем. На этом пороге организменные биологи могут предложить глубокие взгляды на биологический контекст материальной структуры и функции, в то же время имея беспрецедентный доступ к инструментам и сотрудничеству, относящимся к обширным вопросам.Выбираются ли характеристики материала эволюционно? Как абиотические факторы ограничивают или усиливают функцию биоматериала? Как эти материалы адаптируются и развиваются вместе с организмами в разных средах? От морфологических структурных иерархий до более широких организменных и поведенческих контекстов, до адаптации и эволюции функциональных свойств в различных масштабах и временных масштабах — существует множество интересных областей, в которых взгляд на биологические материалы в целом может взаимно информировать наше понимание как биологических, так и материальных аспектов. системы.
Из симпозиума «Адаптация и эволюция биологических материалов», представленного на ежегодном собрании Общества интегративной и сравнительной биологии 3–7 января 2019 г. в Тампе, Флорида.
Благодарности
Организаторы хотели бы поблагодарить спикеров симпозиума и руководителей семинаров, чьи презентации заинтересовали и вдохновили аудиторию SICB. Было приятно работать со всеми вами. Мы также хотели бы поблагодарить Шахруз Амини, Джулию Чаумель, Джона Данлопа, Мэтью Харрингтона и Рональда Зайделя за полезные комментарии к рукописи.
Финансирование
Эта работа была предоставлена в связи с симпозиумом «Адаптация и эволюция биологических материалов» в Обществе интегративной и сравнительной биологии 2019 г. и была щедро поддержана многочисленными подразделениями Общества интегративной и сравнительной биологии (DCB, DCPB, DEE, DIZ, и DVM), Американское общество микроскопии, Компания биологов, журнал Bioinspiration and Biomimetics, Micro Photonics Inc., Overleaf и Thermo Fisher Scientific.
Список литературы
Aydin
YO
,Culver
J
,Tennenbaum
M
,Goldman
D
,Bhamla
MS.
2019
. Динамика червячной капли [аннотация]. В: Ежегодное собрание Общества интегративной и сравнительной биологии, 2019 г. Тампа, Маклин (Вирджиния): Общество интегративной и сравнительной биологии. п. S4–5.Baer
A
,Horbelt
N
,Nijemeisland
M
,Garcia
SJ
,Fratzl
P
,000000Schmidt
Харрингтон
МДж.
2019a
.Вызванный сдвигом β-кристаллит, разворачивающийся в нанокаплях конденсированной фазы, способствует образованию волокон в биологическом адгезиве
.ACS Nano
13
:4992.
Baer
A
,Schmidt
S
,Mayer
G
,Harrington
MJ.
2019b
.Волокна на лету: многомасштабные механизмы образования волокон при улавливании слизи бархатных червей
.Integr Comp Biol опубликовано в Интернете
(https://doi.org/10.1093/icb/icz048).Bagge
LE
,Kinsey
ST
,Gladman
J
,Johnsen
S.
2017
.Прозрачные креветки-анемоны ( Ancylomenes pedersoni ) становятся непрозрачными после физических нагрузок и физиологического стресса, что коррелирует с увеличением перфузии гемолимфы
.J Exp Biol
220
:4225
—33
.Багге
LE.
2019
.Не так однозначно, как может показаться: проблемы, связанные с прозрачным камуфляжем в океане
.Integr Comp Biol
опубликовано в Интернете (doi: 10.1093 / icb / icz066).Barthlott
W
,Mail
M
,Neinhuis
C.
2016
.Супергидрофобные иерархически структурированные поверхности в биологии: эволюция, структурные принципы и биомиметические приложения
.Philos Trans A Math Phys Eng Sci
374
:20160191.
Baum
D
,Weaver
JC
,Zlotnikov
I
,000
000
000
0009 Л,Дин
МН.
2019
.Высокопроизводительная сегментация мозаичных биологических структур с использованием преобразований случайных расстояний
.Integr Comp Biol
опубликовано в Интернете (doi: 10.1093 / icb / icz117).Bott
RA
,Baumgartner
W
,Bräunig
P
,Menzel
F
,Joel
A-C.
2017
.Повышение адгезии нитей, улавливающих крибелляты, эпикутикулярными восками насекомых-жертв проливает новый свет на эволюцию паутины
.Proc R Soc B
284
:20170363
.Brodoceanu
D
,Bauer
CT
,Kroner
E
,Arzt
E
,Kraus
T.
2016
.Иерархические биоинспирированные адгезивные поверхности — обзор
.Биоинспир Биомим
11
:051001
.Burla
F
,Mulla
Y
,Vos
BE
,Aufderhorst-Roberts
A
,Koenderink
GH.
2019
.От механической устойчивости к свойствам активного материала в биополимерных сетях
.Nat Rev Phys
1
:249
—63
.Кукуруза
KA
,Farina
SC
,Brash
J
,Summers
AP.
2016
.Моделирование взаимодействия зубов и жертв у акул: важность динамического тестирования
.R Soc Open Sci
3
:160141
—6
.Cranford
S
,Buehler
MJ.
2010
.Материомика: биологические белковые материалы, от нано до макро
.Nanotechnol Sci Appl
3
:127
—48
.де Пабло
JJ
,Джексон
NE
,Webb
MA
,Chen
LQ
,Мур
JE
,9000 R 9000 9000
JE
Morgan
000 ,Минтай
T
,Schlom
DG
,Toberer
ES
и др..2019
.Новые рубежи инициативы «Геном материалов»
.npj Comput Mater
5
:41
.Dunlop
JWC
,Weinkamer
R
,Fratzl
P.
2011
.Искусные интерфейсы в биологических материалах
.Матери сегодня
14
:70
—8
.Eder
M
,Amini
S
,Fratzl
P.
2018
.Биологические композиты — сложные структуры для функционального разнообразия
.Наука
362
:543
—7
.Fratzl
P
,Barth
FG.
2009
.Системы биоматериалов для механочувствительности и приведения в действие
.Природа
462
:442
—8
.Gustafsson
A
,Mathavan
N
,Turunen
MJ
,Engqvist
J
,Khayyeri
9000sson4 Hak,Hall
2018
.Связь многомасштабной деформации с микроструктурой кортикальной кости с использованием загрузки на месте, корреляции цифровых изображений и синхротронного рассеяния рентгеновских лучей
.Acta Biomater
69
:323
—31
.Харрингтон
MJ
,Razghandi
K
,Ditsch
F
,Guiducci
L
,Rueggeberg
M
ratzNeinhuis
C
,Burgert
I.
2011
.Как оригами раскладывание капсул с семенами ледяных растений, приводимых в действие гидроциклом
.Nat Commun
2
:337.
Hesse
L
,Masselter
T
,Leupold
J
,Spengler
000000 Speck4,
JG.
2016
.Магнитно-резонансная томография раскрывает функциональную анатомию и биомеханику живого драконова дерева
.Sci Rep
6
:32685.
Hesse
L
,Leupold
J
,Poppinga
S
,Wick
M
M
T
,Speck
T.
2019
.Устранение взаимосвязей между формой и структурой в растениях с помощью МРТ для переноса биомиметиков
.Integr Comp Biol
опубликовано в Интернете (doi: https: // doi.org / 10.1093 / icb / icz051).Голландия
C
,Vollrath
F
,Ryan
AJ
,Mykhaylyk
OO.
2012
.Шелк и синтетические полимеры: согласование 100 градусов разделения
.Adv Mater
24
:105
—9
.Huss
JC
,Schoeppler
V
,Merritt
DJ
,Best
C
,Maire
E
,O0004
0009000Adrien
Janssen
N
,Gladisch
J
,Gierlinger
N
и др..2018
.Климатически-зависимый механизм открывания семенных коробок Banksia , запускаемый при нагревании,
.Adv Sci
5
:1700572.
Joel
AC
,Weissbach
M.
2019
.Те же принципы, но разные цели: пассивное обращение с жидкостями в животном мире
.Integr Comp Biol
опубликовано в Интернете (https://doi.org/10.1093/icb/icz018).Knoll
AH.
2003
.Биоминерализация и эволюционная история
.Рев Mineral Geochem
54
:329
—56
.Knötel
D
,Seidel
R
,Prohaska
S
,Dean
MN
,Baum
D.
2017
.Автоматизированная сегментация сложных паттернов в биологических тканях: уроки мозаичного хряща ската
.PLoS One
12
:e0188018
.Курут
А
,Дико
С
,Лунд
М.
2015
.Димеризация терминальных доменов в белках шелка пауков контролируется электростатической анизотропией и модулируется гидрофобными пятнами
.ACS Biomater Sci Eng
1
:363
—71
.Озера
р.
1993
.Материалы со структурной иерархией
.Природа
361
:511
—5
.Lucas
PW
,Philip
SM
,Al-Qeoud
D
,Al-Draihim
N
,Saji
S
000, Casterenфургон 2016
.Структура и масштаб механики зубной эмали млекопитающих с точки зрения эволюции
.Evol Dev
18
:54
—61
.Mortimer
B
,Soler
A
,Siviour
CR
,Zaera
R
,Vollrath
F.
2016
2016
Настройка инструмента: звуковые свойства паутины
.J R Интерфейс Soc
13
.Мортимер
B
и др. .2019
.Вибрационный ландшафт паука: приспособления для передачи вибрационной информации в паутине сфер
.Integr Comp Biol
опубликовано в Интернете (doi: 10.1093 / icb / icz043).Phonekeo
S
,Mlot
N
,Монаенкова
D
,Hu
DL
,Тови
C.
2017
.Огненные муравьи постоянно восстанавливают тонущие башни
.R Soc Open Sci
4
:170475
.Резников
N
,Bilton
M
,Lari
L
,Stevens
MM
,Kröger
R.
2018
.Фрактальная иерархическая организация кости начинается на наномасштабе
.Наука
360
:eaao2189
.Саранатан
V
,Seago
AE
,Сэнди
A
,Нараянан
S
,Mochrie
SGJ
,0009000Ca Dufresne Ca Dufresne
Osuji
CO
,Prum
RO.
2015
.Структурное разнообразие биофотонных наноструктур членистоногих охватывает амфифильное фазовое пространство
.Nano Lett
15
:3735
—42
.Seago
AE
,Oberprieler
R
,Saranathan
V.
2019
.Эволюция радужной оболочки насекомых: происхождение трехмерных фотонных кристаллов у долгоносиков (Coleoptera: Curculionoidea)
.Integr Comp Biol
опубликовано в Интернете (https://doi.org/10.1093/icb/icz040).Шишков
O
,Hu
M
,Johnson
C
,Hu
DL.
2019a
.Личинки черной солдатской мухи питаются, образуя фонтан вокруг еды
.J R Интерфейс Soc
16
:20180735
.Шишков О., Требюшон Дж., Юнкер П.Дж., Франклин С., Ху DL. 2019b. Личинки черной солдатской мухи переставляются при сжатии.Интегр Комп Биол. опубликовано в Интернете (doi: 10.1093 / icb / icz133).
Sponner
A
,Vater
W
,Monajembashi
S
,Unger
E
,Grosse
F
0000009000000000Weiss9000
Weiss
Состав и иерархическая организация паучьего шелка
.PLoS One
2
:e998
.Стоддард
MC
,Йонг
EH
,Аккайнак
D
,Sheard
C
,Tobias
JA
de4 L. Maha2017
.Форма птичьего яйца: форма, функции и эволюция
.Наука
356
:1249
—54
.van Casteren
A
,Crofts
S.
2019
.Материалы жевания: материаловедение скромного зуба
.Integr Comp Biol
опубликовано в Интернете (https://doi.org/10.1093/icb/icz129).Vernerey
FJ
,Shen
T
,Sridhar
SL
,Wagner
RJ.
2018
.Как огненные муравьи контролируют реологию своих скоплений? Подход статистической механики
.Интерфейс JR Soc
15
:20180642.
Weber
B
,Tranfield
EM
,Höög
JL
,Baum
D4000 HymanT
,Verbavatz
JM
,Prohaska
S.
2014
.Автоматическое сшивание осевых линий микротрубочек на серийных электронных томограммах
.PLOS One
9
:e113222.
Wegst
UGK
,Ashby
MF.
2004
.Механическая эффективность природных материалов
.Philos Mag
84
:2167
—86
.Wipfler
B
,Pohl
H
,Яворская
MI
,Beutel
RG.
2016
.Обзор методов анализа структур насекомых — роль морфологии в эпоху филогеномики
.Curr Opin Insect Sci
18
:60
—8
.Заметки автора
© Автор (ы) 2019. Опубликовано Oxford University Press от имени Общества интегративной и сравнительной биологии. Все права защищены. За разрешениями обращайтесь по электронной почте: журналы[email protected].