47. Генотип и фенотип программных систем | by Michael Dubakov
Полгода назад, читая Лема, я набрел на генотипы и фенотипы. И конечно в голову полезли мысли, как это распространить на программные продукты.
Генотип и ФенотипЕсли очень кратко, то генотип закладывает какие-то свойства в систему, а внешняя среда накладывает свой отпечаток и формирует фенотип на основе этих свойств и контекста. То есть отдельная живая особь содержит больше информации, чем генотип, потому что к информации генотипа добавляется внешняя информация среды. Под влиянием внешней среды происходит экспрессия генов. К примеру, под воздействием антибактериального мыла и заботливой мамы у вас развивается склонность к аллергии. В генах это заложено, но вот Васе мама разрешала копаться в грязи, и у него нет аллергии. А вы ненавидите апрель.
Возьмем для конкретики любой продукт. Ну, например, Targetprocess. Он имеет генотип (заложенные в него функции), но каждый клиент настраивает систему под себя, так что каждый отдельный аккаунт Targetprocess — это фенотип.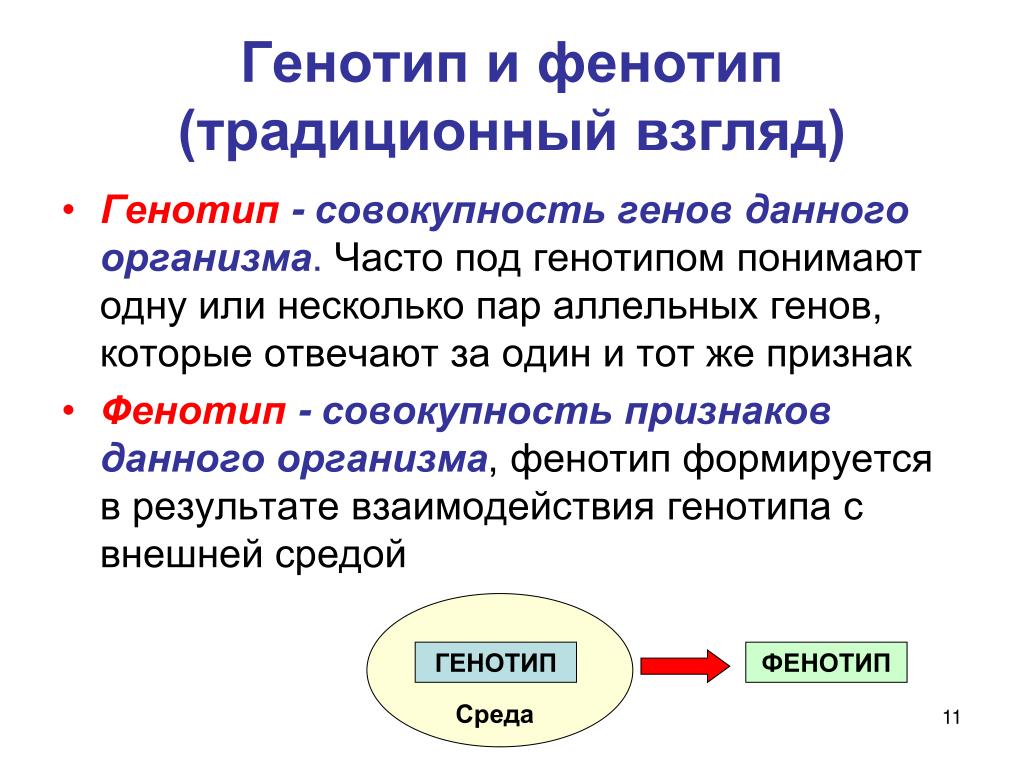 Мы сейчас можем смотреть на ситуацию с эволюционной точки зрения. Генотип Targetprocess порождает множество своих экземпляров, которые сталкиваются с внешней средой (клиентами). В этой внешней среде экземпляры либо выживают (компания пользуется продуктом и все хорошо), либо умирают (компания отказывается от продукта).
Мы сейчас можем смотреть на ситуацию с эволюционной точки зрения. Генотип Targetprocess порождает множество своих экземпляров, которые сталкиваются с внешней средой (клиентами). В этой внешней среде экземпляры либо выживают (компания пользуется продуктом и все хорошо), либо умирают (компания отказывается от продукта).
Внешняя среда (контекст использования) может оказывать существенное влияние на конкретный экземпляр системы. Фенотип накапливает в себе информацию: домен компании, данные, недовольные пользователи и их фидбек, настройки, расширения, интеграции, и тп. В будущем эта накопленная информация по идее может вызывать мутации в генотипе. Фактически, это и есть задача Product Owner —собирать данные о фенотипах и закладывать нужные мутации в генотип продукта. От того, что заложено в генотипе, зависит выживаемость каждого конкретного экземпляра системы, который попал в свой специфический внешний контекст.
Однообразный контекстДальше самое интересное. При однообразии контекстов генотип может быть довольно однозначный, потому что ясно к чему стремиться и что оптимизировать. Продукт используют похожим образом и Product Owner будет получать большое количество одинаковых сигналов от разных экземпляров, поэтому легко понять, что нужно изменить в системе. В предельном случае, если вы делаете систему под одного заказчика, генотип = фенотипу. Вы просто делаете все, что нужно этому заказчику.
При однообразии контекстов генотип может быть довольно однозначный, потому что ясно к чему стремиться и что оптимизировать. Продукт используют похожим образом и Product Owner будет получать большое количество одинаковых сигналов от разных экземпляров, поэтому легко понять, что нужно изменить в системе. В предельном случае, если вы делаете систему под одного заказчика, генотип = фенотипу. Вы просто делаете все, что нужно этому заказчику.
Чуть более сложный случай, когда все ваши клиенты очень похожи. Например, это софт для ведения бухгалтерии ИП. Единый стандарт с минимальными изменениями контекста использования делает жизнь PO довольно простой. Вам достаточно иметь несколько клиентов и поддерживать с ними контакт, чтобы реализовать всю нужную функциональность.
Разнообразный контекст
При разнообразии контекстов все становится гораздо сложнее. Каждый экземпляр имеет уникальный фенотип. Клиенты просят довольно разные вещи, и вы не можете делать однозначные выводы на основе фидбека даже сотни клиентов. Жизнь PO становится невыносимой. Принимать правильные решения невероятно сложно, потому что фича, решающая проблему 10% клиентов, совершенно не нужна оставшимся 90%. Что самое хреновое, очень мало таких фич, которые нужны почти всем. Каждая фича нужна немногим. И вы попадаете в ад.
Жизнь PO становится невыносимой. Принимать правильные решения невероятно сложно, потому что фича, решающая проблему 10% клиентов, совершенно не нужна оставшимся 90%. Что самое хреновое, очень мало таких фич, которые нужны почти всем. Каждая фича нужна немногим. И вы попадаете в ад.
Фактически вы приходите к тому, что при разнообразии контекстов генотип должен быть вариативный. Вы должны обеспечить возможность экспрессии генов. Это значит, что отдельные экземпляры системы должны иметь возможность приспособиться к очень разным контекстам и выжить в непредсказуемой среде. В софте эта вариативность реализуется через обширность настроек или платформу с приложениями. Если вы решили делать софт на рынке с большим разнообразием контекстов — нужно делать систему гибкой. Другого пути просто нет. Сразу скажу, что обширность настроек — очень стремный путь. Рано или поздно продукт просто умрет из-за внутренней сложности.
Если взять Fibery, то мы пошли от предпосылки большого разнообразия контекстов. На самом деле за годы эксплуатации Targetprocess мы выяснили, что компании работают очень разнообразно. Их процессы часто уникальны и выдвигаемые требования хрен реализуешь в рамках одного целостного продукта. Поэтому, если мы хотим быть на рынке систем управления проектами, мы должны заложить в генотип Fibery значительную гибкость.
На самом деле за годы эксплуатации Targetprocess мы выяснили, что компании работают очень разнообразно. Их процессы часто уникальны и выдвигаемые требования хрен реализуешь в рамках одного целостного продукта. Поэтому, если мы хотим быть на рынке систем управления проектами, мы должны заложить в генотип Fibery значительную гибкость.
Что интересно, сам факт закладывания этой гибкости приведет к тому, что Fibery будут чаще использовать в самых непредсказуемых и разнообразных контекстах. Мы получаем очень веселый positive feedback loop:
Гибкая система → неожиданное использование → требование еще большей гибкости.
Пока сложно представить, к чему это все приведет. Но кажется впереди у нас много веселых бесед об эволюции, сложности и качестве 🌶.
МОДИФИКАЦИИ • Большая российская энциклопедия
В книжной версии
Том 20.
 Москва, 2012, стр. 590
Москва, 2012, стр. 590Скопировать библиографическую ссылку:
 Москва, 2012, стр. 590
Москва, 2012, стр. 590Авторы: В. С. Михеев
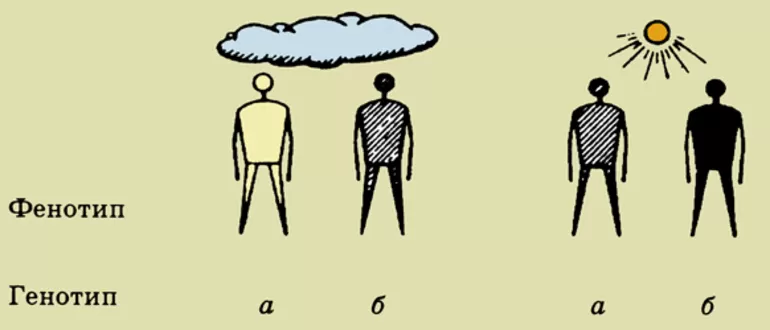
МОДИФИКА́ЦИИ в биологии (позднелатинское modificatio, от лат. modus – мера, вид и facio – делать), спонтанные или вызванные факторами внешней среды изменения признаков организма (его фенотипа), не связанные с изменениями генотипа. Исторически явление М. связывали с «обычными» для существования данного вида организмов факторами внешней среды и подчёркивали адаптивный и обратимый характер М. Так, УФ-лучи вызывают у человека усиление пигментации кожи – загар (причём у разных людей – в разной степени). После прекращения действия лучей загар постепенно исчезает.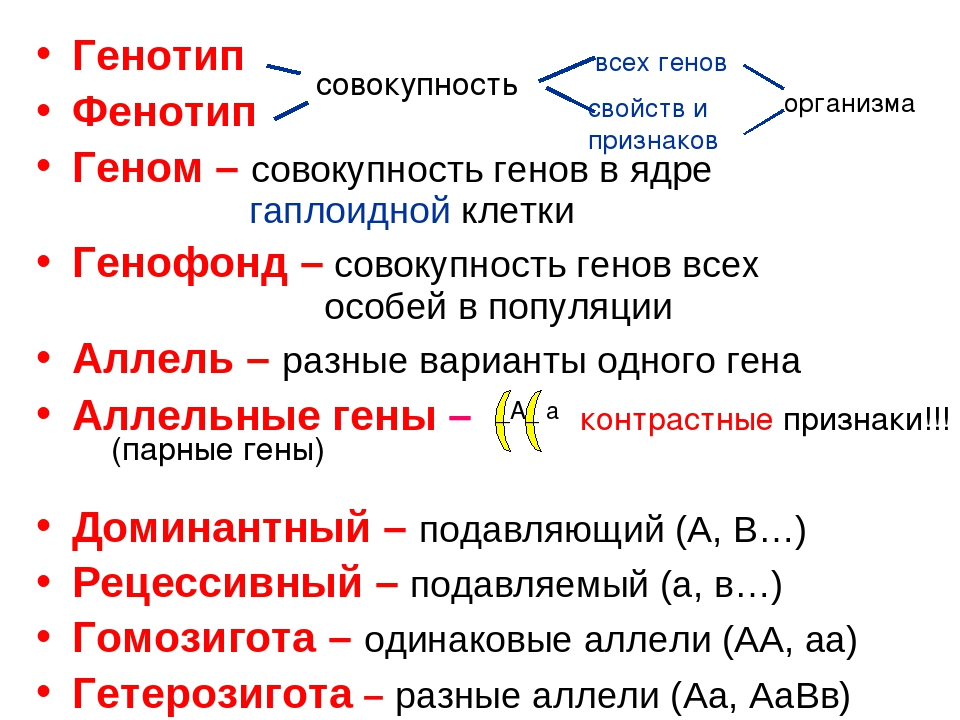
Методы исследования наследственности. Фенотип и генотип
1. Дайте определения понятий.
Гибрид – потомки от скрещивание двух организмов, отличающихся друг от друга одним или несколькими признаками.
Фенотип — совокупность всех внешних и внутренних признаков и свойств организма.
Генотип — совокупность генов, которую организм получает от родителей.
2. Какие методы генетики как науки вам известны?
Цитогенетический метод — заключается в изучении хромосом объекта при помощи микроскопа и позволяющий определить их число и форму. Гибридологический метод – скрещивание организмов.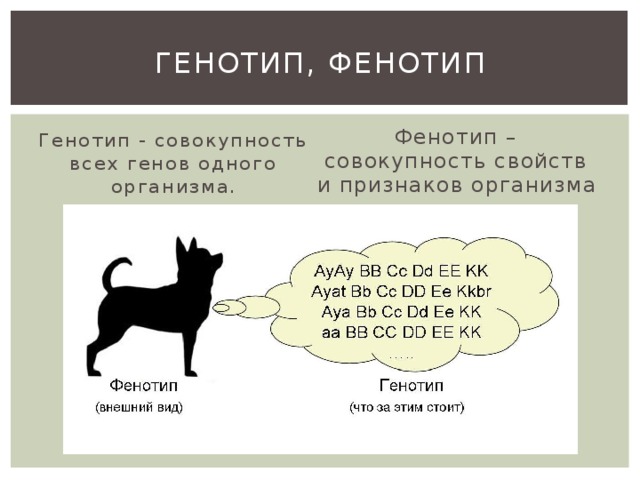 Математический метод – используется при подсчете результатов.
Математический метод – используется при подсчете результатов.
3. В чём состоит сущность гибридологического метода изучения наследования признаков?
Суть гибридологического метода заключается в скрещивании специально подобранных пар организмов и тщательном изучении полученных потомков. Этот метод лежит в основе генетических исследований.
4. Почему горох можно считать удачным объектом генетических исследований?
Существует множество сортов гороха, отличающегося цветом и формой горошин. Горох легко выращивать, он может давать несколько урожаев в год. Горох — самоопыляемое растение, но в эксперименте самоопыление легко предотвратить, и экспериментатор может опылять растение пыльцой с другого растения, т. е. перекрёстно.
5. Наследование каких пар признаков у гороха изучал Г. Мендель?
Он изучал наследование окраски горошин и их формы.
6. Что понимают в генетике под чистой линией?
Чистые линии – организмы, которые при скрещивании всегда воспроизводят один вариант проявления признака. Данные организмы являются гомозиготами по данному признаку.
7. От чего зависит проявление у организма в фенотипе тех или иных признаков, заложенных в генотипе?
Фенотип зависит от сочетания генов в организме.
Полная база данных ассоциаций генотип-фенотип у мышей
Подготовил Илья Скляр
Найти связь гена и фенотипического признака — решить одну из основных задач генетики и молекулярной биологии. Международная группа биоинформатиков решала эту задачу для всех известных на сегодня генов мышек линии BXD. С этой линией мышей работают с 1973 года. Информации о генах, транскриптах и белках за годы исследований накопилось очень много. Причем каждая группа, работавшая с линией BXD, создавала свою вариацию этой линии (например, с определенными выключенными или мутантными генами). Количество новых фенотипов растет экспоненциально, и к декабрю 2016 накопилось более 5000 различных феномов (так биоинформатики называют совокупность всех фенотипов всех вариантов клеток в организме).
Причем каждая группа, работавшая с линией BXD, создавала свою вариацию этой линии (например, с определенными выключенными или мутантными генами). Количество новых фенотипов растет экспоненциально, и к декабрю 2016 накопилось более 5000 различных феномов (так биоинформатики называют совокупность всех фенотипов всех вариантов клеток в организме).
Ученые смогли упорядочить всю совокупность этих феномов в единую базу данных и выявили несколько новых интересных ассоциаций ген-фенотипический признак. Так, было установлено, что ген Rpl26 связан с регуляцией веса тела. Установлена связь между геном Oprm1 и ответом на морфин, Cpt1 и регуляцией липидного метаболизма и т.д.
Для анализа исследователи создали новые программные инструменты, позволяющие сопоставлять разные группы феномов по отдельности. Также по отдельности сравнивали транскриптом и протеом, искали ассоциации феномов и профилей экспрессии генов, проводили обратный анализ белок-ген. Часть использованных программ и исследований находится в открытом доступе на сайте systems-genetics.org.
Часть использованных программ и исследований находится в открытом доступе на сайте systems-genetics.org.
Методология оправдала себя. «Этот новый подход — одна из первых попыток заложить фундамент для новой эры биологии. Доктора скоро смогут практиковать подобные подходы и персонализовать лечение пациентов», — сообщает один из ученых, Иоганн Ауверкс.
Источник
Hao Li et al. // An integrated systems genetics and omics toolkit to probe gene function. // Cell Systems, November 2017. DOI: 10.1016/j.cels.2017.10.016
Добавить в избранноеГЕНОТИП-ФЕНОТИПИЧЕСКИЕ КОРРЕЛЯЦИИ ТЕЧЕНИЯ КИСТОЗНОГО ФИБРОЗА У РОССИЙСКИХ ДЕТЕЙ. ПЕРВОЕ ОПИСАНИЕ ОДИННАДЦАТИ НОВЫХ МУТАЦИЙ | Горинова
1.
2. Farrell P. The prevalence of cystic fibrosis in the European Union. J Cyst Fibros. 2008;7(5):450–453. doi: 10.1016/j.jcf.2008.03.007.
3. Красовский С.А., Черняк А.В., Каширская Н.Ю., и др. Муковисцидоз в России: Создание национального регистра // Педиатрия. Журнал им. Г.Н. Сперанского. — 2014. — Т. 93. — № 4 — С. 44–55.
4. Каширская Н.Ю., Красовский С.А., Черняк А.В., и др. Динамика продолжительности жизни больных муковисцидозом, проживающих в Москве, и ее связь с получаемой терапией: ретроспективный анализ за 1993–2013 гг. // Вопросы современной педиатрии. — 2015. — Т. 14. — № 4 — С. 503–508. doi: 10.15690/vsp.v14.i4.1390.
5. Riordan JR, Rommens JM, Kerem B, et al. Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science. 1989;245(4922):1066–1073. doi: 10.1126/science.2475911.
Riordan JR, Rommens JM, Kerem B, et al. Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science. 1989;245(4922):1066–1073. doi: 10.1126/science.2475911.
7. genet.sickkids.on.ca [Internet]. Cystic Fibrosis Mutation Database [cited 2017 Dec 24]. Available from: www.genet.sickkids. on.ca/cftr.
8. Audrezet MP, Dabricot A, Le Marechal C, Ferec C. Validation of high-resolution DNA melting analysis for mutation scanning of the cystic fibrosis transmembrane conductance regulator (CFTR) gene. J Mol Diagn. 2008;10(5):424–434. doi: 10.2353/jmoldx. 2008.080056.
2008.080056.
9. Le Marechal C, Audrezet MP, Quere I, et al. Complete and rapid scanning of the cystic fibrosis transmembrane conductance regulator (CFTR) gene by denaturing high-performance liquid chromatography (DHPLC): major inplications for genetics counseling. Hum Genet. 2001;108(4):290–298. doi: 10.1007/s004390100490.
10. Galvin P, Clarke L, Harvey S, Amaral M. Microarray analysis in cystic fibrosis. J Cyst Fibros. 2004;3(2):29–33. doi: 10.1016/j.jcf.2004.05.006.
11. Баранов А.А., Капранов Н.И., Каширская Н.Ю., и др. Проблемы диагностики муковисцидоза и пути их решения в России / Педиатрическая фармакология. — 2014. — Т. 11. — № 6 — С. 16–23. doi: 10.15690/pf.v11i6.1211.
12. Lucarelli M, Narzi L, Piergentili R, et al. A 96-well formatted method for exon and exon/intron boundary full sequencing of the CFTR gene. Anal Biochem. 2006;353(2):226–235. doi: 10.1016/j.ab.2006.03.022.
Lucarelli M, Narzi L, Piergentili R, et al. A 96-well formatted method for exon and exon/intron boundary full sequencing of the CFTR gene. Anal Biochem. 2006;353(2):226–235. doi: 10.1016/j.ab.2006.03.022.
13. A new targeted CFTR mutation panel based on next-generation sequencing technology. J Mol Diagn. 2017;19(5):788–800. doi: 10.1016/j.jmoldx.2017.06.002.
14. Schrijver I, Rappahahn K, Pique L, et al. Multiplex ligationdependent probe amplification identification of whole exon and single nucleotide deletions in the CFTR gene of Hispanic individuals with cystic fibrosis. J Mol Diagn. 2008;10(4):368–375. doi: 10.2353/jmoldx.2008.080004.
15. Mehdizadeh Hakkak A, Keramatipour M, Talebi S, et al. Analysis of CFTR gene mutations in children with cystic fibrosis, first report from North-East of Iran. Iran J Basic Med Sci. 2013;16(8):918–921.
Iran J Basic Med Sci. 2013;16(8):918–921.
16. Регистр больных муковисцидозом в Российской Федерации. 2015 год / Под ред. Е.И. Кондратьевой, С.А. Красовского, А.Ю. Воронковой, и др. — М.: Медпрактика-М; 2016. — 72 с.
17. Gurwitz D, Corey M, Francis PW. Perspectives in cystic fibrosis. Pediatr Clin North Am. 1979;26(3):603–615. doi: 10.1016/S00313955(16)33752-X.
18. Rowntree RK, Harris A. The phenotypic consequences of CFTR mutations. Ann Hum Genet. 2003;67(Pt 5):471–485. doi: 10.1046/j.1469-1809.2003.00028.x.
19. Green DM, McDougal KE, Blackman SM, et al. Mutations that permit residual CFTR function delay acquisition of multiple respiratory pathogens in CF patients. Respir Res. 2010;11:140. doi: 10.1186/1465-9921-11-140.
2010;11:140. doi: 10.1186/1465-9921-11-140.
20. Bombieri C, Seia M, Castellani C. Genotypes and phenotypes in cystic fibrosis and cystic fibrosis transmembrane regulator-related disorders. Semin Respir Crit Care Med. 2015;36(2):180–193. doi: 10.1055/s-0035-1547318.
21. Cutting GR. Cystic fibrosis genetics: from molecular understanding to clinical application. Nat Rev Genet. 2015;16(1):45–56. doi: 10.1038/nrg3849.
22. Elborn JS. Personalised medicine for cystic fibrosis: treating the basic defect. Eur Respir Rev. 2013;22(127):3–5. doi: 10.1183/09059180.00008112.
23. Foucher J, Chanteloup E, Vergniol J, et al. Diagnosis of cirrhosis by transient elastography (FibroScan): a prospective study. Gut. 2006;55(3):403–408. doi: 10.1136/gut.2005.069153.
Gut. 2006;55(3):403–408. doi: 10.1136/gut.2005.069153.
24. hgmd.cf.ac.uk [Internet]. HGMD® Professional 2017.4. [cited 2018 Feb 20]. Available from: http://www.hgmd.cf.ac.uk/ac/index.php.
25. Одинокова О.Н. Расширенный поиск мутаций гена CFTR в выборке больных муковисцидозом из Сибирского региона. / VII ежегодная Северо-Западная с международным участием научно-практическая конференция по муковисцидозу «Практика лечения муковисцидоза»; Май 27–28, 2016; Санкт-Петербург. Доступно по: http://ostrovaru.com/%D0%BF%D1%80%D0%BE%D0%B3%D1%80%D0%B0%D0%BC%D0%BC%D1%8B/2015/01/08/%D0%BA%D0%BE%D0%BD%D1%84%D0%B5%D1%80%D0%B5%D0%BD%D1%86%D0%B8%D0%B8%D0%B8-%D1%81%D0%B5%D0%BC%D0%B8%D0%BD%D0%B0%D1%80%D1%8B/. Ссылка активна на 12.02.2018.
26. Sinaasappel M, Stern M, Littlewood J, et al. Nutrition in patients with cystic fibrosis: a European Consensus. J Cyst Fibros. 2002; 1(2):51–75. doi: 10.1016/S1569-1993(02)00032-2.
Nutrition in patients with cystic fibrosis: a European Consensus. J Cyst Fibros. 2002; 1(2):51–75. doi: 10.1016/S1569-1993(02)00032-2.
27. Richards S, Aziz N, Bale S, et al. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet Med. 2015;17(5):405–424. doi: 10.1038/gim.2015.30.
28. Кондратьева Е.И., Шерман В.Д., Амелина Е.Л., и др. Клинико-генетическая характеристика и исходы мекониевого илеуса при муковисцидозе // Российский вестник перинатологии и педиатрии. — 2016. — Т. 61. — № 6 — С. 77–81. doi: 10.21508/1027-4065-2016-61-6-77-81.
Сведения об организации
Мы рады приветствовать Вас на официальном сайтеГосударственного бюджетного образовательного учреждения
Среднего профессионального образования
Магнитогорского Медицинского Колледжа имени П.
 Ф. Надеждина.
Ф. Надеждина.В 2012 году мы отметили свой 80-летний день рождения. История учреждений складывается из множества больших и малых событий, которые становятся неотъемлемой частью жизни его коллектива. Возможно, кто-то усомнится: а нужно ли сегодня вспомнить то, что для истории покажется незначительным?! Однако за историей нашего колледжа, за «малыми» цифрами и фактами не только чья-то судьба, но и история города, история страны. В биографии нашего учебного заведения (как впрочем, во множестве тех, чья история имеет советские истоки), заключен социально-политический урок, о котором стоит говорить. Поступь времени — вот, пожалуй, главное, что отличает события «малого масштаба». Словно маленькие ручейки, они, сливаясь воедино, образовали большую реку. И в этой реке жизни десятки замечательных судеб, годы жизни, отданные выбранному делу.
Магнитогорским медицинским колледжем подготовлено и выпущено на 01.01.2012г 16.919 средних медицинских специалистов, в том числе:
- фельдшеров -6. 580 чел
- санитарных фельдшеров -1.238 чел
- акушерок -1.893 чел
- фельдшеров лаборантов — 604 чел
- медсестер общего профиля- 5.916 чел
- медсестер детских учрежден -198 чел
- медсестер повышен уровня — 80 чел
- зубных врачей -362 чел
- зубных техников -133 чел
- фармацевтов — 99 чел
 580 чел
580 челВыпускники ММК им.Надеждина работают во всех лечебно-профилактических учреждениях города Магнитогорска, Челябинской области, России и за её пределами. Многие из них стали врачами, окончив высшие медицинские учреждения, ведущими специалистами, организаторами здравоохранения, главными и старшими медсестрами отделений, заведующими фельдшерско-акушерскими здравпунктами и здравпунктами предприятий.
Восемьдесят лет.… Это целая жизнь, если речь идёт о человеке. Это молодость, если речь идёт об учебном заведении. Это солидный период в жизни большого дружного коллектива, который и сотрудники, и выпускники, и студенты считают своей второй семьёй. ..
..
Основные сведения
Структура и органы управления образовательной организацией
Документы
Образование
Образовательные стандарты
Руководство. Научно-педагогический состав
Материально-техническое обеспечение
Стипендии и иные виды материальной поддержки
Платные образовательные услуги
Финансово-хозяйственная деятельность
Вакантные места для приёма (перевода)
Metris — Фенотипирование
Фенотип – (от греческого слова phaino- являю, обнаруживаю) совокупность характеристик, присущих индивиду на определенной стадии развития. Фенотип формируется на основе генотипа, опосредованного рядом внешнесредовых факторов. Генотип организма – унаследованные инструкции, которые он несет в своем генетическом коде.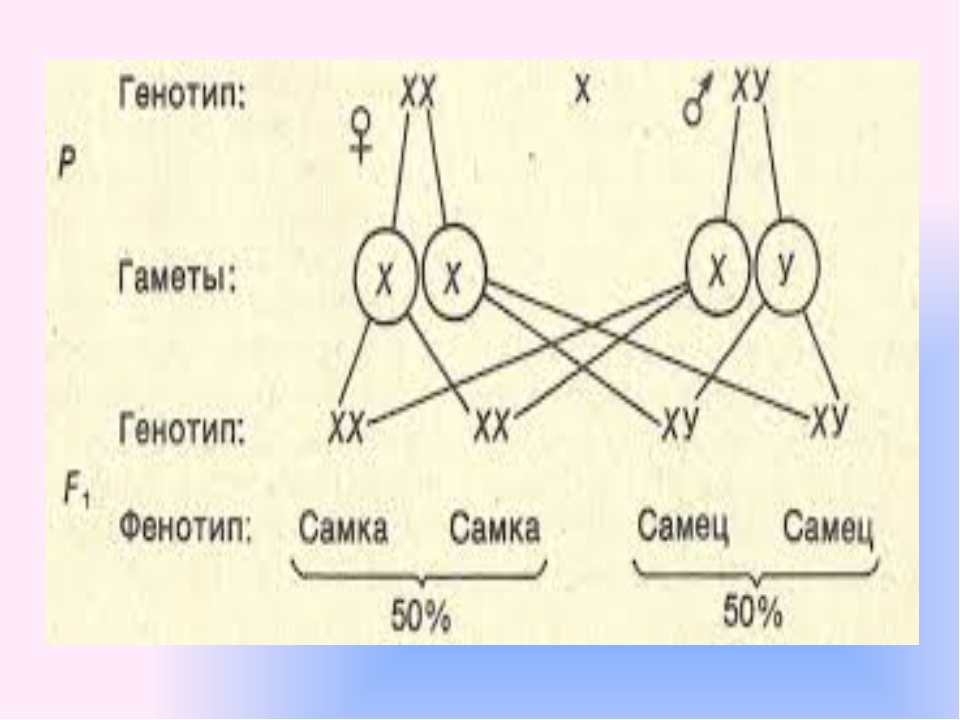 Не все организмы с тем же самым генотипом выглядят или ведут себя одинаковым образом, так как внешний вид и поведение модифицируются условиями окружающей среды и условиями развития. Таким же образом, не все организмы, которые похожи друг на друга, обязательно имеют одинаковый генотип. (Определение из Википедии).
Не все организмы с тем же самым генотипом выглядят или ведут себя одинаковым образом, так как внешний вид и поведение модифицируются условиями окружающей среды и условиями развития. Таким же образом, не все организмы, которые похожи друг на друга, обязательно имеют одинаковый генотип. (Определение из Википедии).
Метрис предлагает несколько путей исследования фенотипов лабораторных животных управляемым автоматизированным способом, который также является эффективным и воспроизводимым. Система LABORAS способна обеспечить данные для исследования долгосрочных поведенческих реакций, которые включают в себя нормальные поведения, стереотипные поведения, а также социальные поведение (используя специальный модуль «Sociality клетки» системы LABORAS). В отличие от большинства других систем, основанных на “зависимых” параметрах видео отслеживания (полученные из позиции животного), LABORAS обеспечивает ряд независимых параметров, полученных из энергии вызванной мышечной активностью частей тела (челюсти, головы, передних лап, задних конечностей, тела и т. д.).
д.).
Кроме того Метрис предлагает функциональные возможности для анализа ультразвуковых вокализаций (USV звуки) лабораторных животных, тем самым добавляя важное число параметров, которые могут быть использованы для фенотипирования животного.
Все системы созданы для долгосрочного, высоко-пропускного тестирования, путем сбора важных фенотипических параметров многочисленных животных в течение длительного испытательного периода. Конечные итоги предоставляются компактным и удобным для пользователя способом. Все это делает оборудование очень подходящим для фенотипирования.
Генотип против фенотипа: примеры и определения
Любой организм является побочным продуктом как его генетической структуры, так и окружающей среды. Чтобы понять это в деталях, мы должны сначала оценить некоторые основные генетические термины и концепции. Здесь мы даем определения терминов генотип и фенотип , обсуждаем их взаимосвязь и рассматриваем, почему и как мы могли бы их изучить.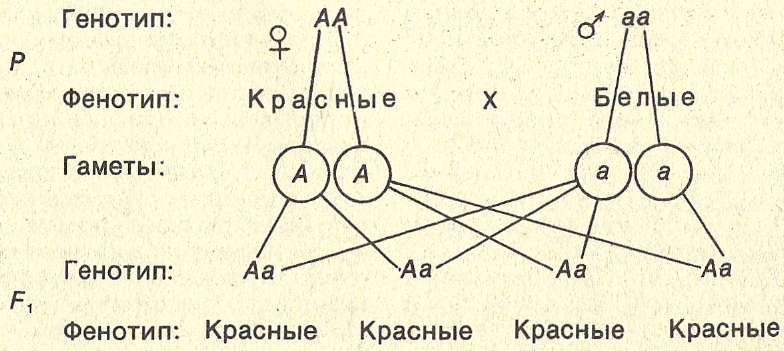
В биологии ген — это часть ДНК, которая кодирует признак.Точное расположение нуклеотидов (каждый из которых состоит из фосфатной группы, сахара и основания) в гене может различаться в разных копиях одного и того же гена. Следовательно, у разных организмов ген может существовать в разных формах. Эти разные формы известны как аллели. Точное фиксированное положение на хромосоме, которое содержит конкретный ген, называется локусом.
Диплоидный организм наследует либо две копии одного и того же аллеля, либо одну копию двух разных аллелей от своих родителей. Если человек наследует два идентичных аллеля, его генотип считается гомозиготным по этому локусу.
Однако, если они обладают двумя разными аллелями, их генотип классифицируется как гетерозиготный для этого локуса. Аллели одного и того же гена могут быть аутосомно-доминантными или рецессивными. Аутосомно-доминантный аллель всегда будет предпочтительно выражаться по сравнению с рецессивным аллелем.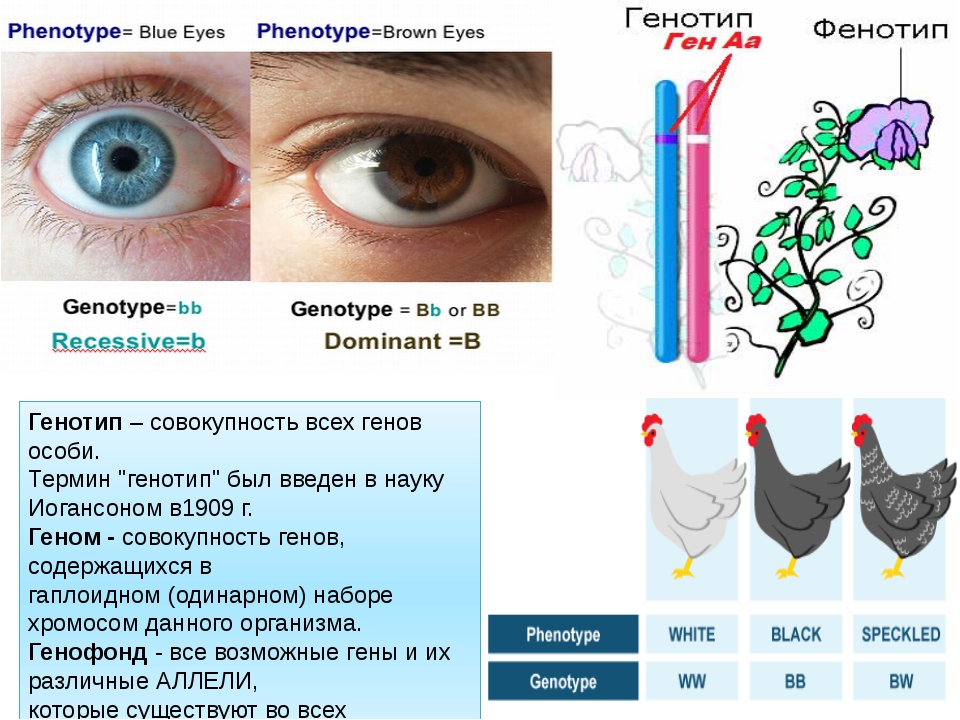
Последующая комбинация аллелей, которыми обладает человек для определенного гена, — это его генотип .
Рассмотрим классический пример — цвет глаз.
- Ген кодирует цвет глаз.
- В этом примере аллель либо коричневый, либо синий, причем один унаследован от матери, а другой — от отца.
- Коричневый аллель является доминантным (B), а синий аллель — рецессивным (b). Если ребенок наследует два разных аллеля (гетерозиготных), то у него будут карие глаза. Чтобы у ребенка были голубые глаза, они должны быть гомозиготными по аллелю голубого глаза.
Рис. 1. Диаграмма наследования, в которой подробно показано, как индивид может унаследовать голубые или карие глаза в зависимости от аллелей, принадлежащих их родителям, причем аллель цвета карих глаз является доминирующим, а аллель цвета голубых глаз — рецессивным.
Другие примеры генотипа включают:
- Цвет волос
- Рост
- Размер обуви
Сумма наблюдаемых характеристик организма — это их фенотип.
 Ключевое различие между фенотипом и генотипом состоит в том, что, хотя генотип наследуется от родителей организма, фенотип — нет.
Ключевое различие между фенотипом и генотипом состоит в том, что, хотя генотип наследуется от родителей организма, фенотип — нет.Хотя фенотип влияет на генотип, генотип не равен фенотипу.На фенотип влияет генотип и факторы, включая:
- Эпигенетические модификации
- Факторы окружающей среды и образа жизни
Рисунок 2: Фламинго от природы имеют белый цвет, их вызывают только пигменты организмов, которые они едят стать ярко-розовым.
Примеры фенотипов
Факторы окружающей среды, которые могут влиять на фенотип, включают питание, температуру, влажность и стресс.Фламинго — классический пример того, как окружающая среда влияет на фенотип. Несмотря на то, что они известны своим ярко-розовым цветом, их естественный цвет белый — розовый цвет вызван пигментами организмов в их рационе.
Второй пример — цвет кожи человека. Наши гены контролируют количество и тип меланина, который мы производим, однако воздействие ультрафиолетового света в солнечном климате вызывает потемнение существующего меланина и способствует усилению меланогенеза и, следовательно, более темной коже.
Наблюдать за фенотипом просто — мы смотрим на внешние особенности и характеристики организма и делаем выводы о них.Однако наблюдение за генотипом немного сложнее.
Генотипирование — это процесс, при котором различия в генотипе человека анализируются с помощью биологических анализов. Полученные данные затем можно сравнить либо с последовательностью второго человека, либо с базой данных последовательностей.
Ранее генотипирование позволяло получать только частичные последовательности. Теперь, благодаря крупным технологическим достижениям последних лет, мы получили самое современное секвенирование всего генома.
Рис. 3. Рабочий процесс, изображающий различные этапы секвенирования всего генома (WGS).
(WGS) позволяет получать целые последовательности. WGS — это эффективный процесс, который становится все более доступным и включает использование высокопроизводительных методов секвенирования, таких как секвенирование одной молекулы в реальном времени (SMRT), для идентификации исходной последовательности нуклеотидов, составляющих ДНК организма.
WGS — это не единственный способ анализа генома организма. Доступны различные методы.
Почему важно изучать генотип и фенотип?Понимание взаимосвязи между генотипом и фенотипом может быть чрезвычайно полезным в различных областях исследований.
Особенно интересная область — фармакогеномика. Генетические вариации могут возникать в ферментах печени, необходимых для метаболизма лекарств, таких как CYP450. Таким образом, фенотип человека, то есть его способность метаболизировать конкретное лекарство, может варьироваться в зависимости от того, какой формой гена, кодирующего фермент, он обладает. Для фармацевтических компаний и врачей эти знания являются ключевыми для определения рекомендуемых дозировок лекарств для разных групп населения.
Использование методов генотипирования и фенотипирования в тандеме кажется лучше, чем использование только генотипических тестов.В сравнительном клиническом исследовании фармакогеномики мультиплексный подход выявил большие различия в способности метаболизма лекарств, чем это было предсказано только генотипированием. Это имеет важное значение для персонализированной медицины и подчеркивает необходимость проявлять осторожность, полагаясь исключительно на генотипирование.
Это имеет важное значение для персонализированной медицины и подчеркивает необходимость проявлять осторожность, полагаясь исключительно на генотипирование.
Используя модели животных, например мышей, ученые могут генетически модифицировать организм так, чтобы он больше не экспрессировал определенный ген — известные как «нокаутные мыши».Сравнивая фенотип этого животного с фенотипом дикого типа (то есть фенотипом, который существует, когда ген не был удален), мы можем изучить роль определенных генов в доставке определенных фенотипов.
Инициатива в области информатики генома мышей (MGI) составила базу данных тысяч фенотипов, которые могут быть созданы и изучены, а также генов, которые должны быть выбиты для получения каждого конкретного фенотипа.
| | Генотип | набор генов в нашей ДНК, которые отвечают за определенный признак | Наблюдаемые характеристики и признаки организма | |
| Характеризуется | Методы генотипирования, такие как WGS | Наблюдение за внешними характеристиками организма | ||
| Зависит от | Последовательности генов, которыми обладает организм | Генотип, ПЛЮС эпигенетика и факторы окружающей среды | ||
| Унаследовано? | Да | Нет | ||
| Пример | Гены, кодирующие цвет глаз | Человек с карими глазами |
Фенотипы и генотипы — основы биологии
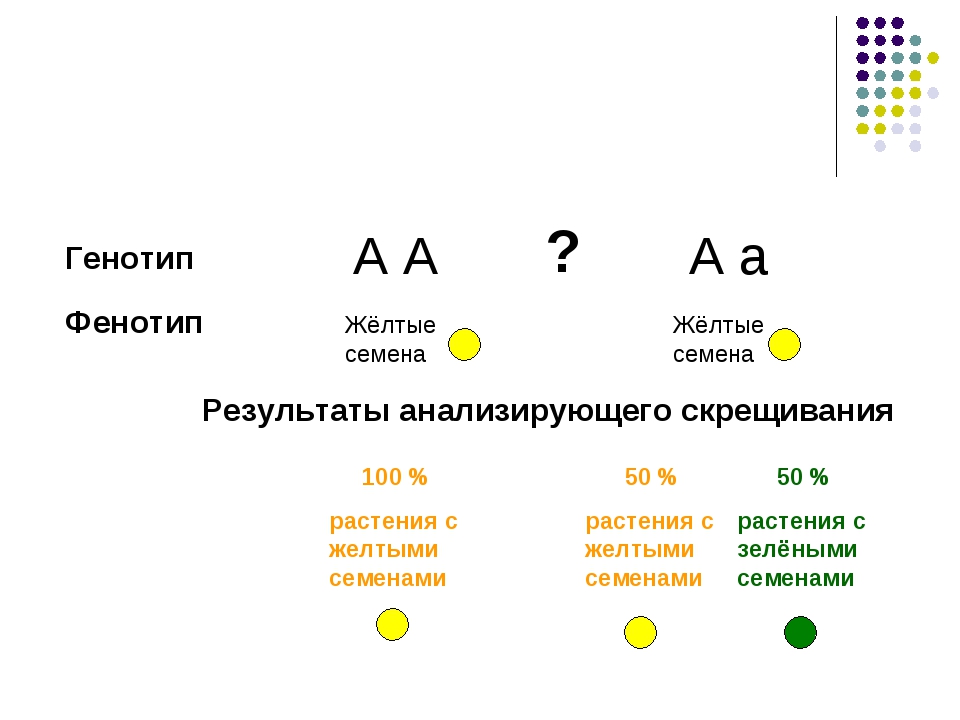
Два аллеля для данного гена в диплоидном организме экспрессируются и взаимодействуют, создавая физические характеристики. Наблюдаемые признаки, выраженные организмом, обозначаются как его фенотип . Основная генетическая структура организма, состоящая как из физически видимых, так и из невыраженных аллелей, называется его генотипом . Эксперименты Менделя по гибридизации демонстрируют разницу между фенотипом и генотипом. Например, фенотипы, которые Мендель наблюдал при скрещивании растений гороха с разными признаками, связаны с диплоидными генотипами растений в поколениях P, F1 и F2.В качестве примера мы будем использовать вторую черту, которую исследовал Мендель, — цвет семян. Цвет семян определяется одним геном с двумя аллелями. Аллель желтого семени является доминантным, а аллель зеленого семени — рецессивным. При перекрестном оплодотворении истинно племенных растений, в которых у одного родителя были желтые семена, а у другого — зеленые семена, все гибридное потомство F1 имело желтые семена. То есть гибридное потомство фенотипически было идентично истинному родителю с желтыми семенами.
Наблюдаемые признаки, выраженные организмом, обозначаются как его фенотип . Основная генетическая структура организма, состоящая как из физически видимых, так и из невыраженных аллелей, называется его генотипом . Эксперименты Менделя по гибридизации демонстрируют разницу между фенотипом и генотипом. Например, фенотипы, которые Мендель наблюдал при скрещивании растений гороха с разными признаками, связаны с диплоидными генотипами растений в поколениях P, F1 и F2.В качестве примера мы будем использовать вторую черту, которую исследовал Мендель, — цвет семян. Цвет семян определяется одним геном с двумя аллелями. Аллель желтого семени является доминантным, а аллель зеленого семени — рецессивным. При перекрестном оплодотворении истинно племенных растений, в которых у одного родителя были желтые семена, а у другого — зеленые семена, все гибридное потомство F1 имело желтые семена. То есть гибридное потомство фенотипически было идентично истинному родителю с желтыми семенами.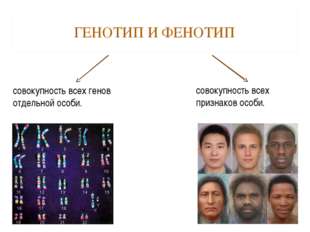 Однако мы знаем, что аллель, подаренный родителем с зелеными семенами, не был просто утерян, потому что он снова появился у некоторых потомков F2 ( Рисунок 5 ).Следовательно, растения F1 должны были генотипически отличаться от родителя с желтыми семенами.
Однако мы знаем, что аллель, подаренный родителем с зелеными семенами, не был просто утерян, потому что он снова появился у некоторых потомков F2 ( Рисунок 5 ).Следовательно, растения F1 должны были генотипически отличаться от родителя с желтыми семенами.
Растения P, которые Мендель использовал в своих экспериментах, были гомозиготными по признаку, который он изучал. Диплоидные организмы, которые являются гомозиготными по по гену, имеют два идентичных аллеля, по одному на каждой из их гомологичных хромосом. Генотип часто записывается как YY или yy , где каждая буква представляет собой один из двух аллелей в генотипе. Доминантный аллель пишется заглавными буквами, а рецессивный аллель — строчными буквами.Буква, используемая для обозначения гена (в данном случае цвет семян), обычно связана с доминантным признаком (желтым аллелем в данном случае или « Y »). Родительские растения гороха Менделя всегда воспроизводили истинно, потому что обе продуцированные гаметы несли один и тот же аллель. Когда P-растения с контрастирующими признаками были перекрестно оплодотворены, все потомство было гетерозиготным по контрастному признаку, что означает, что их генотип имел разные аллели исследуемого гена. Например, желтые растения F1, которые получили аллель Y от своего желтого родителя и аллель y от своего зеленого родителя, имели генотип Yy .
Когда P-растения с контрастирующими признаками были перекрестно оплодотворены, все потомство было гетерозиготным по контрастному признаку, что означает, что их генотип имел разные аллели исследуемого гена. Например, желтые растения F1, которые получили аллель Y от своего желтого родителя и аллель y от своего зеленого родителя, имели генотип Yy .
Рисунок 5: Фенотипы — это физические выражения признаков, которые передаются аллелями. Заглавные буквы обозначают доминантные аллели, а строчные буквы — рецессивные аллели. Фенотипические отношения — это отношения видимых характеристик. Генотипические соотношения — это соотношения комбинаций генов в потомстве, и они не всегда различимы по фенотипам.
Наше обсуждение гомозиготных и гетерозиготных организмов подводит нас к тому, почему гетерозиготное потомство F1 было идентично одному из родителей, а не экспрессировало оба аллеля.По всем семи характеристикам растений один из двух контрастирующих аллелей был доминантным, а другой — рецессивным. Мендель назвал доминантный аллель выраженным единичным фактором; рецессивный аллель был назван фактором латентной единицы. Теперь мы знаем, что эти так называемые единичные факторы на самом деле являются генами на гомологичных хромосомах. Для гена, который экспрессируется по доминантному и рецессивному типу, гомозиготные доминантные и гетерозиготные организмы будут выглядеть одинаково (то есть, они будут иметь разные генотипы, но один и тот же фенотип), а рецессивный аллель будет наблюдаться только у гомозиготных рецессивных особей ( Таблица 1 ).
Мендель назвал доминантный аллель выраженным единичным фактором; рецессивный аллель был назван фактором латентной единицы. Теперь мы знаем, что эти так называемые единичные факторы на самом деле являются генами на гомологичных хромосомах. Для гена, который экспрессируется по доминантному и рецессивному типу, гомозиготные доминантные и гетерозиготные организмы будут выглядеть одинаково (то есть, они будут иметь разные генотипы, но один и тот же фенотип), а рецессивный аллель будет наблюдаться только у гомозиготных рецессивных особей ( Таблица 1 ).
Таблица 1: Соответствие между генотипом и фенотипом для доминантно-рецессивного признака.
| Гомозиготный | Гетерозигота | Гомозиготный | |
| Генотип | ГГ | Гг | гг |
| Фенотип | желтый | желтый | зеленый |
Закон доминирования Менделя гласит, что в гетерозиготе один признак будет скрывать присутствие другого признака того же признака. Например, при скрещивании истинно-размножающихся растений с фиолетовыми цветками с настоящими размножающимися растениями с белыми цветками все потомки были с фиолетовыми цветками, хотя все они имели один аллель для фиолетового и один аллель для белого. Вместо обоих аллелей, вносящих вклад в фенотип, будет выражаться исключительно доминантный аллель. Рецессивный аллель останется латентным, но будет передаваться потомству таким же образом, как и доминантный аллель. Рецессивный признак будет выражен только потомством, которое имеет две копии этого аллеля (, рис. 6, ), и это потомство будет воспроизводиться истинно при самокрещении.
Например, при скрещивании истинно-размножающихся растений с фиолетовыми цветками с настоящими размножающимися растениями с белыми цветками все потомки были с фиолетовыми цветками, хотя все они имели один аллель для фиолетового и один аллель для белого. Вместо обоих аллелей, вносящих вклад в фенотип, будет выражаться исключительно доминантный аллель. Рецессивный аллель останется латентным, но будет передаваться потомству таким же образом, как и доминантный аллель. Рецессивный признак будет выражен только потомством, которое имеет две копии этого аллеля (, рис. 6, ), и это потомство будет воспроизводиться истинно при самокрещении.
Рисунок 6: Аллель альбинизма, выраженный здесь у людей, является рецессивным. Оба родителя этого ребенка несли рецессивный аллель.
Если не указано иное, изображения на этой странице лицензированы OpenStax по лицензии CC-BY 4.0.
OpenStax, Биология. OpenStax CNX. 27 мая 2016 г. http://cnx.org/contents/GFy_h8cu@10.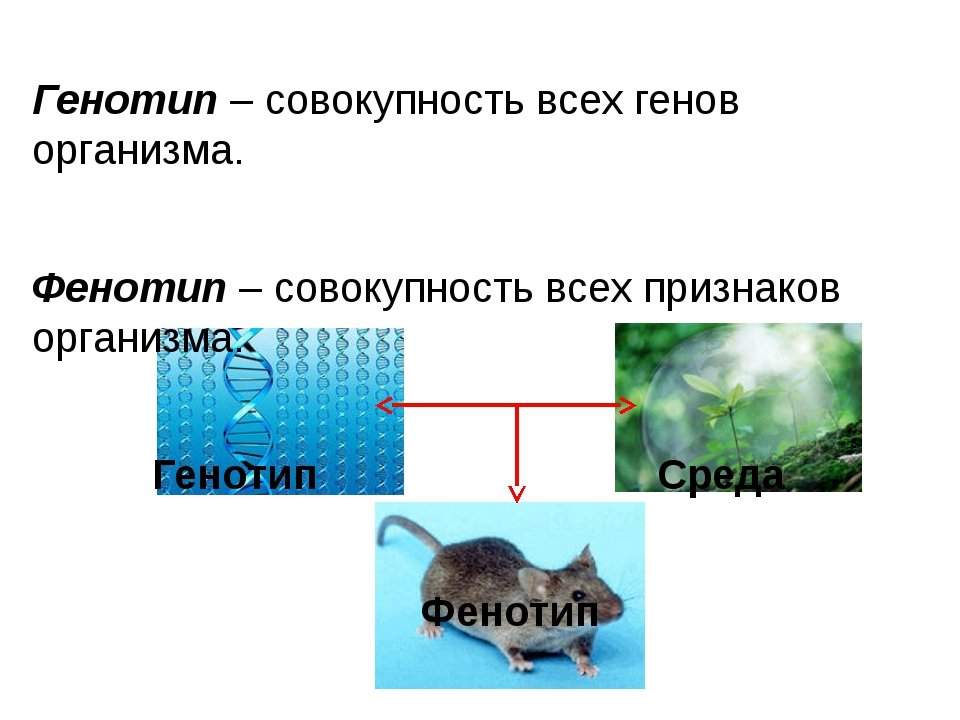 57:4qg08nt-@8/Characteristics-and-Traits
57:4qg08nt-@8/Characteristics-and-Traits
Связывание фенотипа, генотипа и окружающей среды для раскрытия генетических компонентов, лежащих в основе холодоустойчивости прибрежной пихты Дугласовой (Pseudotsuga menziesii var.menziesii)
Эйткен С.Н., Адамс В.Т. (1996) Генетика осенне-зимней холодоустойчивости прибрежной пихты Дугласа в Орегоне. Can J For Res 26 (10): 1828–1837. https://doi.org/10.1139/x26-208
Артикул Google ученый
Эйткен С.Н., Йеман С., Холлидей Дж. А., Ван Т.Л., Кертис-Маклейн С. (2008) Адаптация, миграция или искоренение: последствия изменения климата для популяций деревьев. Evol Appl 1 (1): 95–111.https://doi.org/10.1111/j.1752-4571.2007.00013.x
Артикул PubMed PubMed Central Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Альберто Ф.Дж., Айткен С.Н., Алия Р., Гонсалес-Мартинес С.К., Ханнинен Х., Кремер А., Лефевр Ф., Ленорман Т., Йеман С., Веттен Р., Саволайнен О (2013). Потенциал эволюционного реагирования на изменение климата — доказательства из древесных популяций. Глоб Чанг Биол 19 (6): 1645–1661. https://doi.org/10.1111/gcb.12181
Артикул PubMed PubMed Central Google ученый
Арора Р., Агарвал П., Рэй С., Сингх А.К., Сингх В.П., Тьяги А.К., Капур С. (2007) Семейство генов MADS-бокса у риса: полногеномная идентификация, организация и профили экспрессии во время репродуктивного развития и стресса. BMC Genomics 8 (1): 242–242. https://doi.org/10.1186/1471-2164-8-242
Артикул PubMed PubMed Central Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Bansal S, St Clair JB, Harrington CA, Gould PJ (2015) Влияние изменения климата на холодоустойчивость пихты Дугласовой ( Pseudotsuga menziesii ): экологические и генетические соображения.Глоб Чанг Биол 21 (10): 3814–3826. https://doi.org/10.1111/gcb.12958
Артикул PubMed Google ученый
Barrett RDH, Hoekstra HE (2011) Молекулярные пеленки: тесты адаптации на генетическом уровне. Нат Рев Генет 12 (11): 767–780. https://doi.org/10.1038/nrg3015
CAS Статья PubMed Google ученый
Бек Э. Х., Хайм Р., Хансен Дж. (2004) Устойчивость растений к холодному стрессу: механизмы и сигналы окружающей среды, запускающие морозное затвердевание и рассыхание. J Biosci 29 (4): 449–459. https://doi.org/10.1007/BF02712118
J Biosci 29 (4): 449–459. https://doi.org/10.1007/BF02712118
Артикул PubMed Google ученый
Benjamini Y, Hochberg Y (1995) Контроль уровня ложного обнаружения — практичный и эффективный подход к множественному тестированию. J Roy Stat Soc Ser B-Methodol 57: 289–300
Google ученый
Berg JJ, Coop G (2014) Популяционный генетический сигнал полигенной адаптации.PLoS Genet 10 (8): e1004412. https://doi.org/10.1371/journal.pgen.1004412
Артикул PubMed PubMed Central Google ученый
Bigras FJ, S Colombo 2001 Холодостойкость хвойных, Дордрехт, DOI: https://doi. org/10.1007/978-94-015-9650-3
org/10.1007/978-94-015-9650-3
Bragg JG, Supple MA, Andrew RL, Borevitz JO (2015) Геномные вариации в разных ландшафтах: идеи и приложения.Новый Фитол. 207 (4): 953–967. https://doi.org/10.1111/nph.13410
Артикул PubMed Google ученый
Порода М.Ф., Оттуэлл К.М., Гарднер М.Г., Лоу А.Дж. (2011) Уточнение мер адаптации к изменению климата для разбросанных деревьев в измененных ландшафтах. J Appl Ecol 48 (3): 637–641. https://doi.org/10.1111/j.1365-2664.2011.01969.x
Артикул Google ученый
Chowdhury MEK, Choi B, Cho BK, Kim JB, Park SU, Natarajan S, Lim HS, Bae H (2013) Регулирование 4CL, кодирующее 4-кумарат: коэнзим A лигаза, экспрессия в кенафе в различных стрессовых условиях . Завод OMICS 6: 254–262
Завод OMICS 6: 254–262
CAS Google ученый
Colaneri AC, Jones AM (2014) Схема подключения для сигнализации предприятия G. Curr Opin Plant Biol 22: 56–64. https://doi.org/10.1016/j.pbi.2014.09.004
CAS Статья PubMed PubMed Central Google ученый
Coop G, Witonsky D, Di Rienzo A, Pritchard JK (2010) Использование экологических корреляций для определения локусов, лежащих в основе местной адаптации.Генетика 185 (4): 1411–1423. https://doi.org/10.1534/genetics.110.114819
CAS Статья PubMed PubMed Central Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>De Kort H, Vandepitte K, Bruun HH, Closset-Kopp D, Honnay O, Mergeay J (2014) Пейзажная геномика и обычное садовое испытание показывают адаптивную дифференциацию к температуре по всей Европе у древесных видов Alnus glutinosa . Мол Экол 23 (19): 4709–4721.https://doi.org/10.1111/mec.12813
Артикул PubMed Google ученый
de Villemereuil P, Frichot É, Bazin É, François O, Gaggiotti OE (2014) Методы сканирования генома по более сложным моделям: когда и насколько мы должны им доверять? Мол Экол 23 (8): 2006–2019. https://doi.org/10.1111/mec.12705
Артикул PubMed Google ученый
Дэн В., Касао М. К., Ван П., Сато К., Хейс П.М., Финнеган Э.Дж., Треваскис Б. (2015) Прямая связь между реакцией яровизации и другими ключевыми характеристиками зерновых культур.Нац Коммуна 6: 5882. https://doi.org/10.1038/ncomms6882
К., Ван П., Сато К., Хейс П.М., Финнеган Э.Дж., Треваскис Б. (2015) Прямая связь между реакцией яровизации и другими ключевыми характеристиками зерновых культур.Нац Коммуна 6: 5882. https://doi.org/10.1038/ncomms6882
Артикул PubMed Google ученый
Eckert A, Maloney P, Vogler D, Jensen C, Mix A, Neale D (2015) Локальная адаптация в мелких пространственных масштабах: пример из сахарной сосны ( Pinus lambertiana , Pinaceae). Tree Genet Genomes 11: 1–17
Статья Google ученый
Eckert AJ, Bower AD, González-Martinez SC, Wegrzyn JL, Coop G, Neale DB (2010) Назад к природе: экологическая геномика долькой сосны ( Pinus taeda , Pinaceae).Мол Экол 19 (17): 3789–3805. https://doi.org/10.1111/j.1365-294X.2010.04698.x
https://doi.org/10.1111/j.1365-294X.2010.04698.x
CAS Статья PubMed Google ученый
Eckert AJ, Bower AD, Wegrzyn JL, Pande B, Jermstad KD, Krutovsky KV, Clair JBS, Neale DB (2009a) Генетика ассоциации прибрежной пихты Дугласа ( Pseudotsuga menziesu varaceii. menaceii. I. Признаки, связанные с холодоустойчивостью. Генетика 182 (4): 1289–1302.https://doi.org/10.1534/genetics.109.102350
CAS Статья PubMed PubMed Central Google ученый
Eckert AJ, Wegrzyn JL, Pande B, Jermstad KD, Lee JM, Liechty JD, Tearse BR, Krutovsky KV, Neale DB (2009b) Мультилокусные паттерны нуклеотидного разнообразия и дивергенции показывают положительный отбор генов-кандидатов, связанных с морозоустойчивостью в прибрежной пихте Дугласа ( Pseudotsuga menziesii var. menziesii ). Генетика 183 (1): 289–298. https://doi.org/10.1534/genetics.109.103895
menziesii ). Генетика 183 (1): 289–298. https://doi.org/10.1534/genetics.109.103895
Артикул PubMed PubMed Central Google ученый
Эдмондс Калифорния, Лилли А.С., Кавалли-Сфорца Л.Л. (2004) Мутации, возникающие во фронте волны расширяющейся популяции. Proc Natl Acad Sci U S A 101 (4): 975–979. https://doi.org/10.1073/pnas.0308064100
CAS Статья PubMed PubMed Central Google ученый
Falconer DS, TFC Mackay 1996 Введение в количественную генетику
Fins L, ST Friedman, JV Brotschol 1992 Справочник по количественной генетике лесов, Дордрехт, DOI: https: // doi. org / 10.1007 / 978-94-015-7987-2
org / 10.1007 / 978-94-015-7987-2
Forester BR, JR Lasky, HH Wagner, DL Urban 2017 Использование ассоциаций генотипа с окружающей средой для определения мультилокусной локальной адаптации. bioRxiv: 1–24. DOI: https://doi.org/10.1101/129460
Fournier-Level A, Korte A, Cooper MD, Nordborg M, Schmitt J, Wilczek AM (2011) Карта местной адаптации Arabidopsis thaliana . Наука 334 (6052): 86–89. https://doi.org/10.1126 / наука.1209271
CAS Статья PubMed Google ученый
Frichot E, Schoville SD, Bouchard G, François O (2013) Тестирование ассоциаций между локусами и градиентами окружающей среды с использованием смешанных моделей латентных факторов. Mol Biol Evol 30 (7): 1687–1699. https://doi.org/10.1093/molbev/mst063
Mol Biol Evol 30 (7): 1687–1699. https://doi.org/10.1093/molbev/mst063
CAS Статья PubMed PubMed Central Google ученый
Friendly M (2002) Corrgrams: поисковые дисплеи для корреляционных матриц.Am Stat 56 (4): 316–324. https://doi.org/10.1198/000313002533
Артикул Google ученый
Gao F, Zhou Y, Zhu W, Li X, Fan L, Zhang G (2009) Протеомный анализ белков, реагирующих на холодовой стресс, в розеточных листьях Thellungiella . Planta 230 (5): 1033–1046. https://doi.org/10.1007/s00425-009-1003-6
CAS Статья PubMed Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Garay-Arroyo A, Colmenero-Flores JM, Garciarrubio A, Covarrubias AA (2000) Высокогидрофильные белки у прокариот и эукариот обычны в условиях дефицита воды.J Biol Chem 275 (8): 5668–5674. https://doi.org/10.1074/jbc.275.8.5668
CAS Статья PubMed Google ученый
Гонсалес-Мартинес С.К., Крутовский К.В., Нил Д.Б. (2006) Геномика популяций лесных деревьев и адаптивная эволюция. Новый Phytol 170 (2): 227–238. https://doi.org/10.1111/j.1469-8137.2006.01686.x
Артикул PubMed Google ученый
Гуак С., Олсызк Д.М., Фучигами Л.Х., Тинги Д.Т. (1998) Влияние повышенного содержания CO (2) и температуры на морозостойкость, весеннее распускание и рост почек у пихты Дугласовой (Pseudotsuga menziesii). Tree Physiol 18 (10): 671–679. https://doi.org/10.1093/treephys/18.10.671
Tree Physiol 18 (10): 671–679. https://doi.org/10.1093/treephys/18.10.671
Артикул PubMed Google ученый
Günther T, Coop G (2013) Надежная идентификация локальной адаптации по частотам аллелей. Генетика 195 (1): 205–220. https://doi.org/10.1534/genetics.113.152462
Артикул PubMed PubMed Central Google ученый
Hancock AM, Brachi B, Faure N, Horton MW, Jarymowycz LB, Sperone FG, Toomajian C, Roux F, Bergelson J (2011) Адаптация к климату в геноме Arabidopsis thaliana .Наука 334 (6052): 83–86. https://doi.org/10.1126/science.1209244
CAS Статья PubMed Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Ханна М.А., Хейер А.Г., Хинча Д.К. (2005) Глобальный обзор регуляции генов во время акклиматизации к холоду у Arabidopsis thaliana . PLoS Genet 1 (2): e26. https://doi.org/10.1371/journal.pgen.0010026
Артикул PubMed PubMed Central Google ученый
Харриссон К.А., Павлова А., Телонис-Скотт М., Саннакс П. (2014) Использование геномики для характеристики эволюционного потенциала сохранения диких популяций.Evol Appl. 7 (9): 1008–1025. https://doi.org/10.1111/eva.12149
Артикул PubMed PubMed Central Google ученый
Hijmans RJ, Parra JL, Jones PG, Jarvis A (2005) Интерполированные климатические поверхности с очень высоким разрешением для глобальных областей суши. Int J Climatol 25 (15): 1965–1978. https://doi.org/10.1002/joc.1276
Int J Climatol 25 (15): 1965–1978. https://doi.org/10.1002/joc.1276
Артикул Google ученый
Холлидей Дж. А., Ритланд К., Эйткен С. Н. (2010) Широко распространенные, экологически значимые генетические маркеры, разработанные на основе ассоциативного картирования климатических признаков у ели ситкинской (Picea sitchensis).Новый Фитол. 188 (2): 501–514. https://doi.org/10.1111/j.1469-8137.2010.03380.x
Артикул PubMed Google ученый
Холлидей Дж. А., Ван Т., Эйткен С. (2012) Прогнозирование адаптивных фенотипов на основе мультилокусных генотипов ели ситкинской ( Picea sitchensis ) с использованием случайного леса. G3: Гены, геномы, генетика 2: 1085–1093
CAS Статья Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>IPCC (2013) Пятый оценочный доклад межправительственной группы экспертов по изменению климата.Cambridge University Press, Нью-Йорк, США
Jermstad KD, Bassoni DL, Jech KS, Ritchie GA, Wheeler NC, Neale DB (2003) Картирование локусов количественных признаков, контролирующих адаптивные признаки у прибрежной пихты Дугласа. III. Количественные характеристики взаимодействия локусов с окружающей средой. Генетика 165 (3): 1489–1506
CAS PubMed PubMed Central Google ученый
Jermstad KD, Bassoni DL, Jech KS, Wheeler NC, Neale DB (2001a) Картирование локусов количественных признаков, контролирующих адаптивные признаки у прибрежной пихты Дугласа.I. Сроки распускания вегетативных почек. Theor Appl Genet 102 (8): 1142–1151. https://doi.org/10.1007/s001220000505
https://doi.org/10.1007/s001220000505
CAS Статья Google ученый
Джермстад К.Д., Бассони Д.Л., Уилер Н.С., Анеконда Т.С., Эйткен С.Н., Адамс В.Т., Нил Д.Б. (2001b) Картирование локусов количественных признаков, контролирующих адаптивные признаки у прибрежной пихты Дугласа. II. Весенне-осенняя холодостойкость. Theor Appl Genet 102 (8): 1152–1158. https: // doi.org / 10.1007 / s001220000506
CAS Статья Google ученый
Джонс MR, Forester BR, Teufel AI, Adams RV, Anstett DN, Goodrich BA, Landguth EL, Joost S, Manel S (2013) Интеграция ландшафтной геномики и пространственно явных подходов для обнаружения локусов под отбором в клинальных популяциях. Evolution 67 (12): 3455–3468. https://doi.org/10.1111/evo.12237
Evolution 67 (12): 3455–3468. https://doi.org/10.1111/evo.12237
CAS Статья PubMed Google ученый
Klopfstein S, Currat M, Excoffier L (2006) Судьба мутаций, перемещающихся по волне расширения диапазона.Mol Biol Evol 23 (3): 482–490. https://doi.org/10.1093/molbev/msj057
CAS Статья PubMed Google ученый
Кремер А., Ронсе О., Робледо-Арнунцио Дж. Дж., Гийом Ф., Борер Дж., Натан Р., Бридл Дж. Р., Гомулкевич Р., Кляйн Е. К., Ритланд К., Купаринен А., Гербер С., Шулер С. (2012). поток генов и адаптация лесных деревьев к быстрым изменениям климата. Ecol Lett 15 (4): 378–392. https://doi.org/10.1111 / j.1461-0248.2012.01746. x
x
Артикул PubMed PubMed Central Google ученый
Крутовский К.В., Clair JBS, Saich R, Hipkins VD, Neale DB (2009) Оценка структуры популяции прибрежной пихты Дугласовой Pseudotsuga menziesii (Mirb.) Franco var. menziesii с использованием аллозимных и микросателлитных маркеров. Древовидные генетические геномы 5 (4): 641–658. https://doi.org/10.1007/s11295-009-0216-y
Артикул Google ученый
Ланглет О. (1971) Двести лет генетики.Таксон 20 (5/6): 653–722. https://doi.org/10.2307/1218596
Артикул Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Ле Корре В., Кремер А. (2003) Генетическая изменчивость по нейтральным маркерам, локусам количественных признаков и признаку в подразделяемой популяции при отборе. Генетика 164 (3): 1205–1219
PubMed PubMed Central Google ученый
Ле Корре В., Кремер А. (2012) Генетическая дифференциация по локусам количественных признаков при локальной адаптации.Мол Экол 21 (7): 1548–1566. https://doi.org/10.1111/j.1365-294X.2012.05479.x
Артикул PubMed Google ученый
Lee BH, Henderson DA, Zhu J-K (2005) Чувствительный к холоду транскриптом Arabidopsis и его регуляция с помощью ICE1. Растительная клетка 17 (11): 3155–3175. https://doi. org/10.1105/tpc.105.035568
org/10.1105/tpc.105.035568
CAS Статья PubMed PubMed Central Google ученый
Lepais O, Bacles CFE (2014) Два лучше, чем один: сочетание ландшафтной геномики и обычных садов для выявления местной адаптации лесных деревьев.Мол Экол 23 (19): 4671–4673. https://doi.org/10.1111/mec.12906
Артикул PubMed Google ученый
Li B, Ning L, Zhang J, Bao M, Zhang W (2015) Транскрипционное профилирование проростков петунии выявляет кандидатов в регуляторы реакции на холодовой стресс. Front Plant Sci 6: 118
PubMed PubMed Central Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Lind BM, Friedline CJ, Wegrzyn J, Maloney PE, Vogler DR, Neale DB, Eckert A (2017) Доступность воды определяет признаки местной адаптации у белой сосны ( Pinus albicaulis Englm.) в мелких пространственных масштабах бассейна озера Тахо, США. Мол Экол 26 (12): 3168–3185. https://doi.org/10.1111/mec.14106
Артикул PubMed Google ученый
Lorenz WW, Sun F, Liang C, Kolychev D, Wang H, Zhao X, Cordonnier-Pratt MM, Pratt LH, Dean JF (2006) Гены, чувствительные к водному стрессу, в корнях дольковой сосны ( Pinus taeda ) идентифицированы анализом библиотек экспрессируемых последовательностей.Tree Physiol 26 (1): 1–16. https://doi.org/10.1093/treephys/26.1.1
Артикул PubMed Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Moura JC, Bonine CA, de Oliveira Fernandes Viana J, Dornelas MC, Mazzafera P (2010) Абиотические и биотические стрессы и изменения содержания и состава лигнина в растениях. Дж. Интегр Биол Растений 52 (4): 360–376. https://doi.org/10.1111/j.1744-7909.2010.00892.x
CAS Статья PubMed Google ученый
Надо С., Мейрманс П.Г., Эйткен С.Н., Ритланд К., Изабель Н. (2016) Проблема отделения признаков местной адаптации от признаков изоляции расстоянием и историей колонизации: случай двух белых сосен.Ecol Evol 6 (24): 8649–8664. https://doi.org/10.1002/ece3.2550
Артикул PubMed PubMed Central Google ученый
Narum SR, Hess JE (2011) Сравнение тестов с выбросами F (ST) для выбранных локусов SNP.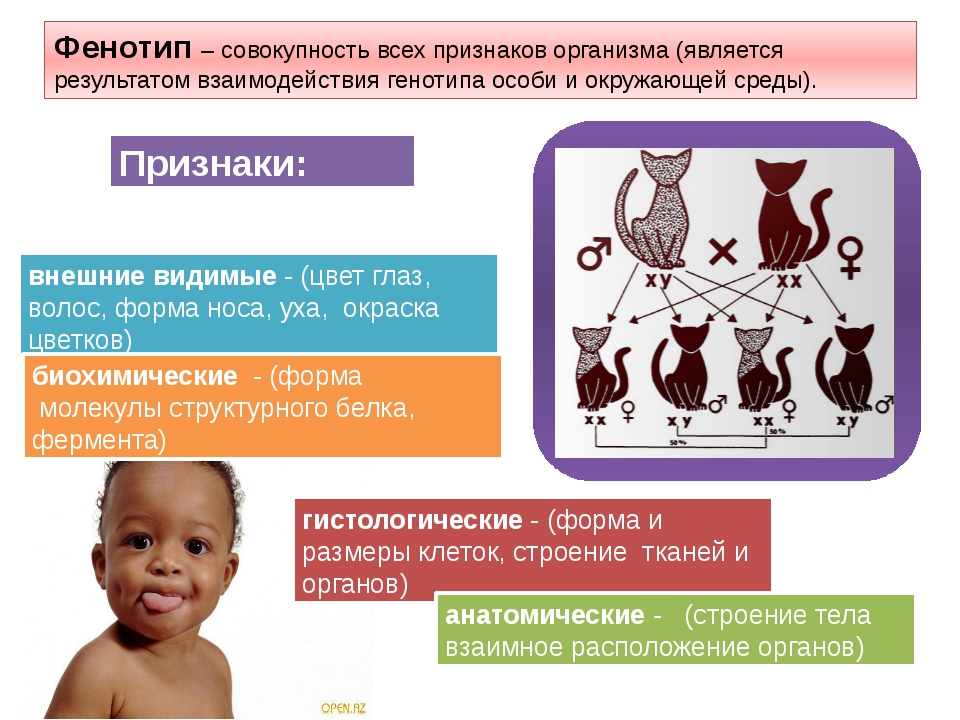 Мол Экол Ресур 11: 184–194. https://doi.org/10.1111/j.1755-0998.2011.02987.x
Мол Экол Ресур 11: 184–194. https://doi.org/10.1111/j.1755-0998.2011.02987.x
Артикул PubMed Google ученый
Neale DB, Kremer A (2011) Геномика лесных деревьев: растущие ресурсы и приложения.Нат Рев Генет 12 (2): 111–122. https://doi.org/10.1038/nrg2931
CAS Статья PubMed Google ученый
Neale DB, Savolainen O (2004) Ассоциативная генетика сложных признаков у хвойных деревьев. Trends Plant Sci 9 (7): 325–330. https://doi.org/10.1016/j.tplants.2004.05.006
CAS Статья PubMed Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>О’Нил Г.А., Эйткен С.Н., Адамс В.Т. (2000) Генетическая селекция на холодоустойчивость саженцев и саженцев прибрежной пихты Дугласа.Can J For Res 30 (11): 1799–1807. https://doi.org/10.1139/x00-114
Артикул Google ученый
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin RP, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner APH 2017 Vegan: экологический пакет сообщества . Версия пакета R 2.4–2
Пардо-Диаз К., Салазар С., Джиггинс С.Д. (2015) На пути к идентификации локусов адаптивной эволюции.Методы Ecol Evol 6 (4): 445–464. https://doi.org/10.1111/2041-210X.12324
Артикул PubMed PubMed Central Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Peakall R, Ruibal M, Lindenmayer DB (2003) Анализ пространственной автокорреляции предлагает новое понимание потока генов у австралийских кустарниковых крыс Rattus fuscipes. Эволюция 57 (5): 1182–1195. https://doi.org/10.1111/j.0014-3820.2003.tb00327.x
Артикул PubMed Google ученый
Peakall R, Smouse PE (2006) Genalex 6: генетический анализ в Excel.Популяционно-генетическое программное обеспечение для обучения и исследований. Mol Ecol Notes 6 (1): 288–295. https://doi.org/10.1111/j.1471-8286.2005.01155.x
Артикул Google ученый
Prunier J, Gerardi S, Laroche J, Beaulieu J, Bousquet J (2012) Параллельная и зависящая от линии молекулярная адаптация к климату у северной черной ели. Мол Экол 21 (17): 4270–4286. https://doi.org/10.1111/j.1365-294X.2012.05691.x
Мол Экол 21 (17): 4270–4286. https://doi.org/10.1111/j.1365-294X.2012.05691.x
CAS Статья PubMed Google ученый
Основная команда R (2013).R: Язык и среда для статистических вычислений. R Фонд статистических вычислений, Вена, Австрия. https://www.R-project.org/.
Rajora OP, Eckert AJ, Zinck JWR (2016) Однолокусные и многолокусные паттерны местной адаптации к климату у восточной белой сосны (Pinus strobus, Pinaceae). PLoS One 11 (7): e0158691. https://doi.org/10.1371/journal.pone.0158691
Артикул PubMed PubMed Central Google ученый
Rehfeldt GE, Jaquish BC, Sáenz-Romero C, Joyce DG, Leites LP, Bradley St Clair J, López-Upton J (2014) Сравнительный генетический ответ на климат у разновидностей Pinus ponderosa и Pseudotsuga menziesii: восстановление лесов. Для Ecol Manag 324: 147–157. https://doi.org/10.1016/j.foreco.2014.02.040
Для Ecol Manag 324: 147–157. https://doi.org/10.1016/j.foreco.2014.02.040
Артикул Google ученый
Институт SAS (2008) Руководство пользователя SAS / STAT, версия 9.2. Кэри, Северная Каролина: SAS Institute Inc.
Schoville SD, Bonin A, François O, Lobreaux S, Melodelima C, Manel S (2012) Адаптивная генетическая изменчивость ландшафта: методы и случаи. Annu Rev Ecol Evol Syst 43 (1): 23–43. https: // doi.org / 10.1146 / annurev-ecolsys-110411-160248
Артикул Google ученый
Сорк В.Л., Эйткен С.Н., Дайер Р.Дж., Эккерт А.Дж., Лежандр П. , Нил Д.Б. (2013) Включение ландшафта в геномику деревьев: подходы к пониманию местной адаптации и реакции населения на изменение климата. Tree Genet Genomes 9 (4): 901–911. https://doi.org/10.1007/s11295-013-0596-x
, Нил Д.Б. (2013) Включение ландшафта в геномику деревьев: подходы к пониманию местной адаптации и реакции населения на изменение климата. Tree Genet Genomes 9 (4): 901–911. https://doi.org/10.1007/s11295-013-0596-x
Артикул Google ученый
St Clair JB (2006) Генетическая изменчивость морозостойкости прибрежной пихты Дугласа в западном Орегоне и Вашингтоне.Can J Bot Rev. Can Bot 84: 1110–1121
Статья Google ученый
St Clair JB, Mandel NL, Vance-Boland KW (2005) Генекология пихты Дугласа в западном Орегоне и Вашингтоне. Энн Бот 96 (7): 1199–1214. https://doi.org/10.1093/aob/mci278
Артикул PubMed PubMed Central Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Стори Дж. Д. (2002) Прямой подход к количеству ложных открытий.J Roy Stat Soc Ser B Stat Methodol 64 (3): 479–498. https://doi.org/10.1111/1467-9868.00346
Артикул Google ученый
Storey JD, Tibshirani R (2003) Статистическая значимость для полногеномных исследований. Proc Natl Acad Sci U S A 100 (16): 9440–9445. https://doi.org/10.1073/pnas.1530509100
CAS Статья PubMed PubMed Central Google ученый
Travis JMJ, Münkemüller T, Burton OJ, Best A, Dytham C, Johst K (2007) Вредные мутации могут достигать высоких плотностей на фронте волны растущей популяции.Mol Biol Evol 24 (10): 2334–2343. https://doi. org/10.1093/molbev/msm167
org/10.1093/molbev/msm167
CAS Статья PubMed Google ученый
Trevaskis, B., D. J. Bagnall, M. H. Ellis, W. J. Peacock и E. S. Dennis. 2003. Гены MADS-бокса контролируют индуцированное яровизацией цветение злаков. Proceedings of the National Academy of Sciences 100: 13099–13104
Vekemans X, Hardy OJ (2004) Новые открытия, полученные в результате мелкомасштабного анализа пространственной генетической структуры популяций растений.Мол Экол 13 (4): 921–935. https://doi.org/10.1046/j.1365-294X.2004.02076.x
CAS Статья PubMed Google ученый
Ван Т. , Хаманн А., Спитлхаус Д., Мердок Т.Н. (2012) ClimateWNA — пространственные климатические данные с высоким разрешением для западной части Северной Америки. J Appl Meteorol Climatol 61: 16–29
, Хаманн А., Спитлхаус Д., Мердок Т.Н. (2012) ClimateWNA — пространственные климатические данные с высоким разрешением для западной части Северной Америки. J Appl Meteorol Climatol 61: 16–29
Статья Google ученый
Wegrzyn JL, Eckert AJ, Choi M, Lee JM, Stanton BJ, Sykes R, Davis MF, Tsai CJ, Neale DB (2010) Ассоциативная генетика признаков, контролирующих биосинтез лигнина и целлюлозы у черного тополя ( Populus trichocarpa , Salicaceae) вторичной ксилемы.Новый Фитол. 188 (2): 515–532. https://doi.org/10.1111/j.1469-8137.2010.03415.x
CAS Статья PubMed Google ученый
Уиллер NC, Джермстад К.Д., Крутовский К., Эйткен С.Н., Хоу Г.Т., Краковски Дж.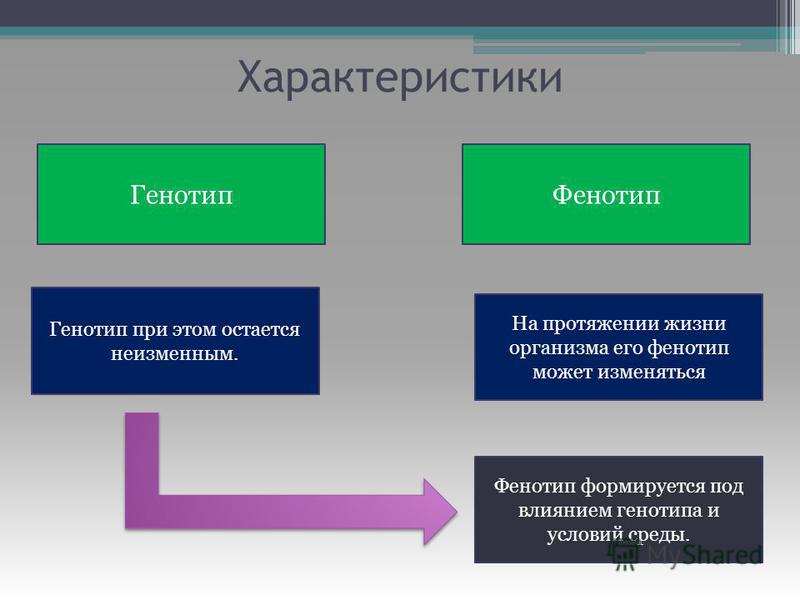 , Нил Д.Б. (2005) Картирование локусов количественных признаков, контролирующих адаптивные признаки у прибрежной пихты Дугласа. IV. Проверка QTL морозостойкости и картирование генов-кандидатов. Мол Порода 15 (2): 145–156. https: // doi.org / 10.1007 / s11032-004-3978-9
, Нил Д.Б. (2005) Картирование локусов количественных признаков, контролирующих адаптивные признаки у прибрежной пихты Дугласа. IV. Проверка QTL морозостойкости и картирование генов-кандидатов. Мол Порода 15 (2): 145–156. https: // doi.org / 10.1007 / s11032-004-3978-9
CAS Статья Google ученый
White TL, WT Adams, DB Neale 2007 Лесная генетика. Кембридж, DOI: https://doi.org/10.1079/9781845932855.0000
Ядав Д.К., Шукла Д., Тутея Н. (2014) Изоляция, in silico характеристика, локализация и анализ экспрессии субъединицы β-субъединицы G-белка риса, чувствительной к абиотическому стрессу (RGB1).Сигнальное поведение растений 9 (5): e28890. https://doi.org/10.4161/psb.28890
Артикул PubMed PubMed Central Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Yeaman S, Hodgins KA, Lotterhos KE, Suren H, Nadeau S, Degner JC, Nurkowski KA, Smets P, Wang T, Gray LK, Liepe KJ, Hamann A, Holliday JA, Whitlock MC, Rieseberg LH, Aitken С.Н. (2016) Конвергентная местная адаптация к климату отдаленных хвойных пород. Science 353 (6306): 1431–1433.https://doi.org/10.1126/science.aaf7812
CAS Статья PubMed Google ученый
Моделирование генотип-фенотипических отношений и болезней человека с помощью сетей генетического взаимодействия
РЕЗЮМЕ
Вероятно, все наследственные признаки, включая восприимчивость к болезням, являются
затронуты взаимодействиями между мутациями в нескольких генах. Мы понимаем
мало, однако, о том, как гены взаимодействуют, чтобы произвести фенотипы, и есть
мало возможностей для обнаружения взаимодействий между генами в популяционных исследованиях людей.Альтернативный подход к пониманию того, как мутации объединяются, чтобы произвести
фенотипов заключается в построении систематических сетей генетического взаимодействия в модели
организмы. Здесь я описываю методы, которые используются для картирования генетических
взаимодействий в дрожжах и C. elegans , и выводы, что эти
сети обеспечивают болезнь человека. Я также обсуждаю механистический
интерпретация сетей генетического взаимодействия, как генетические взаимодействия могут
использоваться для понимания функции генов, а также методы, которые были разработаны для
предсказывать генетические взаимодействия в масштабе всего генома.
Здесь я описываю методы, которые используются для картирования генетических
взаимодействий в дрожжах и C. elegans , и выводы, что эти
сети обеспечивают болезнь человека. Я также обсуждаю механистический
интерпретация сетей генетического взаимодействия, как генетические взаимодействия могут
использоваться для понимания функции генов, а также методы, которые были разработаны для
предсказывать генетические взаимодействия в масштабе всего генома.
Введение
Связь между генотипом организма и его фенотипом
не простое однозначное сопоставление генов и фенотипов. Скорее
фенотипы являются результатом взаимодействия между продуктами многих различных
гены. Сложность этих отношений хорошо иллюстрирует генетика.
болезней у людей: вероятно, все наследственные болезни у людей
генетически сложный, возникший не в результате мутации в одном гене, а в результате
комбинация мутаций в нескольких разных генах
(Бадано и Кацанис, 2002).Например, даже в случае простой «менделевской» болезни кистозная
фиброз, невозможно предсказать клинический фенотип пациента
основанный исключительно на знании точной мутации в «гене кистозного фиброза»,
CFTR. Фактически описано по крайней мере семь различных генов-модификаторов, которые
изменить клинический фенотип этого генетически простого заболевания
(Бадано и Кацанис,
2002).
Фактически описано по крайней мере семь различных генов-модификаторов, которые
изменить клинический фенотип этого генетически простого заболевания
(Бадано и Кацанис,
2002).
Большинство наследственных заболеваний генетически намного сложнее кистозных
фиброз; хотя все большее количество генов было идентифицировано как
мутировал при распространенных патологиях, таких как сердечно-сосудистые заболевания, рак, диабет
и нейродегенеративных заболеваний, эти мутации составляют лишь небольшую часть
доля общей генетической предрасположенности к этим состояниям
(Бадано и Кацанис, 2002).Одна из причин, по которой причинные мутации оказалось так сложно идентифицировать, может заключаться в
проблема синтетических взаимодействий между генами: мутации, имеющие
небольшое влияние на фенотипы болезни может иметь сильные синтетические эффекты
в сочетании (Hartman et al.,
2001). Действительно, в большинстве исследований связей или ассоциаций
недостаточная статистическая мощность для выявления этих взаимодействий между генами
(Бадано и Кацанис, 2002). Следовательно, степень и важность генетических взаимодействий в заболеваниях человека
остается в основном неизвестным.
Следовательно, степень и важность генетических взаимодействий в заболеваниях человека
остается в основном неизвестным.
Альтернативный подход к пониманию того, как гены взаимодействуют, чтобы производить
фенотипы — это выявление генетических взаимодействий между мутациями в модели
организмов (Hartman et al.,
2001). Идея этого подхода состоит в том, чтобы взять простой фенотип.
(обычно самый простой, жизнеспособный) и всесторонне определить, как
комбинации мутаций в генах могут повлиять на этот фенотип. Хотя там
много важных типов усугубления и облегчения генетических взаимодействий
это может происходить между генами (Drees et al.,
2005), на сегодняшний день большая часть работ сосредоточена на синтетических летальных
взаимодействия. Синтетическое (или синергетическое) летальное взаимодействие формально
определяется, когда выживаемость в результате объединения мутаций в двух генах равна
меньше, чем продукт выживаемости в результате каждой мутации
индивидуально (Drees et al.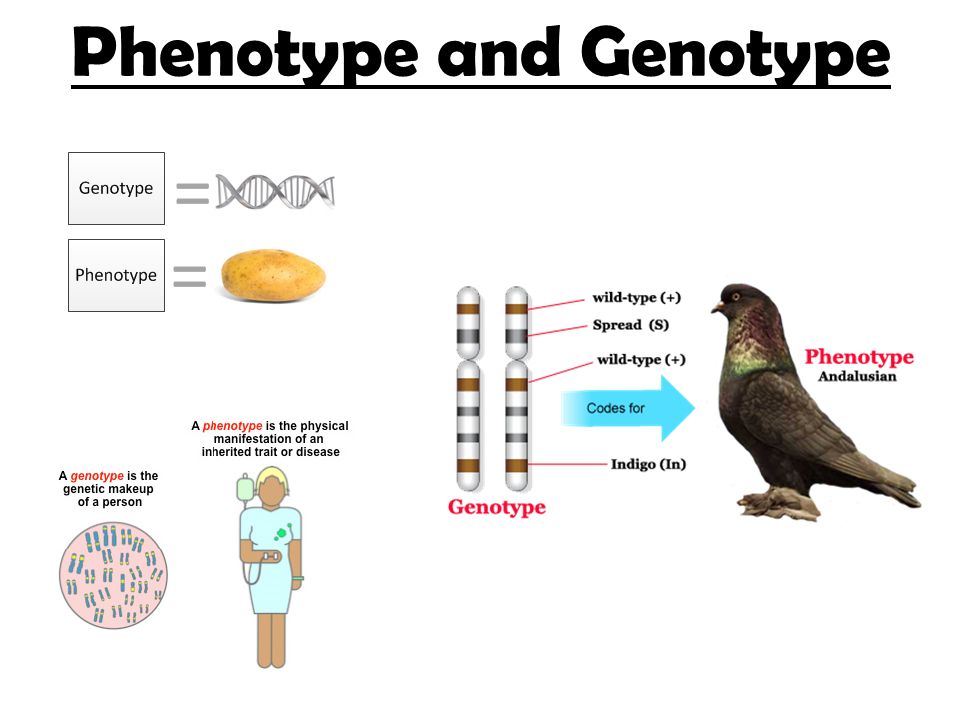 ,
2005). Выявлены наиболее часто синтетические летальные взаимодействия.
экспериментально, когда комбинация мутаций в двух несущественных генах
производит летальный фенотип (рис.
1).
,
2005). Выявлены наиболее часто синтетические летальные взаимодействия.
экспериментально, когда комбинация мутаций в двух несущественных генах
производит летальный фенотип (рис.
1).
Синтетические генетические взаимодействия. Синтетическое смертельное взаимодействие между двумя
гены определяется, когда выживаемость комбинированной мутации меньше, чем
продукт выживания двух одиночных мутаций. В дрожжах генетический
взаимодействия определяются путем объединения мутантных штаммов с использованием систематического скрещивания
протоколы (A), и синтетические летальные или больные фенотипы определены там, где
двойной мутантный штамм демонстрирует фенотип, который не наблюдается ни у одного
мутантный штамм. В г.elegans , генетические взаимодействия определяются
комбинирование генетических мутаций с РНКи для нацеливания на второй ген (B) или с помощью
комбинаторные РНКи для одновременной нацеливания на два гена (C)
(Tischler et al., 2006).
Синтетические отягчающие фенотипы могут быть аналогичным образом определены для других фенотипов. такие как бесплодие или рост (Lehner et al.
al., 2006b) и многие другие возможные комбинации отягчающих или отягчающих обстоятельств.
облегчение взаимодействия также возможно
(Drees et al., 2005).
такие как бесплодие или рост (Lehner et al.
al., 2006b) и многие другие возможные комбинации отягчающих или отягчающих обстоятельств.
облегчение взаимодействия также возможно
(Drees et al., 2005).
Картирование сетей генетического взаимодействия дрожжей
В настоящее время существует наиболее широко отображаемая сеть генетического взаимодействия.
для зародышевых дрожжей Saccharomyces cerevisiae (Ooi et al., 2006;
Пан и др., 2006;
Шульдинер и др., 2005;
Тонг и др., 2001;
Тонг и др., 2004). Два
были разработаны общие стратегии, использующие библиотеку дрожжей
штаммы с делецией генов (Giaever et al.,
2002) для систематического выявления генетических взаимодействий в этом
гаплоидный организм. В первом подходе конструируются двойные мутанты и
анализируется на жизнеспособность параллельно путем скрещивания штамма дрожжей, несущего запрос
мутация в полную библиотеку жизнеспособных делеционных штаммов в массиве
формат [анализ `синтетического генетического массива ‘, SGA
(Тонг и др. , 2001)]. В
напротив, во втором подходе конструируются и анализируются двойные мутанты как
единый пул, и определяются относительные скорости роста каждого штамма
использование микрочипов ДНК, которые могут определять присутствие каждого штамма в пуле
[`синтетический летальный анализ с помощью микроматрицы ‘, SLAM
(Ooi et al., 2003)]. я буду
сначала обрисуйте эти два метода, а затем обсудите генетическое взаимодействие
сети, которые были созданы с их использованием.
, 2001)]. В
напротив, во втором подходе конструируются и анализируются двойные мутанты как
единый пул, и определяются относительные скорости роста каждого штамма
использование микрочипов ДНК, которые могут определять присутствие каждого штамма в пуле
[`синтетический летальный анализ с помощью микроматрицы ‘, SLAM
(Ooi et al., 2003)]. я буду
сначала обрисуйте эти два метода, а затем обсудите генетическое взаимодействие
сети, которые были созданы с их использованием.
SGA
В подходе SGA (Tong et al.,
2001), гаплоидный штамм дрожжей, несущий «запросную» мутацию в
представляющий интерес ген связывается с библиотекой дрожжевых штаммов с делецией в массиве
форматирование с использованием инструментов копирования и робототехники. Затем диплоидные дрожжи
спорулированное и двойное мутантное гаплоидное потомство отбирают с использованием умного
сконструированная репортерная конструкция (репортер SGA, can1 Δ :: MFA1pr-HIS3 , который присутствует в MATalpha штамм запроса , но в отсутствие гистидина позволяет
рост гаплоидов типа спаривания MATa , т. е.е. только двойной мутант
потомство). Синтетические больные или нежизнеспособные двойные мутанты идентифицируются слабо
растущие или отсутствующие двойные мутантные колонии, и их идентичность определяется
их положение в массиве. Возможные синтетические летальные или болезненные взаимодействия
затем индивидуально подтверждаются тетрадным или случайным анализом спор
(Тонг и др., 2001).
е.е. только двойной мутант
потомство). Синтетические больные или нежизнеспособные двойные мутанты идентифицируются слабо
растущие или отсутствующие двойные мутантные колонии, и их идентичность определяется
их положение в массиве. Возможные синтетические летальные или болезненные взаимодействия
затем индивидуально подтверждаются тетрадным или случайным анализом спор
(Тонг и др., 2001).
SLAM
В подходе SLAM мутация запроса вводится в пул
штаммы с гаплоидной делецией путем прямой интегративной трансформации
(Ooi et al., 2003). Двойной
затем мутанты выращивают в одном сосуде для конкуренции, а нерастущие или
медленнорастущие двойные мутантные штаммы идентифицируют с помощью микрочипов. Этот
подход возможен благодаря двум последовательностям ДНК «штрих-кода», которые однозначно
идентифицировать каждый штамм делеции. Эти штрих-коды позволяют удалять штаммы, которые
присутствуют в пуле, чтобы быть индивидуально идентифицированными путем гибридизации
геномной ДНК на микроматрицу, содержащую последовательности, комплементарные каждой из
штрих-коды. В отличие от качественного подхода SGA, в процедуре SLAM
определение синтетического взаимодействия зависит от количественного ограничения
по интенсивности гибридизации для выявления медленно растущих или отсутствующих штаммов.А
модификация процедуры SLAM использует гетерозиготные диплоидные делеционные штаммы
в качестве отправной точки [SLAM на основе диплоидов или dSLAM
(Pan et al., 2004)].
Сохранение делеционных штаммов в качестве гетерозиготных диплоидов защищает их от
отбор компенсаторных или обратных мутаций, которые преодолевают приспособленность
дефекты, что снижает количество ложных отрицательных результатов подхода. В результате
Подход dSLAM, вероятно, имеет более низкий уровень ложноотрицательных результатов, чем SGA.
(Тонг и др., 2004). Один
Недостатком подхода dSLAM является то, что некоторые теги штрих-кода имеют низкую
интенсивности сигнала гибридизации (например, из-за мутаций в теге
последовательностей), в результате чего достоверной информации о некоторых
гены (Eason et al.
В отличие от качественного подхода SGA, в процедуре SLAM
определение синтетического взаимодействия зависит от количественного ограничения
по интенсивности гибридизации для выявления медленно растущих или отсутствующих штаммов.А
модификация процедуры SLAM использует гетерозиготные диплоидные делеционные штаммы
в качестве отправной точки [SLAM на основе диплоидов или dSLAM
(Pan et al., 2004)].
Сохранение делеционных штаммов в качестве гетерозиготных диплоидов защищает их от
отбор компенсаторных или обратных мутаций, которые преодолевают приспособленность
дефекты, что снижает количество ложных отрицательных результатов подхода. В результате
Подход dSLAM, вероятно, имеет более низкий уровень ложноотрицательных результатов, чем SGA.
(Тонг и др., 2004). Один
Недостатком подхода dSLAM является то, что некоторые теги штрих-кода имеют низкую
интенсивности сигнала гибридизации (например, из-за мутаций в теге
последовательностей), в результате чего достоверной информации о некоторых
гены (Eason et al. , 2004).
Однако это ограничение было устранено за счет изменения конструкции используемых микрочипов.
для обнаружения штрих-кодов (Pierce et al.,
2006; Юань и др.,
2005).
, 2004).
Однако это ограничение было устранено за счет изменения конструкции используемых микрочипов.
для обнаружения штрих-кодов (Pierce et al.,
2006; Юань и др.,
2005).
Оба подхода — SGA и dSLAM — использовались для построения обширных
сети генетического взаимодействия у дрожжей. Используя подход SGA, Тонг и др.
проверили 132 запросных штамма (несущих мутации в генах с различными функциями
в полярности клеток, биосинтезе клеточной стенки, сегрегации хромосом и ДНК
синтез и ремонт) против полной библиотеки ∼4700 жизнеспособных гаплоидов
делеционных штаммов, и идентифицировали в общей сложности 2012 синтетических летальных и 2113
синтетические взаимодействия с больными с участием ~ 1000 генов
(Тонг и др., 2004). Обе
делеции несущественных генов и точечные мутации в основных генах были
использовались в качестве генов запросов, а синтетические летальные взаимодействия были обнаружены в 80%
штаммов запросов, в среднем 34 взаимодействия на ген запроса (и диапазон
1–146 взаимодействий на ген). Используя подход dSLAM, Pan et al.
проверили 74 запросных штамма, которые, как известно, участвуют в репликации и репарации ДНК
против той же самой делеционной библиотеки и идентифицировали в общей сложности 4956 синтетических
дефекты пригодности или летальности с участием 875 генов
(Пан и др., 2006). Более 91%
из этих взаимодействий были совершенно новыми
(Пан и др., 2006).
Используя подход dSLAM, Pan et al.
проверили 74 запросных штамма, которые, как известно, участвуют в репликации и репарации ДНК
против той же самой делеционной библиотеки и идентифицировали в общей сложности 4956 синтетических
дефекты пригодности или летальности с участием 875 генов
(Пан и др., 2006). Более 91%
из этих взаимодействий были совершенно новыми
(Пан и др., 2006).
Картирование генетических взаимодействий основных генов
И Тонг, и др. и Pan et al. исследует проверенные штаммы запросов на
~ 4700 жизнеспособных штаммов с делецией дрожжей, поэтому взаимодействие с ~
Не удалось обнаружить 1000 основных генов в геноме дрожжей. Два
были разработаны подходы для выявления генетических взаимодействий с основными
гены (Davierwala et al., 2005;
Schuldiner et al., 2005). В
первый подход, Davierwala et al. построил библиотеку штаммов дрожжей
которые несут аллели замены промотора. Эти аллели позволяют выражать
каждый ген отключается добавлением доксициклина с небольшой молекулой
СМИ (система «тет-офф»).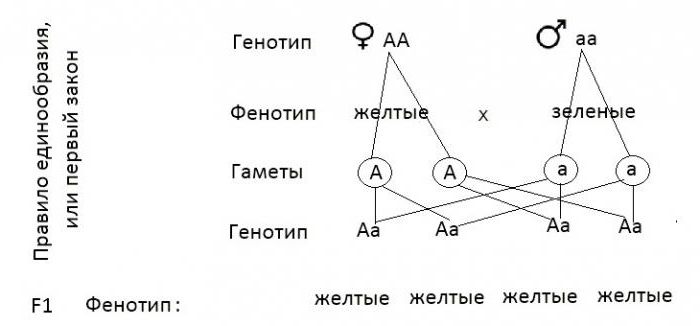 Добавление промежуточных уровней
поэтому доксициклин можно использовать для снижения экспрессии каждого существенного
ген, таким образом продуцируя гипоморфные (снижение функции) аллели каждого гена.
Авторы создали библиотеку, состоящую из аллелей замены промоторов для
575 основных генов (что составляет примерно половину от общего числа основных
генов) и проверил его по 30 запросам, которые были либо условными
аллели основных генов или делеции несущественных генов, идентифицирующие
всего 567 взаимодействий.Интересно, что среднее количество взаимодействий
обнаружено для каждого существенного гена примерно в шесть раз больше, чем для несущественного
гены (Davierwala et al.,
2005).
Добавление промежуточных уровней
поэтому доксициклин можно использовать для снижения экспрессии каждого существенного
ген, таким образом продуцируя гипоморфные (снижение функции) аллели каждого гена.
Авторы создали библиотеку, состоящую из аллелей замены промоторов для
575 основных генов (что составляет примерно половину от общего числа основных
генов) и проверил его по 30 запросам, которые были либо условными
аллели основных генов или делеции несущественных генов, идентифицирующие
всего 567 взаимодействий.Интересно, что среднее количество взаимодействий
обнаружено для каждого существенного гена примерно в шесть раз больше, чем для несущественного
гены (Davierwala et al.,
2005).
Вторая стратегия, которая использовалась для определения взаимодействий для
существенные гены должны генерировать гипоморфные аллели путем замены
3’UTR каждого гена с кассетой устойчивости к антибиотикам [`уменьшено
численность по пертурбации мРНК ‘, DAmP
(Schuldiner et al., 2005)]. В
При таком подходе кассета устойчивости к антибиотикам служит для дестабилизации
экспрессия мРНК, что снижает экспрессию каждого важного гена. В
Подход DAmP был использован для выявления генетических взаимодействий между генами, которые
функция в раннем секреторном пути
(Schuldiner et al., 2005) (см.
ниже).
В
Подход DAmP был использован для выявления генетических взаимодействий между генами, которые
функция в раннем секреторном пути
(Schuldiner et al., 2005) (см.
ниже).
Теоретически, все эти методы выявления взаимодействий между парами
генов также можно использовать для выявления взаимодействий более высокого порядка между большим количеством
чем два гена. Например, Тонг и др. также использовал SGA для отображения двух разных
штаммы с двойными мутантами для взаимодействия с третьим геном для идентификации тригенных
взаимодействия (Тонг и др.,
2004 г.). Всего авторы идентифицировали 171 и 156 взаимодействий.
на этих экранах, хотя только 4 и 29 взаимодействий можно отнести
к эффекту тройного мутанта (остальные также наблюдались в одном из трех
только возможные комбинации двойных мутантов). Однако, поскольку есть ~
Возможных триплетов генов в 2000 раз больше, чем пар генов в S.
cerevisiae , общее количество тригенных синтетических летальных
взаимодействий может быть больше, чем количество дигенных взаимодействий.
Количественные скрины генетического взаимодействия
Подходы SGA и dSLAM использовались для выявления синтетических летальных
и больные фенотипы в геномном масштабе. Однако есть много других классов
взаимодействий, которые могут происходить между генами, и подходы также были
разработан, чтобы начать систематическое выявление этих взаимодействий
(Коллинз и др., 2006;
Drees et al., 2005;
Хартман и Типпери, 2004;
Schuldiner et al., 2005). За
например, Schuldiner et al.использовали цифровые изображения для количественной оценки роста
колонии дрожжей, позволяющие измерить как отягчающее, так и облегчающее
взаимодействия между 424 дрожжевыми генами, которые функционируют в ранней секреторной
путь (Schuldiner et al.,
2005). Вместо того, чтобы разделять наблюдаемые взаимодействия на
различные типы генетического взаимодействия, они использовали непрерывный счет для
описать силу взаимодействия и сгруппировать гены в соответствии с
их профили взаимодействия. Авторы продемонстрировали, что использование количественных
измерения силы взаимодействия помогли идентифицировать модули генов, которые
разделяют точные молекулярные функции
(Шульдинер и др., 2005).
Авторы продемонстрировали, что использование количественных
измерения силы взаимодействия помогли идентифицировать модули генов, которые
разделяют точные молекулярные функции
(Шульдинер и др., 2005).
Картирование сетей генетического взаимодействия в
C. elegansЭти исследования на S. cerevisiae предоставили беспрецедентное понимание в масштабе и свойствах сетей генетического взаимодействия. Тем не мение, S. cerevisiae — одноклеточные дрожжи, не содержащие многих из гены и пути, присутствующие в многоклеточных организмах. В в частности, многие из сигнальных путей, участвующих в заболеваниях человека, таких как поскольку рак не закодирован в геноме дрожжей.Поэтому понять, как гены будут взаимодействовать у людей, важно, чтобы мы также систематически выявлять генетические взаимодействия у многоклеточных животных.
Систематическое определение генетических взаимодействий путем скрещивания мутантных штаммов
не является логистически практичным в многоклеточных организмах — всеобъемлющий
коллекции делеционных штаммов недоступны, и диплоидия этих
организмов требует громоздких экранов для спаривания и отбора из нескольких поколений
использоваться. Альтернативный подход к идентификации генетических взаимодействий в
Metazoans — использовать РНК-интерференцию (РНКи) для подавления экспрессии генов
(Baugh et al., 2005;
Холуэй и др., 2005;
Ленер и др., 2006а;
Ленер и др., 2006b;
Ленер и др., 2006c;
Сузуки и Хан, 2006 г .;
Тишлер и др., 2006;
van Haaften et al., 2004).
Здесь либо генетический мутант сочетается с РНКи против второго гена.
(Рис. 1B), или можно использовать РНКи
подавлять экспрессию двух генов одновременно
(Рис. 1C)
(Tischler et al., 2006).Один
Преимущество использования РНКи по сравнению со штаммами с делецией состоит в том, что, поскольку РНКи
обычно производит «нокдаун», а не «нокаут», это также
можно идентифицировать взаимодействия для основных генов.
Альтернативный подход к идентификации генетических взаимодействий в
Metazoans — использовать РНК-интерференцию (РНКи) для подавления экспрессии генов
(Baugh et al., 2005;
Холуэй и др., 2005;
Ленер и др., 2006а;
Ленер и др., 2006b;
Ленер и др., 2006c;
Сузуки и Хан, 2006 г .;
Тишлер и др., 2006;
van Haaften et al., 2004).
Здесь либо генетический мутант сочетается с РНКи против второго гена.
(Рис. 1B), или можно использовать РНКи
подавлять экспрессию двух генов одновременно
(Рис. 1C)
(Tischler et al., 2006).Один
Преимущество использования РНКи по сравнению со штаммами с делецией состоит в том, что, поскольку РНКи
обычно производит «нокдаун», а не «нокаут», это также
можно идентифицировать взаимодействия для основных генов.
C. elegans — уникальная модель животного, у которой генетические взаимодействия
может быть идентифицирован in vivo в контексте развивающегося организма;
экспрессия любого гена может быть системно подавлена с помощью длинных дцРНК
доставляется путем бактериального вскармливания (Timmons
and Fire, 1998).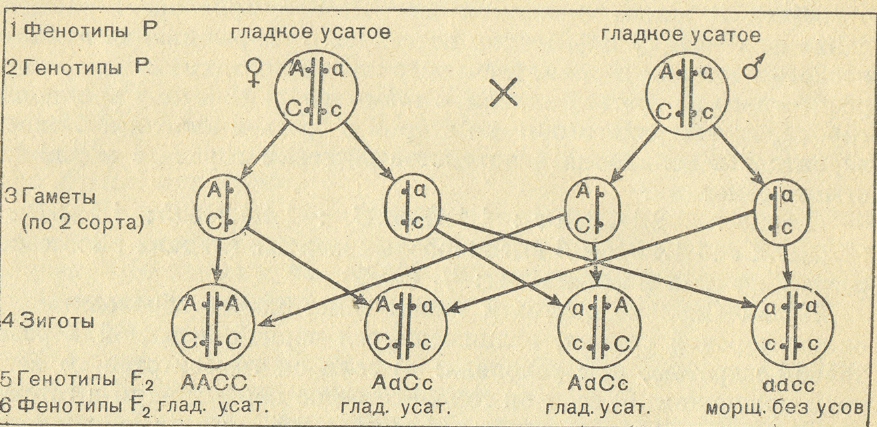 Хотя должно быть возможно идентифицировать
генетические взаимодействия между генами с использованием РНКи в культуре клеток млекопитающих или
клетки мух, C. elegans в настоящее время является единственным модельным организмом, в котором
этот подход может быть использован in vivo в широком масштабе. В г.
elegans , скрининг РНКи можно проводить в жидкой культуре в 96-луночных
пластины (Lehner et al., 2006c;
van Haaften et al., 2004)
использование библиотеки бактериального кормления
(Камат и др., 2003). Этот
позволяет проводить скрининг РНКи с достаточной пропускной способностью, чтобы иметь возможность
проверить десятки тысяч пар генов на их способность к генетическому взаимодействию in vivo .
Хотя должно быть возможно идентифицировать
генетические взаимодействия между генами с использованием РНКи в культуре клеток млекопитающих или
клетки мух, C. elegans в настоящее время является единственным модельным организмом, в котором
этот подход может быть использован in vivo в широком масштабе. В г.
elegans , скрининг РНКи можно проводить в жидкой культуре в 96-луночных
пластины (Lehner et al., 2006c;
van Haaften et al., 2004)
использование библиотеки бактериального кормления
(Камат и др., 2003). Этот
позволяет проводить скрининг РНКи с достаточной пропускной способностью, чтобы иметь возможность
проверить десятки тысяч пар генов на их способность к генетическому взаимодействию in vivo .
Используя высокопроизводительные экраны РНКи в C. elegans , мы недавно
построил первую сеть систематического генетического взаимодействия для любого животного
(Lehner et al., 2006b). Мы
сосредоточено на генах, которые функционируют в сигнальных путях, и протестировано> 65 000
пары генов для их способности взаимодействовать in vivo с использованием обоих
генетические мутантные запросные штаммы и комбинаторные РНКи.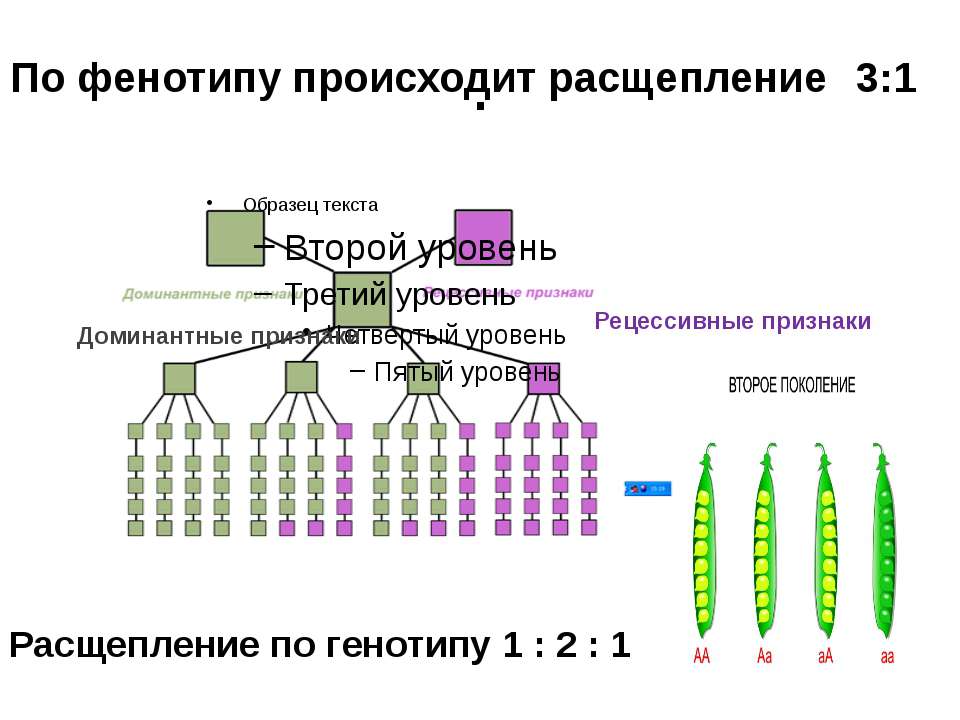 Всего мы определили
351 пара генов, которые при комбинированной инактивации производили синтетический
нежизнеспособный фенотип.
Всего мы определили
351 пара генов, которые при комбинированной инактивации производили синтетический
нежизнеспособный фенотип.
Механистическая интерпретация сетей генетического взаимодействия
Синтетические летальные взаимодействия из мелкомасштабных исследований обычно
интерпретируется как предоставление подтверждающих доказательств того, что два генных продукта действуют
либо одним и тем же биохимическим путем, либо двумя параллельными путями, которые могут
функционально компенсируют друг друга
(Хартман и др., 2001). В
в их оригинальной статье Tong et al. отметил, что хотя ∼27% генетических
взаимодействия, которые они определили, связывают гены со сходной онтологией генов (GO)
аннотации, только ∼1% синтетических летальных взаимодействий происходит между генами
чьи продукты находятся в одном и том же белковом комплексе
(Тонг и др., 2004). Скорее,
они продемонстрировали, что гены, кодирующие продукты, функционируют в том же
белковый комплекс или путь часто имеют схожие профили генетических взаимодействий
(т.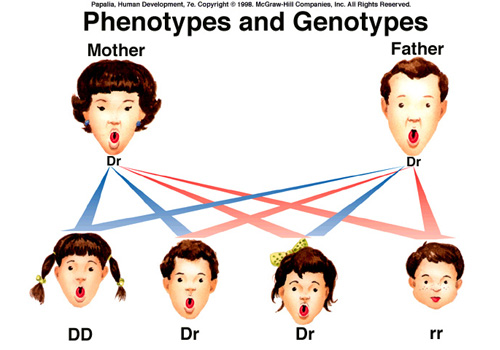 е. гены одного пути, как правило, взаимодействуют с одними и теми же генами, скорее
чем друг с другом). Действительно, чем больше генетических взаимодействий имеют два гена,
тем более вероятно, что эти два генных продукта будут физически взаимодействовать (см. ниже)
(Тонг и др., 2004).
е. гены одного пути, как правило, взаимодействуют с одними и теми же генами, скорее
чем друг с другом). Действительно, чем больше генетических взаимодействий имеют два гена,
тем более вероятно, что эти два генных продукта будут физически взаимодействовать (см. ниже)
(Тонг и др., 2004).
Путем объединения данных генетического взаимодействия с комплексными
данные белок-белок, белок-ДНК и метаболические сети, Kelley
и Идекер систематически сравнивали возможности моделей «внутри пути»
(также называемые «внутрипутными» или «серийными» моделями,
Инжир.2А) или «промежуточный путь»
(«междурядные» или «параллельные») модели
(Рис. 2B), чтобы объяснить
систематически собираемые данные о генетическом взаимодействии
(Келли и Идекер, 2005).
Используя вероятностную модель, они обнаружили, что модели между путями могут
объясняют в три с половиной раза больше взаимодействий, чем внутрипутьевые модели.
Однако они не смогли дать механистическую интерпретацию для ~
60% наблюдаемых генетических взаимодействий у дрожжей. Действительно обширный
генетические взаимодействия, выявленные в исследованиях SGA и dSLAM, предполагают
что между, казалось бы,
несвязанные клеточные пути.Например, Pan et al. наблюдается обширный
функциональная компенсация между потерей генов пути ответа на повреждение ДНК и
гены, участвующие в транскрипции мРНК, процессинге мРНК и целостности Гольджи
(Пан и др., 2006). Следовательно
механистическая интерпретация генетических взаимодействий остается важной
площадь для будущих работ.
Действительно обширный
генетические взаимодействия, выявленные в исследованиях SGA и dSLAM, предполагают
что между, казалось бы,
несвязанные клеточные пути.Например, Pan et al. наблюдается обширный
функциональная компенсация между потерей генов пути ответа на повреждение ДНК и
гены, участвующие в транскрипции мРНК, процессинге мРНК и целостности Гольджи
(Пан и др., 2006). Следовательно
механистическая интерпретация генетических взаимодействий остается важной
площадь для будущих работ.
Модели генетических взаимодействий внутри и между путями. Синтетический
летальные взаимодействия (пунктирные линии) могут происходить как между двумя компонентами
одиночный биохимический путь (A) или между компонентами двух параллельных путей
которые могут функционально компенсировать друг друга (B).Келли и Идекер нашли
что сочетание моделей внутри и между путями может объяснить
40% синтетических смертельных или болезненных взаимодействий в дрожжах с промежуточными путями
модели преобладают. Примеры взаимодействий внутри пути включают:
взаимодействия между компонентами сплайсосомы и взаимодействия между
компоненты комплекса казеинкиназы 2. Межпутевые взаимодействия
включают обширные взаимодействия между компонентами комплекса Dynactin и
компоненты префолдинового комплекса
(Келли и Идекер, 2005).За
взаимодействия между частичными мутациями потери функции, однако,
внутрипутьевые модели могут преобладать. Гены / белки показаны в виде узлов,
взаимодействия белков как сплошные края, а генетические взаимодействия как нарушенные
края.
Примеры взаимодействий внутри пути включают:
взаимодействия между компонентами сплайсосомы и взаимодействия между
компоненты комплекса казеинкиназы 2. Межпутевые взаимодействия
включают обширные взаимодействия между компонентами комплекса Dynactin и
компоненты префолдинового комплекса
(Келли и Идекер, 2005).За
взаимодействия между частичными мутациями потери функции, однако,
внутрипутьевые модели могут преобладать. Гены / белки показаны в виде узлов,
взаимодействия белков как сплошные края, а генетические взаимодействия как нарушенные
края.
В отличие от дрожжей, у C. elegans генетический
сеть взаимодействия (Lehner et al.,
2006b), внутрипутьевые взаимодействия, по-видимому, объясняют больше
взаимодействия, чем взаимодействия между путями: сосредоточение внимания на известных компонентах
сигнальных путей, в два раза больше взаимодействий между компонентами
того же пути, чем между компонентами отдельных путей (B.Л.,
неопубликованное наблюдение). Объяснение этому, вероятно, кроется в
различие в используемых экспериментальных подходах: в дрожжах анализируемые взаимодействия
в основном находятся между нулевыми аллелями, тогда как в C. elegans большинство
взаимодействия между гипоморфным аллелем и нокдауном РНКи второго
ген. Это имеет интуитивный смысл: тогда как фенотипические последствия
нулевая мутация в гене линейного пути не может быть дополнительно усилена
вторая мутация в этом пути, две частичные мутации потери функции в
один путь может быть объединен для полного подавления этого пути
(Рис.2). Поэтому с нулевым
аллели, многие взаимодействия, вероятно, представляют собой взаимодействия между
полная инактивация двух несущественных путей, которые могут функционально
компенсируют друг друга, тогда как при гипоморфных аллелях многие из
взаимодействия могут представлять собой взаимодействия между частичной инактивацией
два гена, которые действуют одним и тем же основным путем.
elegans большинство
взаимодействия между гипоморфным аллелем и нокдауном РНКи второго
ген. Это имеет интуитивный смысл: тогда как фенотипические последствия
нулевая мутация в гене линейного пути не может быть дополнительно усилена
вторая мутация в этом пути, две частичные мутации потери функции в
один путь может быть объединен для полного подавления этого пути
(Рис.2). Поэтому с нулевым
аллели, многие взаимодействия, вероятно, представляют собой взаимодействия между
полная инактивация двух несущественных путей, которые могут функционально
компенсируют друг друга, тогда как при гипоморфных аллелях многие из
взаимодействия могут представлять собой взаимодействия между частичной инактивацией
два гена, которые действуют одним и тем же основным путем.
Другой простой класс синтетического летального взаимодействия наблюдается между двумя
дублированные гены, кодирующие гомологичные белки.Хотя многие дублировали
гены явно кодируют избыточные функции, которые поддерживаются
значительные эволюционные периоды
(Tischler et al. , 2006),
тщательный анализ фенотипов мутантных одиночных генов
(Вагнер, 2005) и SGA
синтетические летальные данные (Tong et al.,
2004) предполагает, что дупликации генов объясняют только очень небольшие
меньшинство синтетических летальных взаимодействий [<2% взаимодействий
идентифицированные SGA кодируют гомологичные белки
(Тонг и др., 2004)].
, 2006),
тщательный анализ фенотипов мутантных одиночных генов
(Вагнер, 2005) и SGA
синтетические летальные данные (Tong et al.,
2004) предполагает, что дупликации генов объясняют только очень небольшие
меньшинство синтетических летальных взаимодействий [<2% взаимодействий
идентифицированные SGA кодируют гомологичные белки
(Тонг и др., 2004)].
Использование генетических взаимодействий для понимания функции генов
Тенденция генов, действующих как внутри, так и между путями, к
генетически взаимодействовать означает, что генетические взаимодействия могут использоваться для прогнозирования
функции генов.Как впервые отметили Тонг и др., Компоненты одного и того же пути
имеют одинаковые синтетические летальные партнеры
(Тонг и др., 2004;
Ye et al., 2005). Следовательно
количество общих партнеров генетического взаимодействия для двух генов может быть использовано для
ранжируйте вероятность физического взаимодействия двух продуктов гена и
разделяя биологическую функцию.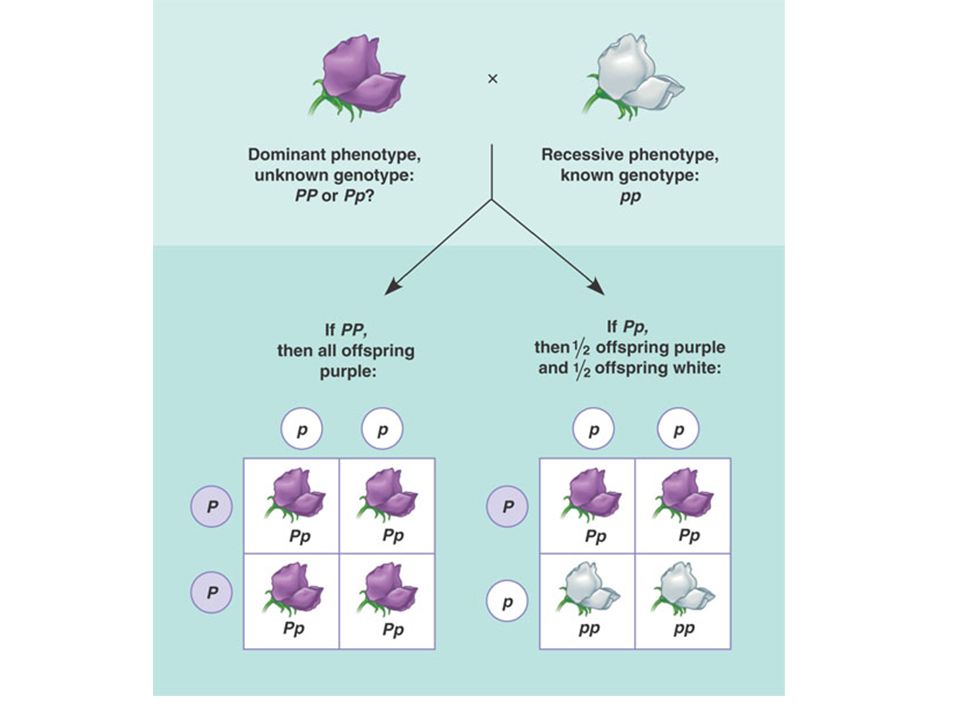 Сходство профилей взаимодействия
два гена можно математически определить по их «конгруэнтности»
(Ye et al., 2005). Конгруэнтность
можно определить как отрицательный логарифм гипергеометрического значения P количество общих генетических взаимодействий соседей двух генов
(Ye et al., 2005), который
учитывает как количество взаимодействий каждого гена, так и общее
размер сети взаимодействия. Используя данные о генетическом взаимодействии дрожжей,
конгруэнтность лучше подходит для прогнозирования физических взаимодействий и общих
функции, чем прямое генетическое взаимодействие или подсчет количества
партнеры по взаимодействию разделяются двумя генами
(Ye et al., 2005). Pan et al.
смогли использовать сравнение, чтобы определить 16 функционально однородных модулей
Гены ответа на повреждение ДНК (Pan et al.,
2006) и смогли использовать профили генетического взаимодействия
новые гены для прогнозирования их функций в ответе на повреждение ДНК
(Пан и др., 2006).
Сходство профилей взаимодействия
два гена можно математически определить по их «конгруэнтности»
(Ye et al., 2005). Конгруэнтность
можно определить как отрицательный логарифм гипергеометрического значения P количество общих генетических взаимодействий соседей двух генов
(Ye et al., 2005), который
учитывает как количество взаимодействий каждого гена, так и общее
размер сети взаимодействия. Используя данные о генетическом взаимодействии дрожжей,
конгруэнтность лучше подходит для прогнозирования физических взаимодействий и общих
функции, чем прямое генетическое взаимодействие или подсчет количества
партнеры по взаимодействию разделяются двумя генами
(Ye et al., 2005). Pan et al.
смогли использовать сравнение, чтобы определить 16 функционально однородных модулей
Гены ответа на повреждение ДНК (Pan et al.,
2006) и смогли использовать профили генетического взаимодействия
новые гены для прогнозирования их функций в ответе на повреждение ДНК
(Пан и др., 2006).
В сети генетического взаимодействия C. elegans , потому что больше
взаимодействия происходят внутри пути, чем между путями, знание
партнеры прямого взаимодействия гена могут использоваться для прогнозирования его функции. За
Например, мы систематически проверяли, все ли обнаруженные гены
взаимодействовать с двумя или более известными компонентами пути EGF / Ras / MAPK действовали
как общие модуляторы этого пути. Для 9/16 таких проверенных случаев мы обнаружили
что гены действительно могут модулировать передачу сигналов EGF в точном процессе развития
установка, предполагая, что они действительно действуют как общие модуляторы этого
путь (Lehner et al.,
2006b).
elegans , потому что больше
взаимодействия происходят внутри пути, чем между путями, знание
партнеры прямого взаимодействия гена могут использоваться для прогнозирования его функции. За
Например, мы систематически проверяли, все ли обнаруженные гены
взаимодействовать с двумя или более известными компонентами пути EGF / Ras / MAPK действовали
как общие модуляторы этого пути. Для 9/16 таких проверенных случаев мы обнаружили
что гены действительно могут модулировать передачу сигналов EGF в точном процессе развития
установка, предполагая, что они действительно действуют как общие модуляторы этого
путь (Lehner et al.,
2006b).
Один из способов определения точности предсказания функции генов на основе генетических
сети взаимодействия, вероятно, можно улучшить, если выявить отягчающие
и облегчение взаимодействий, и для количественной оценки силы взаимодействий.
Действительно, Schuldiner et al. обнаружили, что включая количественные данные о взаимодействии
помогли определить модули генов, которые разделяют точные молекулярные функции в
ранний секреторный путь (Schuldiner et al. др., 2005).
др., 2005).
Прогнозирование генетических взаимодействий
В идеале мы хотели бы построить всеобъемлющее генетическое взаимодействие
сеть для людей.Затем эта сеть будет служить основой для непосредственного
прогнозирование генов-модификаторов при наследственных и соматических заболеваниях, а также
предоставление ресурса для прогнозирования функции генов человека в геномном масштабе.
Однако выявление генетических взаимодействий в многоклеточных организмах
чем C. elegans чрезвычайно трудоемок. Хотя можно
использовать РНКи для выявления генетических взаимодействий в культивируемых клетках млекопитающих или мух
(Wheeler et al., 2004),
неэффективность и стоимость РНКи в клетках млекопитающих
(Pei and Tuschl, 2006) и
преобладание нецелевых эффектов в обеих системах
(Kulkarni et al., 2006;
Ленер и др., 2004;
Ma et al., 2006), до настоящего времени
ограничили применение этого подхода. К тому же не совсем понятно
как синтетические летальные взаимодействия, выявленные в отдельных клетках, будут связаны с
взаимодействия в целых организмах.
Альтернатива идентификации генетических взаимодействий с использованием крупномасштабных
экспериментальные подходы заключается в использовании вычислительных методов для прогнозирования генетических
взаимодействия между генами. Это аналогично сетям взаимодействия белков,
где сейчас находятся методы компьютерного прогнозирования белковых взаимодействий
разработаны до такой степени, что они по крайней мере так же точны, как и большинство
высокопроизводительные экспериментальные протоколы или взаимодействия, основанные на
литература (Jansen et al.,
2003; Ли и др.,
2004; Троянская и др.,
2003 г.). Здесь я обсуждаю три подхода, которые использовались для
прогнозирование генетических взаимодействий: (i) с использованием существующих генетических взаимодействий и
структура локальной сети для прогнозирования новых взаимодействий, (ii) использование
интеграция других наборов геномных данных для прогнозирования генетических взаимодействий и
(iii) использование взаимодействий от одного вида для прогнозирования взаимодействий за второй
разновидность.
Прогнозирование генетических взаимодействий с использованием сетевой структуры
Одним из свойств сети генетического взаимодействия дрожжей является то, что два гена которые имеют генетическое взаимодействие с общим партнером, вероятно, будут взаимодействовать друг с другом (Тонг и др., 2004 г.). Тонг и др. впервые использовали эту особенность «маленького мира» сети генетического взаимодействия для прогнозирования дальнейших взаимодействий и обнаружили, что в ~ В 20% случаев соседи гена запроса также могут взаимодействовать с каждым другие (по сравнению с <1% случайных пар генов).
Прогнозирование генетических взаимодействий с использованием других наборов геномных данных
Гены, имеющие общие известные функции, вероятно, будут иметь сходные генетические
профили взаимодействия. Поэтому генетические взаимодействия можно предсказать, используя
дополнительные наборы геномных данных, которые связывают гены в соответствии с их функциями.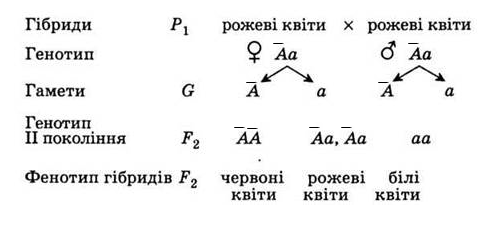 За
Например, генетические взаимодействия генов, кодирующих белки, которые физически
взаимодействовать были успешно предсказаны
(Келли и Идекер, 2005;
Ye et al., 2005). Однако
более эффективный подход — объединить несколько разных наборов данных, которые соединяют
функционально родственные гены и использовать их для прогнозирования генетических взаимодействий.
Здесь я обсуждаю два подхода, которые применялись на сегодняшний день: решение
деревья и байесовская интеграция.
За
Например, генетические взаимодействия генов, кодирующих белки, которые физически
взаимодействовать были успешно предсказаны
(Келли и Идекер, 2005;
Ye et al., 2005). Однако
более эффективный подход — объединить несколько разных наборов данных, которые соединяют
функционально родственные гены и использовать их для прогнозирования генетических взаимодействий.
Здесь я обсуждаю два подхода, которые применялись на сегодняшний день: решение
деревья и байесовская интеграция.
Деревья решений предоставляют метод классификации данных на два или более
классы (здесь «взаимодействующие» и «невзаимодействующие»), использующие несколько различных
Типы доказательств.На каждом шаге дерева список генов делится на те, которые
которые обладают или не обладают определенной характеристикой. На вершине дерева
список генов сначала разбивается с использованием наиболее информативной характеристики
для прогнозирования интересующего свойства (здесь способность прогнозировать генетический
взаимодействие). Затем дополнительные характеристики используются для создания дополнительных
подразделения списка генов до тех пор, пока не будет
информационная и ветка прекращается. Преимущество деревьев решений перед
методы «черного ящика», такие как нейронные сети и опорные векторные машины.
что они явно раскрывают характеристики, используемые для классификации данных.Они также не предполагают независимости между типами предиктивных доказательств, которые
позволяет использовать несколько связанных наборов данных, даже если они содержат корреляции
друг с другом. Wong et al. использовали деревья решений для интеграции белка
локализация, экспрессия мРНК, физическое взаимодействие, известная функция и
данные топологии сети для прогнозирования смертельного исхода или болезни
взаимодействия между генами дрожжей (Wong et al.
др., 2004). Используя тесты перекрестной проверки, они обнаружили, что решение
деревья могут надежно предсказать генетические взаимодействия между генами дрожжей.Они
также проверили прогнозы для восьми новых экранов SGA, которые не использовались при обучении;
49/318 прогнозов подтвердились по сравнению с 2/318, ожидаемыми случайно.
Затем дополнительные характеристики используются для создания дополнительных
подразделения списка генов до тех пор, пока не будет
информационная и ветка прекращается. Преимущество деревьев решений перед
методы «черного ящика», такие как нейронные сети и опорные векторные машины.
что они явно раскрывают характеристики, используемые для классификации данных.Они также не предполагают независимости между типами предиктивных доказательств, которые
позволяет использовать несколько связанных наборов данных, даже если они содержат корреляции
друг с другом. Wong et al. использовали деревья решений для интеграции белка
локализация, экспрессия мРНК, физическое взаимодействие, известная функция и
данные топологии сети для прогнозирования смертельного исхода или болезни
взаимодействия между генами дрожжей (Wong et al.
др., 2004). Используя тесты перекрестной проверки, они обнаружили, что решение
деревья могут надежно предсказать генетические взаимодействия между генами дрожжей.Они
также проверили прогнозы для восьми новых экранов SGA, которые не использовались при обучении;
49/318 прогнозов подтвердились по сравнению с 2/318, ожидаемыми случайно. (Wong et al., 2004). Вершина
предиктором в дереве решений было ранее отмеченное свойство сети, которое
гены, которые имеют общих партнеров по генетическому взаимодействию, также могут взаимодействовать
генетически. Также было обнаружено, что «межпутные» модели являются полезными предикторами
взаимодействий. Однако, отдельно исключая другие виды функциональных данных
само по себе мало влияло на качество прогнозов
(Вонг и др., 2004).
(Wong et al., 2004). Вершина
предиктором в дереве решений было ранее отмеченное свойство сети, которое
гены, которые имеют общих партнеров по генетическому взаимодействию, также могут взаимодействовать
генетически. Также было обнаружено, что «межпутные» модели являются полезными предикторами
взаимодействий. Однако, отдельно исключая другие виды функциональных данных
само по себе мало влияло на качество прогнозов
(Вонг и др., 2004).
Альтернативный метод интеграции наборов геномных данных для прогнозирования генетических
взаимодействия — это байесовская интеграция. В этом подходе прогнозы, сделанные с использованием
разные типы данных взвешиваются в зависимости от возможностей каждого типа данных
для прогнозирования известных генетических взаимодействий, в отличие от генов, которые неизвестны
взаимодействовать (Jansen et al.,
2003; Ли и др.,
2004; Троянская и др.,
2003 г.). Чжун и Штернберг недавно использовали этот подход для прогнозирования
генетические взаимодействия для ∼10% C. elegans , используя
информация об анатомических паттернах экспрессии, фенотипах, функциональных
аннотации, коэкспрессия микрочипов и белковые взаимодействия для прогнозирования
генетические взаимодействия (Чжун и Штернберг,
2006). Авторы использовали свою сеть, чтобы выявить двенадцать тонких
модификаторы мутаций в гене let-60 Ras и два новых
супрессоры мутаций в гене itr-1 . Использование модифицированного байесовского
интеграции, мы расширили этот подход, чтобы построить сеть из> 100
000 взаимодействий, которые покрывают> 60% из C.elegans генов и имеют
использовали эту сеть для выявления новых супрессоров мутаций в
Путь ретинобластомы и перекрестная связь между комплексом дистрофина и
Путь EGF / Ras / MAPK (I. Lee, B.L., C. Crombie, A. Fraser и E. M. Marcotte,
неопубликованные данные).
elegans , используя
информация об анатомических паттернах экспрессии, фенотипах, функциональных
аннотации, коэкспрессия микрочипов и белковые взаимодействия для прогнозирования
генетические взаимодействия (Чжун и Штернберг,
2006). Авторы использовали свою сеть, чтобы выявить двенадцать тонких
модификаторы мутаций в гене let-60 Ras и два новых
супрессоры мутаций в гене itr-1 . Использование модифицированного байесовского
интеграции, мы расширили этот подход, чтобы построить сеть из> 100
000 взаимодействий, которые покрывают> 60% из C.elegans генов и имеют
использовали эту сеть для выявления новых супрессоров мутаций в
Путь ретинобластомы и перекрестная связь между комплексом дистрофина и
Путь EGF / Ras / MAPK (I. Lee, B.L., C. Crombie, A. Fraser и E. M. Marcotte,
неопубликованные данные).
Прогнозирование генетических взаимодействий с использованием ортологических отношений
Последний подход к прогнозированию генетических взаимодействий — использование генетических
взаимодействия, выявленные у одного вида, для прогнозирования генетических взаимодействий в
второй вид. Эволюционная консервация белок-белок
взаимодействия между видами (Matthews et al.
al., 2001), означает, что белок-белковые взаимодействия человека
можно успешно предсказать, используя данные модельных организмов
(Ленер и Фрейзер, 2004). К
проверить, могут ли генетические взаимодействия успешно передаваться между
видов, мы проверили, являются ли ортологи генов синтетически
летальные для S. cerevisiae также синтетически летальны для C.
elegans . Всего мы проверили> 1000 предсказанных взаимодействий, но
обнаружили, что консервативны <1% (J.Тишлер, Б. и А. Фрейзер,
неопубликованные данные). Это контрастирует с мутациями в отдельных генах; > 60%
ортологов генов, которые необходимы для S. cerevisiae , являются
также необходим в C. elegans (Камат и др., 2003).
Следовательно, по крайней мере, для синтетических летальных взаимодействий, взаимодействия
вероятно, не сохраняется напрямую между одноклеточными и многоклеточными
организмы. Это может отражать наличие дополнительных компенсаторных путей.
Эволюционная консервация белок-белок
взаимодействия между видами (Matthews et al.
al., 2001), означает, что белок-белковые взаимодействия человека
можно успешно предсказать, используя данные модельных организмов
(Ленер и Фрейзер, 2004). К
проверить, могут ли генетические взаимодействия успешно передаваться между
видов, мы проверили, являются ли ортологи генов синтетически
летальные для S. cerevisiae также синтетически летальны для C.
elegans . Всего мы проверили> 1000 предсказанных взаимодействий, но
обнаружили, что консервативны <1% (J.Тишлер, Б. и А. Фрейзер,
неопубликованные данные). Это контрастирует с мутациями в отдельных генах; > 60%
ортологов генов, которые необходимы для S. cerevisiae , являются
также необходим в C. elegans (Камат и др., 2003).
Следовательно, по крайней мере, для синтетических летальных взаимодействий, взаимодействия
вероятно, не сохраняется напрямую между одноклеточными и многоклеточными
организмы. Это может отражать наличие дополнительных компенсаторных путей. у высших эукариот или, наоборот, предполагает, что
эволюционный отбор для взаимодействий «более высокого порядка» между мутациями в
пары генов.Однако это не означает, что синтетический смертельный
взаимодействия между генами дрожжей неинформативны для предсказания генов человека
функции — очевидно, что синтетические летальные профили генов дрожжей
очень информативен для прогнозирования молекулярных функций ортологичного человека
гены (Pan et al., 2006) и
эту информацию можно использовать для прогнозирования генетических взаимодействий у людей. В
в будущей работе будет важно проверить, могут ли генетические взаимодействия быть
успешно передан между более родственными видами (например, между С.elegans и людей). Анекдотические примеры из литературы [для
пример сохранения генетических взаимодействий между компонентами
Путь EGF / Ras / MAPK между червями и мухами
(Сундарам, 2005)], предлагаем
что это должно быть более успешным. Дополнительно можно предусмотреть
более сложные подходы, при которых взаимодействие между путями, скорее
чем гены, предсказываются.
у высших эукариот или, наоборот, предполагает, что
эволюционный отбор для взаимодействий «более высокого порядка» между мутациями в
пары генов.Однако это не означает, что синтетический смертельный
взаимодействия между генами дрожжей неинформативны для предсказания генов человека
функции — очевидно, что синтетические летальные профили генов дрожжей
очень информативен для прогнозирования молекулярных функций ортологичного человека
гены (Pan et al., 2006) и
эту информацию можно использовать для прогнозирования генетических взаимодействий у людей. В
в будущей работе будет важно проверить, могут ли генетические взаимодействия быть
успешно передан между более родственными видами (например, между С.elegans и людей). Анекдотические примеры из литературы [для
пример сохранения генетических взаимодействий между компонентами
Путь EGF / Ras / MAPK между червями и мухами
(Сундарам, 2005)], предлагаем
что это должно быть более успешным. Дополнительно можно предусмотреть
более сложные подходы, при которых взаимодействие между путями, скорее
чем гены, предсказываются.
Значение сетей генетического взаимодействия для болезней человека: концентраторы, буферы и новые парадигмы болезней человека
Каковы последствия сетей генетического взаимодействия модельных организмов?
для генетических заболеваний человека? Самое непосредственное наблюдение — генетический
сети взаимодействия очень плотные — есть еще много способов
производят фенотип путем объединения мутаций в двух генах, чем путем мутации
единственный ген (Тонг и др.,
2004 г.). Это говорит о том, что большинство наследственных заболеваний также вероятны.
в основном в результате взаимодействия между мутациями в нескольких генах
а не от мутаций в отдельных генах
(Бадано и Кацанис, 2002).
Действительно, преобладание синтетических взаимодействий между генами может объяснить
относительное отсутствие успеха в выявлении причинных мутаций в наиболее распространенных
наследственные заболевания — каждое заболевание может возникнуть в результате множества различных
комбинации мутаций во многих различных генах, каждый из которых имеет только
незначительное или нулевое влияние только на болезнь. Это рисует довольно пессимистичный
картинка для практических целей генетического препарирования и лечения комплекса
генетические заболевания.
Это рисует довольно пессимистичный
картинка для практических целей генетического препарирования и лечения комплекса
генетические заболевания.
Однако структура сетей генетического взаимодействия модельных организмов также
имеет положительное значение для генетики болезней человека — наличие
сильно связанные гены «концентратора» в сетях предполагают, что будут
общие модификаторы генетических мутаций у человека. Например, в C.
elegans в сети генетического взаимодействия мы определили класс генов,
генетически взаимодействуют со многими разнообразными генами.Инактивация каждого из этих
гены-концентраторы могут усиливать фенотип потери функции в результате
мутации во множестве разных генов с разными молекулярными функциями
(Рис. 3). Действительно, потеря хаба
ген может усиливать множество различных фенотипов, в зависимости от другого гена, который
мутирует в комбинации. Таким образом, их можно рассматривать как «буферизацию»
организм от фенотипических последствий мутаций. Замечательно все
наиболее связанные гены-концентраторы, которые мы определили, функционируют в
комплексы, модифицирующие хроматин (Lehner et al.
al., 2006b).
Замечательно все
наиболее связанные гены-концентраторы, которые мы определили, функционируют в
комплексы, модифицирующие хроматин (Lehner et al.
al., 2006b).
Генетические узлы и генетические заболевания человека. Генетические узлы — это гены, которые
в инактивированном состоянии может усилить фенотипические последствия мутаций во многих
разные гены. Часто гены-концентраторы могут усиливать последствия мутаций в
гены, действующие в различных функционально несвязанных путях. Примеры включают
набор генов, модифицирующих хроматин, в C. elegans (гены mys-1,
trr-1, dpy-22, hmg-1.2, din-1 и egl-27 )
(Ленер и др., 2006b),
Комплекс префолдин в S. cerevisiae (Tong et al., 2004), а
ген hsp90 в дрожжах (Zhao et al.,
2005), летает (Резерфорд и
Линдквист, 1998) и растения
(Queitsch et al., 2002). Здесь
красный узел представляет собой ген концентратора, а остальные узлы окрашены
в соответствии с их функцией. Белковые взаимодействия показаны как
сплошные линии и генетические взаимодействия пунктирными линиями.
Вероятно, есть еще много генов-концентраторов в дополнение к этим гены, модифицирующие хроматин.Например, в генетическом взаимодействии дрожжей SGA сети, четыре из пяти наиболее связанных генов кодируют компоненты префолдин-шапероновый комплекс (Tong et al., 2004 г.). Более того, шаперон hsp90 также может быть отнесен к классу в качестве гена-концентратора из-за его способности усиливать фенотипические последствия мутаций в нескольких генах при инактивации у мух, растений и дрожжей (Queitsch et al., 2002; Резерфорд и Линдквист, 1998; Чжао и др., 2005). Таким образом, по крайней мере два класса генов могут функционировать как генетические. хабы: модификаторы хроматина и шапероны.Интересно hsp90 май соединить эти два функциональных класса; хотя он известен как сопровождающий, он также может влиять на фенотипические вариации через его влияние на хроматин структура (Sollars et al., 2003 г.).
Хотя есть вероятность, что многие другие гены-концентраторы еще
выявлено, их значение для болезней человека очевидно: вероятно
быть человеческими генами, которые действуют как гены-модификаторы во многих механистически не связанных
болезни. Действительно, концепция генов-концентраторов предлагает новую парадигму генетического
болезнь человека (Lehner et al.,
2006b). В этой парадигме есть два класса болезней человека.
ген: первый класс состоит из генов «спецификаторов», которые определяют конкретный
болезнь, а второй класс состоит из генов «модификаторов» или «концентраторов», которые служат
для усиления силы болезни, возникшей в результате мутации в
ген-спецификатор. Есть убедительные доказательства того, что гены-концентраторы, идентифицированные в
один организм также функционирует как центры у других видов
(Lehner et al., 2006b;
Queitsch et al., 2002;
Zhao et al., 2005), и поэтому
определенные гены, идентифицированные как центры в модельных организмах, также могут функционировать как
гены-концентраторы у людей.
Действительно, концепция генов-концентраторов предлагает новую парадигму генетического
болезнь человека (Lehner et al.,
2006b). В этой парадигме есть два класса болезней человека.
ген: первый класс состоит из генов «спецификаторов», которые определяют конкретный
болезнь, а второй класс состоит из генов «модификаторов» или «концентраторов», которые служат
для усиления силы болезни, возникшей в результате мутации в
ген-спецификатор. Есть убедительные доказательства того, что гены-концентраторы, идентифицированные в
один организм также функционирует как центры у других видов
(Lehner et al., 2006b;
Queitsch et al., 2002;
Zhao et al., 2005), и поэтому
определенные гены, идентифицированные как центры в модельных организмах, также могут функционировать как
гены-концентраторы у людей.
БЛАГОДАРНОСТИ
Работа в моей лаборатории финансируется Программой системной биологии EMBL-CRG, которая
при поддержке гранта Министерства науки и образования Испании
(Ministryio de Educación y Ciencia, MEC).
- © Компания Биологов Лимитед 2007
Ссылки
- ↵
Бадано, Дж. Л. и Кацанис, Н. (2002). Вне Мендель: развивающийся взгляд на передачу генетических заболеваний человека.Nat. Преподобный Жене. 3779 -789.
- ↵
Боуг, Л. Р., Вэнь, Дж. К., Хилл, А. А., Слоним, Д. К., Браун, Э. Л. и Хантер, С. П. (2005). Синтетический летальный анализ Caenorhabditis elegans гены формирования заднего эмбриона определяет консервативные генетические взаимодействия. Геном Биол. 6, R45 .
- ↵
Коллинз, С. Р., Шульдинер, М., Кроган, Н. Дж. И Вайсман, Дж. С. (2006).Стратегия извлечения и анализа крупномасштабные количественные данные эпистатического взаимодействия. Геном Биол. 7, R63 .
- ↵
Davierwala, A. P., Haynes, J., Li, Z., Brost, R.
L., Robinson,
M. D., Yu, L., Mnaimneh, S., Ding, H., Zhu, H., Chen, Y. et al. (2005). Спектр синтетического генетического взаимодействия основных
гены. Nat. Genet. 37,1147
-1152. - ↵
Дриес, Б.Л., Торссон, В., Картер, Дж. У., Ривз, А. У., Раймонд М.З., Авила-Кампилло И., Шеннон П. и Галицкий Т. (2005). Вывод сетей генетического взаимодействия из количественные данные фенотипа. Genome Biol. 6, R38.
- ↵
Исон, Р. Г., Пурманд, Н., Тонгпрасит, В., Герман, З. С., Энтони, К., Джеджелоу, О., Дэвис, Р. В. и Столк, В. (2004). Характеристика штрих-кодов синтетической ДНК в сахаромицете cerevisiae штаммов с делецией гена.Proc. Natl. Акад. Sci. США 101,11046 -11051.
- ↵
Джавер, Г., Чу, А. М., Ни, Л., Коннелли, К., Райлз, Л., Veronneau, S., Dow, S., Lucau-Danila, A., Anderson, K., Andre, B.
et al. (2002). Функциональное профилирование сахаромицетов cerevisiae геном. Природа
418 387
-391. - ↵
Hartman, J. L., 4th and Tippery, N. P. (2004). Систематическая количественная оценка взаимодействий генов с помощью анализа фенотипического массива.Genome Biol. 5, R49 .
- ↵
Хартман, Дж. Л., 4-й, Гарвик, Б. и Хартвелл, Л. (2001). Принципы буферизации генетической изменчивости. Наука 291,1001 -1004.
- ↵
Холуэй, А. Х., Хунг, К. и Майкл, У. М. (2005). Систематическая идентификация с помощью РНК-интерференции Гены-модификаторы mus-101 у Caenorhabditis elegans. Генетика 169,1451 -1460.
- ↵
Янсен Р., Ю., Х., Гринбаум, Д., Клюгер, Ю., Кроган, Н. Дж., Чанг, С., Эмили, А., Снайдер, М., Гринблатт, Дж. Ф. и Герштейн, М. (2003). Подход байесовских сетей для прогнозирования белок-белковые взаимодействия из геномных данных. Наука 302 449 -453.
- ↵
Камат Р. С., Фрейзер А. Г., Донг Ю., Пулин Г., Дурбин Р., Gotta, M., Kanapin, A., Le Bot, N., Moreno, S., Sohrmann, M. et al. (2003). Систематический функциональный анализ Caenorhabditis elegans с использованием РНКи.Природа 421 231 -237.
- ↵
Келли Р. и Идекер Т. (2005). Систематический интерпретация генетических взаимодействий с использованием белковых сетей. Nat. Biotechnol. 23 561 -566.
- ↵
Кулькарни, М. М., Букер, М., Сильвер, С. Дж., Фридман, А., Хонг, П., Перримон Н. и Мэти-Превот Б. (2006). Доказательство того нецелевые эффекты, связанные с длинными дцРНК у Drosophila melanogaster клеточные анализы.Nat. Методы 3, 833-838.
- ↵
Ли, И., Дате, С. В., Адай, А. Т. и Маркотт, Э. М. (2004). Вероятностная функциональная сеть дрожжевых генов. Наука 306,1555 -1558.
- ↵
Ленер Б. и Фрейзер А. Г. (2004). А первый набросок карты взаимодействия белков человека. Геном Биол. 5, R63 .
- ↵
Ленер, Б., Фрейзер, А.Г. и Сандерсон, К.М. (2004). Обзор методики: как использовать РНК-интерференцию. Краткий. Функц. Геномный. Протеомный. 3, 68-83.
- ↵
Ленер, Б., Каликсто, А., Кромби, К., Тишлер, Дж., Фортунато, А., Чалфи М. и Фрейзер А. Г. (2006a). Потеря LIN-35, Caenorhabditis elegans супрессора опухолей p105Rb, приводит к усиленная интерференция РНК. Genome Biol. 7, R4.
- ↵
Ленер, Б., Кромби, К., Тишлер, Дж., Фортунато, А. и Фрейзер, A. G. (2006b). Систематическое картирование генетических взаимодействий в Caenorhabditis elegans определяет общие модификаторы различных сигнальные пути. Nat. Genet. 38 896 -903.
- ↵
Ленер, Б., Тишлер, Дж. И Фрейзер, А. Г. (2006c). РНКи скрининг в C. elegans в 96-луночной жидкости формат и их применение для систематической идентификации генетических взаимодействия.Nat. Protoc. 1,1617 -1620.
- ↵
Ma, Y., Creanga, A., Lum, L. and Beachy, P.A. (2006). Преобладание нецелевых эффектов в РНК дрозофилы интерференционные экраны. Природа 443 359 -363.
- ↵
Мэтьюз, Л. Р., Вальо, П., Ребул, Дж., Ге, Х., Дэвис, Б. П., Гаррелс, Дж., Винсент, С. и Видаль, М. (2001). Идентификация потенциальных сетей взаимодействия с использованием последовательного поиска сохраненных белок-белковые взаимодействия или «интерологи».Геном Res. 11 2120 -2126.
- ↵
Оои, С. Л., Шумейкер, Д. Д. и Боке, Дж. Д. (2003). Сеть взаимодействия генов ДНК-геликазы, определенная с помощью синтетическая летальность анализируется микрочипом. Nat. Genet. 35 277 -286.
- ↵
Ooi, S. L., Pan, X., Peyser, B. D., Ye, P., Meluh, P. B., Yuan, Д. С., Иризарри, Р. А., Бадер, Дж. С., Спенсер, Ф. А. и Боке, Д. Д. (2006). Общий анализ синтетической летальности и дрожжевой функционал профилирование.Тенденции Genet. 22, 56-63.
- ↵
Пан, X., Юань, D. S., Xiang, D., Wang, X., Sookhai-Mahadeo, S., Бадер, Дж. С., Хитер, П., Спенсер, Ф. и Боке, Дж. Д. (2004). Надежный набор инструментов для функционального профилирования дрожжей геном. Мол. Ячейка 16,487 -496.
- ↵
Пан, X., Ye, P., Yuan, D. S., Wang, X., Bader, J. S. и Boeke, Дж. Д. (2006). Сеть целостности ДНК в дрожжах Saccharomyces cerevisiae. Ячейка 124 1069 -1081.
- ↵
Пей, Ю. и Тушл, Т. (2006). Об искусстве выявление эффективных и специфических миРНК. Nat. Методы 3670 -676.
- ↵
Пирс, С. Э., Фунг, Э. Л., Джарамилло, Д. Ф., Чу, А. М., Дэвис, Р. У., Нислоу, К. и Дживер, G. (2006). Уникальный и универсальный молекулярный массив штрих-кода. Nat. Методы 3, 601-603.
- ↵
Queitsch, C., Сангстер Т. А. и Линдквист С. (2002). Hsp90 как конденсатор фенотипической изменчивости. Природа 417 618 -624.
- ↵
Rutherford, S. L. и Lindquist, S. (1998). Hsp90 как конденсатор для морфологической эволюции. Природа 396 336 -342.
- ↵
Шульдинер М., Коллинз С. Р., Томпсон Н. Дж., Деник В., Бхамидипати А., Пунна Т., Ихмелс Дж., Эндрюс Б., Бун К., Гринблатт Дж.F. et al. (2005). Исследование функции и организация раннего секреторного пути дрожжей через эпистатический профиль miniarray. Клетка 123 507 -519.
- ↵
Солларс, В., Лу, Х., Сяо, Л., Ван, Х., Гарфинкель, М. Д. и Руден, Д. М. (2003). Доказательства эпигенетического механизма который Hsp90 действует как конденсатор для морфологической эволюции. Nat. Genet. 33,70 -74.
- ↵
- ↵
Suzuki, Y.и Хан М. (2006). Генетический избыточность маскирует различные функции гена супрессора опухолей PTEN во время C. elegans разработка. Genes Dev. 20 423 -428.
- ↵
Тиммонс, Л. и Файер, А. (1998). Специфический вмешательство проглоченной дцРНК. Природа 395, 854.
- ↵
Тишлер Дж., Ленер Б., Чен Н. и Фрейзер А. Г. (2006). Комбинаторная РНК-интерференция у C.elegans показывает, что избыточность между дубликатами генов может поддерживаться для большего более 80 миллионов лет эволюции. Genome Biol. 7, R69.
- ↵
Тонг, А. Х., Евангелиста, М., Парсонс, А. Б., Сюй, Х., Бадер, Г. Д., Пейдж, Н., Робинсон, М., Рагибизаде, С., Хог, К. В., Бусси, Х. и др. al. (2001). Систематический генетический анализ с упорядоченными массивами делеционных мутантов дрожжей. Наука 294,2364 -2368.
- ↵
Тонг, А.Х., Лесаж, Г., Бадер, Г. Д., Дин, Х., Сюй, Х., Синь, X., Young, J., Berriz, G.F., Brost, R.L., Chang, M. et al. (2004). Глобальное картирование сети генетического взаимодействия дрожжей. Наука 303 808 -813.
- ↵
Троянская, О. Г., Долински, К., Оуэн, А. Б., Альтман, Р. Б. и Ботштейн, Д. (2003). Байесовская структура для комбинирования гетерогенные источники данных для предсказания функции генов (в Saccharomyces cerevisiae ).Proc. Natl. Акад. Sci. Соединенные Штаты Америки 100,8348 -8353.
- ↵
van Haaften, G., Vastenhouw, N.L., Nollen, E.A., Plasterk, R. Х. и Тийстерман, М. (2004). Взаимодействие генов в ДНК Путь повреждение-ответ, идентифицированный с помощью анализа РНК-интерференции всего генома синтетическая летальность. Proc. Natl. Акад. Sci. Соединенные Штаты Америки 101,12992 -12996.
- ↵
Вагнер А. (2005). Распределенная надежность против избыточности как причины мутационной устойчивости.BioEssays 27,176 -188.
- ↵
Уиллер, Д. Б., Бейли, С. Н., Гертин, Д. А., Карпентер, А. Э., Хиггинс, К. О. и Сабатини, Д. М. (2004). РНКи живая клетка микрочипы для экранов потери функции в Drosophila melanogaster клетки. Nat. Методы 1,127 -132.
- ↵
Вонг, С. Л., Чжан, Л. В., Тонг, А. Х., Ли, З., Голдберг, Д. С., Кинг, О. Д., Лесаж, Г., Видаль, М., Эндрюс, Б., Bussey, H. et al. (2004). Объединение биологических сетей для прогнозирования генетических взаимодействия. Proc. Natl. Акад. Sci. Соединенные Штаты Америки 101,15682 -15687.
- ↵
Ye, P., Peyser, B.D., Pan, X., Boeke, J.D., Spencer, F.A. и Бадер, Дж. С. (2005). Прогнозирование функции генов от конгруэнтного синтетические летальные взаимодействия в дрожжах. Мол. Syst. Биол. 1, 2005 0026.
- ↵
Юань, Д.С., Пан, X., Оои, С.Л., Пейзер, Б. Д., Спенсер, Ф. А., Иризарри, Р. А. и Боке, Дж. Д. (2005). Улучшенный микрочип методы профилирования коллекции штаммов Yeast Knockout. Нуклеиновая Acids Res. 33, e103 .
- ↵
Чжао, Р., Дэйви, М., Сюй, Ю. К., Капланек, П., Тонг, А., Парсонс, А.Б., Кроган, Н., Кэгни, Г., Май, Д., Гринблатт, Дж. И др. (2005). Навигация по сети шаперонов: интегральная карта физические и генетические взаимодействия, опосредованные шапероном hsp90.Ячейка 120,715 -727.
- ↵
Чжун В. и Штернберг П. В. (2006). Полногеномное предсказание генетических взаимодействий C. elegans . Наука 311 1481 -1484.
 L., Robinson,
M. D., Yu, L., Mnaimneh, S., Ding, H., Zhu, H., Chen, Y. et al. (2005). Спектр синтетического генетического взаимодействия основных
гены. Nat. Genet. 37,1147
-1152.
L., Robinson,
M. D., Yu, L., Mnaimneh, S., Ding, H., Zhu, H., Chen, Y. et al. (2005). Спектр синтетического генетического взаимодействия основных
гены. Nat. Genet. 37,1147
-1152.  et al. (2002). Функциональное профилирование сахаромицетов cerevisiae геном. Природа
418 387
-391.
et al. (2002). Функциональное профилирование сахаромицетов cerevisiae геном. Природа
418 387
-391. Персонализированная терапия по фенотипу и генотипу
Введение
После публикации результатов 20-летнего проспективного исследования диабета в Великобритании (UKPDS) (1) общая рекомендация заключалась в том, чтобы достичь как можно большего числа пациентов. пациентов с диабетом 2 типа (СД2), целевое значение HbA 1c ≤7%, т.е.е., среднее значение, связанное со значительным снижением риска микро- и макрососудистых осложнений. Однако эта «универсальная» гликемическая цель была поставлена под сомнение в результате результатов последующих интервенционных исследований (2–4). Учитывая критический анализ результатов этих исследований, Американская диабетическая ассоциация (ADA) и Американская кардиологическая ассоциация предложили определить индивидуальную цель HbA 1c на основе продолжительности диабета, ожидаемой продолжительности жизни, наличия и степени тяжести диабета. осложнения и склонность к гипогликемии (5).Это предложение было одобрено несколькими международными руководящими принципами. Они также предлагают индивидуальный выбор стратегий снижения уровня глюкозы, чтобы обеспечить наиболее подходящее соотношение риска и пользы для всех пациентов. Согласно заявлению ADA / Европейской ассоциации по изучению диабета в отношении лечения СД2 (6), этого можно достичь путем сбалансированной оценки эффективности, риска гипогликемии, влияния на массу тела и стоимости доступных глюкозоснижающих средств и их сочетание.
Несмотря на то, что концепция индивидуализации лечения привлекательна, способы ее реализации в клинических условиях остаются предметом споров. Для этой цели может быть уместным тщательное фенотипическое описание человека с СД2. Несколько лет назад был предложен подход ABCD, чтобы помочь врачам индивидуализировать целевой уровень гликемии и выбрать лечение (7). ABCD обозначает четыре фенотипических признака, которые легко собрать и учесть: возраст, массу тела, осложнения и сопутствующие заболевания, а также продолжительность диабета.
Возраст
Возраст — самый сильный фактор риска для продолжительности жизни и смертности. Число пожилых людей с диабетом растет, и> 70% пожилых людей с СД2 имеют некоторую степень нарушения физических способностей. Лечение диабета у этих субъектов может быть проблематичным, так как оно может привести к риску ускорения развития гериатрического синдрома. Лечение должно быть направлено на защиту пациента от острых метаболических осложнений, предотвращение развития и / или прогрессирования осложнений диабета, сохранение когнитивных функций и физических возможностей, обеспечение надлежащего качества жизни и предотвращение реакций на лекарства.Поэтому при выборе глюкозоснижающих средств эти рекомендации следует полностью учитывать.
Недавно была опубликована критическая оценка глюкозоснижающей терапии (8). Хотя возраст сам по себе не является противопоказанием для приема метформина, его использование может быть ограничено из-за побочных эффектов со стороны желудочно-кишечного тракта, которые могут быть нежелательными для пожилых ослабленных пациентов. Метформин может снизить уровень витамина B 12 , что может ускорить когнитивную дисфункцию. Однако основным ограничением для использования препарата остается нарушение функции почек и риск развития лактоацидоза.
Пиоглитазон не противопоказан людям с пониженной скоростью клубочковой фильтрации (СКФ), но его использование ограничено повышенным риском переломов костей, задержкой жидкости и сердечной недостаточностью.
Основной проблемой, связанной с сульфонилмочевиной и глинидами, остается повышенный риск гипогликемии. Мы сообщали, что 40% пациентов, обращавшихся в отделение неотложной помощи из-за гипогликемии, принимали антидиабетические препараты с использованием сульфонилмочевины / глинидов ± сенсибилизаторы инсулина в 92% случаев, а средний возраст составлял 79 ± 11 лет (9).Глибенкламид был наиболее часто используемой сульфонилмочевиной (69%). Более того, среди пациентов, принимавших сульфонилмочевину, 47% имели расчетную СКФ <60 мл / мин / 1,73 м 2 и 13,5% имели <30 мл / мин / 1,73 м 2 . Это наблюдение подтверждает рекомендацию о том, что следует тщательно оценивать риск, связанный с стимуляторами секреции инсулина, особенно при назначении у уязвимых пациентов с СД2.
Испытания, проведенные с участием пожилых пациентов с СД2 (пересмотренный вариант в 10), подтвердили эффективность, нейтральный вес и низкий риск гипогликемии ингибиторов дипептидилпептидазы (DPP) -4.У пациентов с почечной недостаточностью средней и тяжелой степени соответствующая корректировка дозировки, за исключением линаглиптина, который не выводится почками, обеспечивает хорошее соотношение пользы и риска без дальнейшего ухудшения функции почек или увеличения частоты гипогликемии.
Данных по аналогам глюкагоноподобного пептида 1 (GLP-1) у пожилых пациентов с СД2 скуднее. Эти агенты имеют благоприятное соотношение пользы и риска, но их применение у пожилых людей может быть ограничено сопутствующим нарушением функции почек.Агонисты GLP-1, хотя и с некоторыми различиями внутри класса, могут вызывать тошноту и рвоту, что может противопоказать их применение у пожилых пациентов с более низкой массой тела, потерей веса или неравномерным кормлением.
Эффективность ингибиторов натрий-глюкозного котранспортера (SGLT) 2, вероятно, будет самоограничиваться у пациентов со сниженной расчетной СКФ, так как это снизит канальцевую нагрузку и, следовательно, экскрецию глюкозы. Хотя обычно считается, что эти соединения обладают удовлетворительным соотношением эффективности и безопасности, риск обезвоживания и гипотонии выше у пожилых людей, у тех, кто принимает петлевые диуретики, и у людей с нарушенной функцией почек.
Наконец, инсулин остается эффективной формой лечения, хотя необходимы целенаправленные испытания инсулинотерапии у пожилых людей. Риск гипогликемии является самым высоким при приеме инсулина, и это может стать особенно проблемным для ослабленных пожилых пациентов, не имеющих поддержки.
Масса тела
Масса тела и распределение жировой ткани являются общими фенотипическими признаками и потенциальными маркерами инсулинорезистентности и метаболического синдрома. Из 1598 пациентов с СД2, обратившихся в нашу клинику, у 50% ИМТ> 27 кг / м 2 , у 27% ИМТ> 30 кг / м 2 , а у 13% ИМТ> 30 кг / м 2 .При этом у 69% окружность талии> 100 см. В общей популяции было совершенно очевидно, что чем больше ИМТ, тем выше HbA 1c , уровни триглицеридов, систолическое и диастолическое артериальное давление и тем ниже концентрация холестерина ЛПВП. Следовательно, при индивидуальном подходе к лечению вес тела может служить ориентиром (рис. 1). У пациентов с ожирением II и III классов (т.е. ИМТ от 35 до <40 и ≥40 кг / м 2 ) снижение массы тела является основной целью наряду с восстановлением гликемического контроля.Следовательно, лечение, связанное с потерей веса, то есть агонисты рецептора GLP-1 и ингибиторы SGLT2, особенно если их используют в сочетании с метформином, может быть предпочтительным выбором. На противоположной стороне распределения ИМТ несколько более стройных пациентов с СД2, у которых можно предположить серьезный дефект функции β-клеток, могут быть лучшими кандидатами для более ранней инсулиновой терапии. Наконец, у субъектов с диабетом со средней степенью ожирения (ИМТ от 30 до <35 кг / м2 2 ), у которых риски, связанные с ожирением, все еще остаются предметом обсуждения (11), основной целью должно быть предотвращение увеличения веса или стимулирование умеренного ожирения. потеря веса.Это говорит о том, что следует избегать лекарств, которые могут способствовать увеличению веса (например, сульфонилмочевины, пиоглитазона и инсулина), в то время как отдавать предпочтение агентам, не влияющим на вес, таким как ингибиторы DPP-4 или агентам, которые могут вызывать умеренную, но устойчивую потерю веса, такие как ингибиторы SGLT2 (12).
Рис. 1Схематическая рекомендация о том, как масса тела может определять индивидуализацию глюкозоснижающей терапии. РА, агонисты рецепторов; СУ, сульфонилмочевины.
Осложнения
Наличие осложнений диабета требует тщательной оценки фармакологической терапии, снижающей уровень глюкозы.Типичным примером является наличие диабетической нефропатии и потери функции почек, так как количество агентов необходимо либо уменьшить в дозах, либо не использовать. Из многих терапевтических вариантов только инсулин, пиоглитазон и ингибиторы ДПП-4 могут использоваться на всех стадиях почечной функции. Риск гипогликемии высок при приеме инсулина, поэтому требуется тщательная корректировка дозы. Пиоглитазон не выводится через почки, поэтому его можно использовать без ограничений, за исключением риска перелома костей и задержки жидкости, состояний, при которых люди с низкой СКФ уже находятся в группе повышенного риска (8).Ингибиторы ДПП-4 при соответствующей корректировке доз, за исключением линаглиптина, сохраняют эффективность, сравнимую с эффективностью сульфонилмочевины, при почти полном отсутствии риска гипогликемии у пациентов с почечной недостаточностью средней и тяжелой степени (13). Подобные результаты были получены у пациентов с терминальной стадией почечной недостаточности, получающих диализ (14).
Помимо соображений безопасности, врач может также рассмотреть дополнительные преимущества фармакологического агента в отношении предотвращения осложнений.Например, в ряде исследований было обнаружено, что использование ингибиторов DPP-4 связано со снижением альбуминурии (rev. В 15). Эти наблюдения находят основание для идентификации DPP-4 в почках, где он может участвовать в метаболизме нейропептида Y и пептида YY, пролиферации клеток и выработке коллагена прегломерулярными и мезангиальными клетками (16). Рецепторы GLP-1 также экспрессируются в почках, где они могут проявлять натрийуретические и диуретические свойства (16).
Еще больший интерес существует в отношении риска сердечно-сосудистых заболеваний (ССЗ), поскольку метформин по-прежнему обладает некоторой защитой от сердечно-сосудистых заболеваний.PRO-проспективное клиническое испытание пиоглитазона при макрососудистых событиях (PROactive) предоставляет убедительные доказательства благоприятного воздействия пиоглитазона на сердечно-сосудистые исходы (17). Считается, что инсулин свободен от риска, поскольку он используется у лиц из группы высокого риска с ранней стадией СД2 (18), хотя в свете недавнего анализа все же могут потребоваться более целенаправленные исследования (19).
Большие ожидания вызвали исследования результатов сердечно-сосудистых заболеваний, основанные на ингибиторах DPP-4 и агонистах рецепторов GLP-1, из-за их эффективности и сопутствующего воздействия на сердечно-сосудистую систему (20).Доклинические исследования с GLP-1 и его аналогами показали уменьшение площади инфаркта при ишемии (20). Ингибирование DPP-4 может обеспечивать защиту от сердечно-сосудистых заболеваний не только путем ингибирования деградации GLP-1, но также путем блокирования деградации хемокинов и фактора-1α, производного от стромальных клеток, с увеличением количества эндотелиальных клеток-предшественников (15). Результаты SAVOR-TIMI (оценка саксаглиптином сосудистых исходов, зарегистрированных у пациентов с сахарным диабетом — тромболизис при инфаркте миокарда) (21), исследования сердечно-сосудистых исходов с применением алоглиптина по сравнению со стандартом лечения (EXAMINE) (22) и TECOS (Trial Evaluating) Сердечно-сосудистые исходы при применении ситаглиптина) (23), оценивая, соответственно, эффект саксаглиптина, алоглиптина и ситаглиптина, и результаты ELIXA (24), где был протестирован ликсисенатид, предоставили доказательства их безопасного применения у пациентов с СД2 с высоким риском сердечно-сосудистых заболеваний. .Снижения сердечно-сосудистых событий или смертности не было обнаружено, хотя следует иметь в виду, что это были исследования не меньшей эффективности, как того требуют регулирующие органы для всех новых лекарств от диабета.
К настоящему времени завершенные исследования исходов сердечно-сосудистых заболеваний подтверждают безопасное использование инкретиновой терапии у пациентов с СД2 с высоким риском сердечно-сосудистых заболеваний, но оставляют без ответа вопрос о том, может ли их использование на ранних стадиях заболевания также обеспечить защиту. Наконец, повышенный риск госпитализации по поводу сердечной недостаточности, наблюдаемый в SAVOR-TIMI (21), но не в EXAMINE (22) и TECOS (23), потребует дополнительной оценки, чтобы установить, является ли это эффектом лекарственного препарата или может объясняться различиями в дизайне или проведении исследований.
Ингибиторы SGLT2 обладают потенциалом снижения риска сердечно-сосудистых заболеваний, учитывая их глюкозоснижающую эффективность, связанную с устойчивой умеренной потерей веса, снижением артериального давления и уровня мочевой кислоты (11). В поддержку потенциальной защиты от сердечно-сосудистых заболеваний результаты первого исследования сердечно-сосудистых заболеваний, в котором сравнивали эмпаглифлозин с плацебо в дополнение к стандартному лечению, показали сильное снижение риска сердечно-сосудистой смертности и смертности от всех причин у людей с СД2 высокого риска (25). У пациентов с СД2 с предшествующими сердечно-сосудистыми событиями лечение эмпаглифлозином в дополнение к стандартной терапии было связано со снижением риска на 16% (95% ДИ 0.79–0,99, P <0,04) для комбинированной конечной точки смертности от ССЗ, нефатального инфаркта миокарда и нефатального инсульта. Более того, было зарегистрировано значительное снижение риска смерти от сердечно-сосудистых причин (-38%), госпитализации по поводу сердечной недостаточности (-35%) и смерти от любой причины (-32%). Остается выяснить, применимы ли эти результаты к общей популяции с диабетом (т. Е. К людям с более низким риском сердечно-сосудистых заболеваний), но их следует учитывать при работе с пациентами с СД2, у которых уже было сердечно-сосудистое событие.
В заключение, в процессе индивидуализации пункт «осложнения» является сложным, поскольку он требует установления наличия и тяжести осложнений и относительной безопасности фармакологических методов лечения, а также оценки их потенциального вклада в предотвращение осложнений, помимо их сахароснижающего эффекта. .
Продолжительность диабета
Чем больше продолжительность диабета, тем выше вероятность осложнений и тем меньше продолжительность жизни. С этой точки зрения последствия давнего диабета уже обсуждались.Остается прокомментировать случай кратковременной продолжительности и, по умолчанию, всех случаев впервые диагностированного СД2. Результаты, полученные у таких пациентов в UKPDS (1), подтверждают рекомендацию о достижении строгого и стойкого гликемического контроля. Однако на сегодняшний день имеется мало информации о демографических, клинических и других факторах, которые могут повлиять на реакцию на лекарства, что затрудняет принятие решения о выборе лекарства. Метформин повсеместно считается препаратом первой линии. Более неясно, как действовать при неэффективности метформина.Некоторые ответы будут получены из GRADE (Подходы к снижению гликемии при диабете: сравнительное исследование эффективности), в котором сравнивается сульфонилмочевина, ингибитор DPP-4, агонист рецептора GLP-1 и базальный инсулин у пациентов с недавно диагностированным T2DM, получавших метформин, для изучения их эффективности в поддержание целевого уровня гликемии (HbA 1c <7%) в течение долгого времени (26). Другие исходы будут включать относительное влияние на отдельные микрососудистые осложнения и факторы риска сердечно-сосудистых заболеваний, исходы, ориентированные на пациента, и экономическую эффективность.GRADE также оценит фенотипические характеристики, связанные с успехом, неудачей и побочными эффектами различных комбинаций лекарств (26).
Таким образом, подход ABCD может идентифицировать общие фенотипы, которые будут использоваться при выборе фармакологического лечения. Тем не менее, их может быть недостаточно, если не будет добавлена следующая буква алфавита. В данном случае E означает «этиологию», то есть идентификацию основных патофизиологических механизмов, ответственных за прогрессирование заболевания у данного человека.
Этиология заболевания
Лучшее понимание патофизиологических механизмов может помочь в определении наилучшей стратегии лечения СД2. В последние годы, наряду с инсулинорезистентностью и нарушением функции β-клеток, был идентифицирован ряд других механизмов (27), объясняющих неожиданную сложность этиологии заболевания. Большинство из этих механизмов в настоящее время нацелены на фармакологическое вмешательство, и в будущем появятся и другие, как показано на рис. 2. Именно из-за такой сложности использование одного лекарства вряд ли обеспечит устойчивый эффект, в то время как при постановке диагноза учитывается более одного механизма. можно ожидать, что это приведет к более устойчивой эффективности, особенно если будет проводиться рациональная комбинация.
Рисунок 2Органы и механизмы, участвующие в патогенезе СД2, и целенаправленное лечение. Серым цветом: препараты, находящиеся в разработке. Печатается с разрешения DeFronzo et al. (27). GK, глюкокиназа; GPR, рецепторы, связанные с G-белком; HGP, производство глюкозы в печени; ИЛ, интерлейкин; -I, ингибиторы; PPAR, рецепторы, активируемые пролифератором пероксисом; РА, агонисты рецепторов.
Метформин — сенсибилизатор инсулина с преобладающим действием по ограничению выработки глюкозы в печени.Метформин также увеличивает экспрессию гена GLP-1 в L-клетках и усиливает ответ β-клеток на GLP-1 (28). Следовательно, метформин является «естественным» спутником ингибиторов DPP-4, которые предотвращают деградацию эндогенного GLP-1, улучшают чувствительность β-клеток и увеличивают доступность GLP-1 внутри островка. Ряд испытаний доказали эффективность и низкую частоту побочных эффектов этой комбинации у пациентов, ранее не принимавших лекарственные препараты, хотя еще предстоит проверить, приводит ли это к стойкому эффекту. Испытание VERIFY (эффективность вилдаглиптина в сочетании с метформином для раннего лечения сахарного диабета 2 типа) представляет собой 5-летнее многонациональное двойное слепое исследование с параллельными группами, в котором изучаются долгосрочные эффекты начальной комбинации метформина и вилдаглиптина в последнее время. диагностированных пациентов с легкой гипергликемией, предназначенных для оценки устойчивости гликемического контроля (HbA 1c ), изменений функции β-клеток и чувствительности к инсулину, а также времени до начала введения инсулина (29).
Глитазоны обладают большей долговечностью, чем сульфонилмочевины и метформин (30), и доклинические исследования показали потенциальную защиту функции β-клеток. Первоначальная комбинированная терапия ингибиторами ДПП-4 и пиоглитазоном привела к улучшению некоторых маркеров функции β-клеток, что позволяет предположить, что при продолжении эта комбинация может быть полезной для сохранения функции β-клеток при СД2.
Раннее использование ингибиторов SGLT2 может снизить токсичность глюкозы и, в свою очередь, улучшить функцию β-клеток и чувствительность к инсулину.Однако их использование связано с одновременным повышением уровня глюкагона и выработки глюкозы в печени (31,32), что может сдерживать некоторый терапевтический эффект. Одновременный прием метформина может предотвратить увеличение продукции глюкозы в печени (33), тогда как ингибитор DPP-4 может предотвратить повышение уровня глюкагона (34). Комбинация двух ингибиторов продемонстрировала больший, хотя и не аддитивный эффект по сравнению с использованием отдельных компонентов в снижении HbA 1c , при этом почти вдвое больше пациентов достигли целевого уровня HbA 1c <7% и не увеличили в побочных эффектах (35).
Несмотря на потенциально рациональную комбинацию различных средств, снижающих уровень глюкозы, выбор наиболее подходящей комбинации остается сложной задачей. Некоторая информация будет получена из GRADE (26), хотя в исследование не будут включены недавние препараты, такие как ингибиторы SGLT2. ДеФронцо и его коллеги (36) предложили другой подход. Поскольку развитие и прогрессирование гипергликемии требует сочетания печеночной и периферической инсулинорезистентности и нарушения секреции инсулина, у пациентов с впервые диагностированным СД2 была начата тройная терапия метформином, пиоглитазоном и эксенатидом.За 2 года наблюдения комбинированная терапия по сравнению с классической последовательной терапией метформином, сульфонилмочевиной и инсулином была более эффективной (HbA 1c 5,95 против 6,50%; P <0,001) и имела 7,5 в несколько раз ниже частота гипогликемии и средняя потеря веса 1,2 кг по сравнению со средней прибавкой в весе 4,1 кг ( P <0,01) с последовательной дополнительной терапией (36). Однако этот подход предполагает, что все пациенты имеют один и тот же патогенетический профиль. Хотя это разумно, это не может быть универсально приемлемым, что предполагает необходимость индивидуального патогенетического профилирования.Прямая оценка патогенетического механизма теоретически возможна, но нереальна в клинических условиях. Недавно были идентифицированы биомаркеры, которые связаны с чувствительностью к инсулину и риском развития диабета (37), хотя все еще существует неудовлетворенная потребность в биомаркерах функции и массы β-клеток и других патогенетических компонентов.
Таким образом, фенотип пациента может быть сложным, и, хотя некоторые черты фенотипа могут быть легко доступны, их интерпретация может по-прежнему зависеть от личных знаний и опыта врача.Может помочь разработка специального алгоритма (38), но для проверки может потребоваться большая работа. Недавно мы предложили алгоритм (39), основанный на результатах проведенного опроса, в ходе которого 244 ведущих диабетолога из всех регионов мира представили ряд примеров болезни. Затем алгоритм был использован для определения индивидуальных гликемических целей и подтвержден путем представления трех новых случаев 57 ведущим диабетологам, которые предложили гликемические цели, аналогичные тем, которые были рассчитаны с помощью алгоритма. Мы пришли к выводу, что предлагаемый алгоритм может быть дополнительным инструментом принятия решений, предлагаемым клиницисту, чтобы дополнить принятие клинических решений при рассмотрении целевого гликемического показателя для отдельного пациента с диабетом.Доступных данных об индивидуальных реакциях на лекарства по-прежнему недостаточно, чтобы их можно было встроить в эффективный алгоритм, а описание фенотипа может оставаться слишком широким, чтобы предоставить достаточную информацию о лежащих в основе патогенных факторах. Более того, могут потребоваться и другие факторы, такие как ранний ответ на терапию, снижающую уровень глюкозы, как недавно было предложено Fu et al. (40). Совсем недавно разработка более доступных методов генотипирования и открытие ряда генетических вариантов, связанных с риском СД2, повысили ожидания более точного индивидуального лечения.Насколько нам известно, не было полностью исследовано, может ли сочетание фенотипирования и генотипирования предоставить лучшие возможности. В нескольких исследованиях, в которых это предпринималось (41–43), наблюдалось лишь небольшое улучшение способности предсказывать будущее развитие СД2.
Персонализированная терапия по генотипу
СД2 распознает сложный патогенез, и каждый механизм распознает генетический фон. Многие надеются, что с помощью генотипирования можно будет создать точную медицину.Фармакогенетика все еще молода, проведено очень мало исследований, большинство из которых имеют небольшой размер выборки и разный дизайн. Тем не менее, несмотря на текущие ограничения, существуют примеры управляемого выбора лечения диабета (Таблица 1).
Таблица 1Фармакогенетические эффекты лекарств у пациентов с диабетом, несущих зарегистрированные генетические варианты
Уроки моногенных форм диабета
MODY (диабет зрелого возраста у молодых) — это моногенная форма диабета с аутосомно-доминантным наследованием.MODY характеризуется дефектами секреции инсулина, и существует не менее шести, а, скорее всего, больше различных типов MODY. MODY предлагает хороший пример фармакогенетических рекомендаций. MODY 3, наиболее распространенный тип MODY (~ 61%), возникает из-за мутаций в гене HNF1A (ядерный фактор 1-α печени) и часто ошибочно диагностируется как диабет 1 типа, требующий лечения инсулином. Однако эти пациенты очень чувствительны к сульфонилмочевине, поэтому требуется одна четверть или меньше дозы, обычно назначаемой пациентам с СД2 (44).Эта гиперчувствительность возникает из-за снижения экспрессии HNF1A , вызывая снижение поглощения печенью сульфонилмочевины и повышение ее уровней в крови. MODY 2 (~ 15%) вызывается мутациями в гене GCK (глюкокиназа) и характеризуется легкой гипергликемией, обычно не требующей лечения. Мутация приводит к снижению активности глюкокиназы поджелудочной железы, что приводит к увеличению порога чувствительности к глюкозе (от 6 до 8 ммоль / л) для стимуляции секреции инсулина. Диагностика MODY 2 имеет важное значение для лечения женщин с гестационным сахарным диабетом.Если плод несет ту же мутацию, лечение матери инсулином может снизить уровень глюкозы до уровня, недостаточного для поддержания секреции инсулина плода, что приведет к задержке внутриутробного развития (45).
Другим примером генетики, обеспечивающей руководство по лечению, является неонатальный диабет (NDM), вызванный активирующими мутациями в генах KCNJ11 и ABCC8 . Эти гены кодируют две субъединицы каналов K ATP рецептора сульфонилмочевины в β-клетках, вызывая неспособность закрыть каналы K ATP и нарушенную секрецию инсулина.Пациенты, несущие мутации KCNJ11 / ABCC8 , плохо регулируются инсулином, но очень хорошо реагируют на высокие дозы сульфонилмочевины (46). У детей с НСД часто встречается синдром DEND (задержка развития, эпилепсия и неонатальный диабет). Поскольку ген KCNJ11 также экспрессируется в головном мозге, перевод детей на высокие дозы сульфонилмочевины также может положительно повлиять на дефекты развития (46).
Обычный T2DM
Появление технологий генотипирования и секвенирования способствовало открытию многих генетических вариантов, вносящих вклад в патогенетическую сложность T2DM.Эти достижения открывают привлекательную возможность определить полезность участков генома для прогнозирования ответа на лечение. Однако интерпретация фармакогенетических данных не всегда ясна. В то время как в большинстве исследований в качестве основного результата используется улучшение уровней HbA 1c , другие определяют респондентов с точки зрения процента снижения или снижения уровня HbA 1c глюкозы натощак и после приема пищи и, в некоторых случаях, изменений секреции инсулина и чувствительности. Эти различия могут способствовать несогласованности и неоднородности результатов.В следующем разделе будут кратко рассмотрены фармакогеномические исследования с различными сахароснижающими агентами.
Метформин
Ряд генетических вариантов был связан с различными ответами на метформин. Препарат представляет собой органический катион, который не метаболизируется в организме и требует переносчиков для абсорбции, распределения, метаболизма и выведения. Варианты органического переносчика катионов, OCT1 ( SLC22A1 ), были связаны в некоторых (47), но не во всех (48) исследованиях с более низкой эффективностью метформина и повышенным почечным клиренсом.Точно так же лучшая эффективность и снижение риска развития диабета были связаны с генетическими вариантами MATE1 и MATE2 (белки экструзии с множеством лекарственных средств 1 и 2) (49). Наконец, в результате общегеномной ассоциации варианты ATM (мутация телеангиэктазии атаксии) были связаны со значительным снижением уровней HbA 1c после лечения метформином (50).
Sulfonylureas
Фармакогенетические исследования сульфонилмочевины были сосредоточены на генетических вариантах, влияющих на каналы β-клеток поджелудочной железы K ATP и фермент, метаболизирующий лекарственные средства, CYP2C9.Было обнаружено, что варианты в генах KCNJ11 и ABCC8 , кодирующие субъединицы канала АТФ K , связаны с риском СД2 и снижением ответа на сульфонилмочевину (51). Кроме того, было обнаружено, что два варианта гена CYP2C9 ( CYP2C9 * 2 и CYP2C9 * 3) связаны с повышенными уровнями лекарств и повышенным риском гипогликемии после лечения сульфонилмочевиной (52), что соответствует лучшему HbA 1c ответ в исследовании Genetics of Diabetes Audit Research в Tayside Scotland (GoDARTS) (53).Наконец, в GoDARTS сообщалось, что люди, несущие вариант TCF7L2 , имеют меньшую реакцию на сульфонилмочевину по сравнению с таковой у лиц, не являющихся носителями (54).
Тиазолидиндионы
Тиазолидиндионы (TZD) являются привлекательными кандидатами для фармакогенетических исследований, поскольку варианты гена-мишени лекарственного средства PPARG связаны с риском СД2 и чувствительностью к инсулину в популяциях без диабета. Тем не менее, фармакогенетические исследования у субъектов с предиабетом и у субъектов с диабетом дали противоречивые результаты в отношении ответа на терапию TZD (55,56).Генетические варианты фермента CYP2C8 * 3 были связаны со сниженной активностью TZD (57). Наконец, генетические варианты в генах аквапорина 2 ( AQP2 ) и транспортера натрия / калия / хлорида ( SLC12A1 ) были связаны с повышенным риском отека у пациентов, получавших розиглитазон (58), что открывает возможности для фармакогеномики не только в с точки зрения терапевтического ответа, но также и для выявления основных механизмов у субъектов с риском побочных эффектов.
Incretins
Варианты гена химотрипсиногена ( CTRB1 / 2 ) были связаны с нарушением секреции инсулина, стимулированной GLP-1, у 232 человек, перенесших гиперинсулинемический зажим. Варианты также были связаны с повышенной активностью химотрипсиногена и экспрессией генов в островках поджелудочной железы человека (59). Эти результаты нашли поддержку в исследовании GoDARTS, в котором сообщалось о более значительном снижении HbA 1c у носителей варианта CTRB1 / 2 , получавших глиптины (54).Многие из генетических вариантов ( TCF7L2 , WFS1 и KCNQ1 ), повышающие чувствительность к T2DM, связаны с нарушением действия инкретина, тогда как варианты гена рецептора глюкозозависимого инсулинотропного пептида (GIP) ( GIPR ) были связано со снижением секреции инкретина (60). Недавно сообщалось о некоторых различиях в ответе на линаглиптин у пациентов с диабетом с вариантами гена TCF7L2 и без них (61).
Метиглиниды
Метиглиниды (натеглинид и репаглинид) закрывают каналы K ATP , чтобы стимулировать секрецию инсулина посредством взаимодействия с субъединицами, отличными от тех, которые активируются сульфонилмочевиной.Эффекты генетических вариантов в переносчиках ( SLCO1B1 ) и метаболизирующих ферментах ( CYP2C9 ) были связаны со снижением скорости метаболизма глинидов, хотя никакого эффекта в ответе на лечение не наблюдалось (62). Интересно, что вариант импринтированного гена KCNQ1 был связан с лучшими реакциями чувствительности к глюкозе и инсулину после 48-недельной терапии репаглинидом, но не тогда, когда эти эффекты были скорректированы с учетом возраста, пола и ИМТ (63). KCNQ1 ранее был вовлечен в патогенез СД2, проявляя эффекты родительского происхождения, что свидетельствует о важности учета того, наследуются ли аллели от матери или от отца.
Будущие направления
Фармакогенетика обладает потенциалом для открытия новых мишеней для лекарственных средств. Например, недавнее исследование в регионе Ботния в Финляндии, сравнивающее людей с ранним началом СД2 и субъектов, не страдающих диабетом, несмотря на кластеризацию факторов риска СД2, привело к идентификации редкого защитного варианта в гене SLC30A8 (R138X) ( 64). Последующие исследования прольют свет на биологию, лежащую в основе защитного эффекта, и, мы надеемся, облегчат разработку лекарств на основе этих механизмов.
Помимо биологических факторов, реакция на лекарственную терапию также включает взаимодействие генов и окружающей среды, включая режимы питания и физическую активность, а также психосоциальные факторы, такие как мотивация, образование, поддержка семьи и приверженность лечению. Снижение когнитивной функции у пациентов с СД2 может играть роль в приверженности лечению. Было показано, что области генома, связанные с когнитивной функцией, имеют положительный эволюционный отбор, но этот феномен не наблюдался для регионов, связанных с факторами риска поздних метаболических заболеваний (65).Следовательно, будет интересно изучить, в какой степени ответ на терапию и плохое соблюдение режима определяются когнитивной функцией пациента. Более того, индивидуальная генетическая предрасположенность может быть изменена эпигенетическими механизмами, задействованными в различных тканях и органах, включая несколько областей мозга. Наконец, необходимы многоэтнические исследования для изучения обобщения ответов на лечение в различных популяциях. Таким образом, накапливаются доказательства того, что генетика может стать важным инструментом для диагностики, классификации и лечения диабета, как это имеет место в случае моногенных форм диабета.Точно так же фармакогенетика успешно использовалась для выбора терапии при моногенетических формах диабета, хотя до сих пор имела ограниченное клиническое применение при полигенных фенотипах СД2. Однако генетические эффекты могут быть изменены различными факторами окружающей среды с последующими эпигенетическими модификациями. Следовательно, необходимы все более масштабные исследования, чтобы лучше классифицировать гетерогенный СД2 на разные подгруппы, чтобы оценить потенциал фармакогеномики для персонализированного лечения.
Выводы
Точная медицина — это не просто академическая дискуссия, поскольку она уже входит в политическую повестку дня наиболее промышленно развитых стран. Президент Барак Обама в своем послании о положении страны в январе 2015 года объявил о запуске «Инициативы точной медицины, которая приблизит нас к лечению таких болезней, как рак и диабет». Этот сценарий был четко описан в своих задачах, ожиданиях, исследованиях и научных подходах в четкой редакционной статье Коллинза и Вармуса (66), где рак был определен как «ближайший фокус», тогда как «целый ряд болезней» будет быть целевой в более долгосрочной перспективе.Перспективы индивидуализированной терапии для людей с диабетом заключаются в обеспечении эффективного, устойчивого и безопасного лечения. На основе индивидуального — фенотипического и генетического — профиля пациента прецизионная медицина направлена на прогнозирование того, какой пациент с большей вероятностью получит пользу, а какой с большей вероятностью испытает побочные эффекты в ответ на терапевтические методы. Несмотря на то, что был идентифицирован ряд фенотипических признаков и генетических вариантов, их интерпретация и перевод в клинические руководства остаются неопределенными и недостаточно исследованными, и, таким образом, имеют ограниченное, если вообще, клиническое использование.Требуется больше данных и больше клинических испытаний. С этой целью кажется желательным, чтобы данные, полученные в клинических испытаниях новых классов лекарственных средств, можно было объединить, чтобы получить достаточную мощность для определения основных предикторов терапевтического успеха и риска неблагоприятных событий. Такой подход может потребовать от отрасли отказаться от ожиданий «препарата-блокбастера», то есть препарата, который можно использовать для подавляющего большинства людей с СД2. Напротив, было бы очень полезно понять, какое лекарство (а) лучше подходит для данного человека в данной фазе заболевания.Сбор этой информации не может ограничиваться рандомизированными клиническими исследованиями. Скорее, постмаркетинговый крупномасштабный эпиднадзор, электронные реестры здоровья и даже мобильные медицинские технологии могут быть использованы для получения более достоверной информации в течение более длительного периода наблюдения в более крупных и разнообразных группах населения с диабетом. В настоящее время доступно до 13 классов препаратов для лечения СД2, нацеленных на различные патогенетические механизмы. Проблема состоит в том, как использовать правильное лекарство (и правильную комбинацию лекарств) у правильного пациента.Пока мы ждем более научных и менее прагматичных указаний, следует помнить о нескольких вещах. Мы должны понимать, что СД2 — это сложное заболевание со сложным патогенезом и, следовательно, его нелегко лечить, особенно в стремлении к долгосрочному контролю гликемии. С этой точки зрения следует приветствовать множество доступных фармакологических агентов, хотя для их надлежащего использования требуются экспертные знания о преимуществах и недостатках каждого доступного лекарственного средства и возможности рационального комбинирования.Эта сложность может противоречить потребности в эффективном лечении растущего числа людей с СД2 во всем мире и необходимости разработки стратегий лечения, которые могут быть реализованы в первичной медико-санитарной помощи. Тем не менее, неправильное использование лекарств может не дать необходимой реакции и, особенно в случае новых агентов, может привести к неоправданному завышению затрат. Поэтому обязанностью диабетического сообщества является поиск новой стратегической модели взаимодействия между специалистом и другими поставщиками медицинских услуг, обеспечивающей надлежащее управление индивидуальным лечением человека с диабетом.
Генотип против фенотипа | Биологический словарь
Генетический состав организма называется его генотипом, а наблюдаемые черты — его фенотипом. Другими словами, то, как человек выглядит, является результатом его или ее ДНК. Прекрасным примером, объясняющим эту концепцию, являются эксперименты Грегора Менделя с чистыми растениями желтого и зеленого гороха (P 1 ). Фенотип растений — это их цвет, желтый и зеленый. Когда Мендель скрестил эти два растения, все потомство (F 1 ) было желтым.Но зеленый цвет не был потерян, так как он снова появился в следующем поколении (F 2 ). В то время Мендель не знал этого, но лежащие в основе генотипы зеленых и желтых растений были ответственны за черты, наблюдаемые в поколении F 1 .
И зеленые, и желтые растения гороха P 1 имели идентичные аллели гена, контролирующего их цвет, то есть они были гомозиготными. Когда два зеленых или два желтых растения были скрещены вместе, все потомки зеленых растений были зелеными, а все потомки желтых растений были желтыми.Однако, когда зеленое растение было скрещено с желтым, все потомки F 1 были желтыми. Это связано с тем, что поколение F 1 было гетерозиготным по цветному признаку, то есть у них были разные аллели по цвету. И из двух аллелей гена окраски желтый был доминирующим, делая все растения F 1 желтыми. Аллели растения гороха в F 2 должны быть гомозиготными рецессивными, чтобы растение было зеленым
Сравнительная таблица
| Генотип | Фенотип | |
|---|---|---|
| Наблюдаемые признаки организма? | Нет | Да |
| Генетический состав организма? | Да | Нет |
| Как это наблюдается? | Секвенирование ДНК, анализ ПЦР, если необходимо (см. Изображение ниже) | Посмотрев на организм |
| Относится к информации, закодированной по аллелям | Да | Нет |
| Примеры включают цвет волос, цвет глаз , высота | Нет | Да |
| По наследству? | Частично, по одному аллелю от каждого родителя | Нет |
| Что он содержит (показать)? | Вся информация о наследственности для организма, даже о невыраженных генах. | Только экспрессируемые гены. |
На изображении выше показано, как работает метод полимеразной цепной реакции (ПЦР) для амплификации небольших образцов ДНК для генетического анализа. Генетический анализ выявляет генотип человека, который кодирует наблюдаемые характеристики, то есть фенотип.
Ссылки
- Генотип против фенотипа. (нет данных). В Diffen.com . Получено с https://www.diffen.com/difference/Genotype_vs_Phenotype
- OpenStax.(20 мая 2013 г.). Эксперименты Менделя в наследственности. В Биология . (Глава 12). Получено с http://cnx.org/content/col11448/latest/
Подпишитесь на нашу рассылку новостей
Генотип против фенотипа [видео]
Генотип против фенотипа
Привет, и добро пожаловать в этот обзор генотипов и фенотипы! Что такое генотип? Как связаны генотипы и фенотипы? Как мы можем применить эти термины в реальных жизненных ситуациях? В этом видео мы ответим на все эти вопросы.Мы сравним и сопоставим генотипы и фенотипы, чтобы понять, какой цели они служат в мире генетики.
Прежде чем мы сможем полностью понять генотипы и фенотипы, мы сначала должны понять аллеля .
Гены встречаются в определенных местах на каждой хромосоме и состоят из определенной химической последовательности оснований аденина, гуанина, цитозина и тимина. Однако последовательность гена на одной копии хромосомы может немного отличаться от последовательности на другой копии хромосомы.Если это так, то этот вариант последовательности для того же гена является нашим аллелем. Аллель — это вариант гена на хромосоме, который помогает определить наследственность организма. Поскольку у нас есть две копии каждого гена (одна от мамы и одна от папы), это означает, что все соматические клетки имеют по два аллеля для каждого признака. Вместе эти два аллеля определяют наследуемый вами признак.
Возьмем, к примеру, высоту. Несмотря на то, что на рост влияет несколько факторов, мы будем упрощать и говорить, что кто-то может быть высоким или низким.Допустим, ген, который кодирует рост на одной хромосоме, имеет последовательность, которая читается следующим образом: A C GTC. Скажем, этот аллель кодирует слово «высокий». Ген, который кодирует рост на другой хромосоме, имеет последовательность, которая гласит: A G GTC. Этот аллель кодирует «короткое». Опять же, поскольку ген немного отличается (одна последовательность имеет цитозин в качестве второго основания, а другая — гуанин), это считается аллелем. Мы можем присвоить нашим аллелям произвольные заглавные и строчные буквы, чтобы упорядочить их.Давайте не будем усложнять и возьмем заглавную T для «высокого» и строчную t для «короткого», например:
| Аллель | Последовательность | Фенотип |
| T | A C GTC | высокий |
| t | A G GTC | короткий |
Таким образом, мы можем иметь один и тот же ген, кодирующий один и тот же признак на обеих хромосомах, но вариант последовательности или аллель — это то, что дает разные фенотипы.Следовательно, фенотип — это физический признак, который определяется имеющимися аллелями. Фенотипы помогают нам наблюдать и анализировать генетические мутации, особенно когда мы думаем об эксперименте по отслеживанию наследования признака. Если мы хотим знать, проявится ли признак кускового крыла у потомства самца и самки плодовой мухи, все, что нам нужно сделать, это поискать этот признак у их потомства.
Иногда просто увидеть фенотип недостаточно. Может быть, мы хотим знать точное сочетание аллелей, которое вызвало эту черту.Здесь на помощь приходит генотип. Генотип — это пара аллелей, унаследованная от определенного гена, который производит определенный фенотип. На самом деле генотип относится ко всем генам в организме, а фенотип относится ко всем наблюдаемым признакам организма, но на практике они обычно используются для обозначения одного рассматриваемого гена.
Теперь давайте соберем все вместе и посмотрим, как генотипы и фенотипы на самом деле влияют на наследование. Помните, что быть диплоидным организмом означает, что вам нужно бороться не с одной копией аллеля, а с двумя! Различные комбинации этих двух аллелей — вот что приводит к наблюдаемому фенотипу или признаку.Человек с двумя копиями одного и того же аллеля считается гомозиготным по этому признаку, а человек с одной копией одного аллеля и другой копией другого аллеля считается гетерозиготным по этому признаку. Итак, возвращаясь к нашему примеру роста, человек был бы гомозиготным по росту, если бы у него был генотип TT или генотип tt. Следовательно, индивид с генотипом Tt гетерозиготен. «TT» означает то же самое, что и «Tt», поэтому здесь есть только один вариант.
Это важно, потому что то, является ли человек гомозиготным или гетерозиготным по признаку, может означать разницу между передачей унаследованной мутации или нет.Некоторые аллели были идентифицированы как мутантные аллели, то есть они содержат мутацию, которая может вызвать изменение или заболевание. Если аллель не является мутантным аллелем, он называется аллелем дикого типа. Этот тип аллеля считается «нормальным». Итак, как мы узнаем, вызовет ли мутантный аллель изменение или нет? Это зависит от того, как он сочетается со вторым аллелем этого гена. Другими словами, это зависит от того, являются ли аллели доминантными или рецессивными.
Мутантные аллели могут быть доминантными, что означает, что они перекрывают другой аллель, или рецессивными, что означает, что они не перекрывают другой аллель.Доминантные аллели обычно обозначаются заглавной буквой, а рецессивные аллели — строчной. Давайте посмотрим на это более подробно, используя мутацию ямочки. Допустим, доминантный аллель означает, что у вас есть ямочки на щеках, поэтому мы присвоим ему заглавную букву «D». Предположим, рецессивный аллель означает, что у вас нет ямочек. Мы присвоим ему строчную букву «d». Если мы нарисуем простой квадрат Пеннета, мы сможем определить генотипическое и фенотипическое соотношение для ямок. В этом примере у нас есть мама и папа, которые гетерозиготны по ямочкам с генотипом «Dd».
Если мы заполним квадрат Пеннета, перетаскивая каждый аллель вниз и поперек для каждого поля, это будет наш результат. Начнем с соотношения генотипов. У нас есть одно потомство с гомозиготным доминантным «DD», два потомка с гетерозиготным «Dd» и одно гомозиготное рецессивное «dd», поэтому соотношение составляет 1: 2: 1. Поскольку доминантным аллелем является «D», все потомки с генотипами DD и Dd будут иметь ямочки, потому что аллель «D» преобладает над рецессивным аллелем. У нас также есть одно гомозиготное рецессивное потомство, у которого нет доминантного аллеля, поэтому эта комбинация не даст ямок на щеках.При этом мы можем сказать, что фенотипическое соотношение составляет 3: 1, ямочки на щеках отсутствуют. Мы также можем сказать, что 75% потомков будут иметь ямочки, а 25% — нет. Этот пример показывает, как генотип и фенотип, будучи связанными, могут дать нам различную, но одинаково важную информацию о наследовании.
Вот и все! Давайте кратко рассмотрим вопрос, чтобы проверить свои знания.
Что из этого правильно описывает основное различие между генотипами и фенотипами?
- Генотипы — это кодирующие гены, а фенотипы — некодирующие гены
- Генотипы — это рецессивные аллели, а фенотипы — доминантные аллели
- Генотипы — это генетическая информация, а фенотипы — информация об окружающей среде
- Генотип — это совокупность генов организма, а фенотип — это его физические характеристики
Правильный ответ — D! Генотип организма — это все его гены, вплоть до конкретной аллельной последовательности, которую они несут, а фенотип — это все физические характеристики организма.