Непосредственное взаимодействие — Справочник химика 21
Водород проявляет и восстановительные, и окислительные свойства. В обычных условиях благодаря прочности молекул он сравнительно мало активен и непосредственно взаимодействует лишь со фтором. При нагревании же вступает во взаимодействие с многими неметаллами — хлором, бромом, кислородом и пр. Восстановительная способность водорода используется для получения некоторых простых веществ из оксидов и галидов [c.274]Аргон образует молекулярные соединения включения — клатраты— с водой, фенолом, толуолом и другими веществами. Гидрат аргона примерного состава Аг 6Н гО представляет собой кристаллическое вещество, разлагающееся при атмосферном давлении при —42,8 С. Его можно получить непосредственным взаимодействием аргона с водой при 0°С и давлении порядка 1,5 10 Па. С соединениями НаЗ, 502, СОг, ПС1 аргон дает двойные гидраты, т.

При изучении коррозионных свойств нефтепродуктов необходимо рассматривать две разные системы нефтепродукт + металл и нефтепродукт + вода + металл. В первом случае скорость коррозии металлов будет определяться наличием в нефтепродуктах коррозионно-агрессивных веществ и их способностью непосредственно взаимодействовать с металлами (химическая коррозия). Во втором случае корозия металлов в нефтепродуктах должна развиваться преимущественно по электрохимическому механизму. [c.282]
Инструментом называют элемент, с которым непосредственно взаимодействует изделие (фреза, а не станок огонь, а не горелка). В частности, инструментом может быть часть окружающей среды. Инструментом являются и стандартные детали, из которых собирают изделие. Например, набор частей игры Конструктор — это инструмент для изготовления различных моделей.
Полярные протонные растворители легко сольватируют как анионы, так и катионы. Неорганические катионы взаимодействуют со свободными электронными парами, тогда как анионы сольватируются путем образования водородных связей. Крупные четвертичные аммониевые ионы не сольватируются [37] или по крайней мере сольватируются не специфично, т. е. сильного непосредственного взаимодействия с растворителем не существует. В этих растворителях имеет место высокая степень диссоциации на свободные сольватированные ионы. Однако многие анионы обладают относительно низкой реакционноспособностью (нуклеофильностью) из-за сильного экранирования сольватной оболочкой.
Неорганические катионы взаимодействуют со свободными электронными парами, тогда как анионы сольватируются путем образования водородных связей. Крупные четвертичные аммониевые ионы не сольватируются [37] или по крайней мере сольватируются не специфично, т. е. сильного непосредственного взаимодействия с растворителем не существует. В этих растворителях имеет место высокая степень диссоциации на свободные сольватированные ионы. Однако многие анионы обладают относительно низкой реакционноспособностью (нуклеофильностью) из-за сильного экранирования сольватной оболочкой.
Первый член описывает расщепление в нулевом поле, следующие два члена—влияние магнитного поля на спиновую мультиплетность, остающуюся после расщепления в нулевом поле члены с Ац и являются мерой сверхтонкого расщепления параллельно и перпендикулярно главной оси, а Q —мерой небольших изменений в спектре, вызванных ядерным квадрупольным взаимодействием. Все эти эффекты обсуждались в гл. 9. Последний член учитывает тот факт, что ядерный магнитный момент может непосредственно взаимодействовать с внешним полем Яд = Нц /, где у — гиромагнитное отношение ядра, а Р — ядерный магнетон Бора.
Тетрафторид углерода можно получить непосредственным взаимодействием простых веществ или ио обменной реакции между ССЦ и AgP при 300°С. Тетрахлорид получают хлорированием Sj в присутствии катализатора [c.400]
Известно иного соединений серы с галогенами. Они получаются при непосредственном взаимодействии простых веществ. Сера реагирует со всеми галогенами, кроме иода. [c.454]
Из них наиболее реакционноспособны меркаптаны, особенно ароматические (тиофенолы). Меркаптаны могут служить инициаторами окисления углеводородов, вызывая зарождение окислительных цепей. Продукты окисления меркаптанов склонны к конденсации и уплотнению. Ввиду высокой реакционной способности меркаптаны даже в условиях мягкого окисления образуют сульфоновые кислоты, вызывающие сильную коррозию металлов [67, 68]. Меркаптаны способны, кроме того, непосредственно взаимодействовать с металлами, образуя меркаптиды это приводит к накоплению в топливе нерастворимых продуктов коррозии. Сульфиды и тиофены гораздо менее активны, чем меркаптаны. Превращения (окисление) таких соединений наблюдаются только при повышенных температурах. При этом в основном образуются продукты уплотнения, которые дают в топливах нерастворимые осадки. Встречающиеся в топливах в незначительном количестве дисульфиды более активны, чем сульфиды. Они при окислении образуют смолистые вещества.
Меркаптаны могут служить инициаторами окисления углеводородов, вызывая зарождение окислительных цепей. Продукты окисления меркаптанов склонны к конденсации и уплотнению. Ввиду высокой реакционной способности меркаптаны даже в условиях мягкого окисления образуют сульфоновые кислоты, вызывающие сильную коррозию металлов [67, 68]. Меркаптаны способны, кроме того, непосредственно взаимодействовать с металлами, образуя меркаптиды это приводит к накоплению в топливе нерастворимых продуктов коррозии. Сульфиды и тиофены гораздо менее активны, чем меркаптаны. Превращения (окисление) таких соединений наблюдаются только при повышенных температурах. При этом в основном образуются продукты уплотнения, которые дают в топливах нерастворимые осадки. Встречающиеся в топливах в незначительном количестве дисульфиды более активны, чем сульфиды. Они при окислении образуют смолистые вещества.
В превращениях, происходящих в результате непосредственного взаимодействия между твердыми фазами, химическая реакция протекает на поверхности соприкосновения обеих фаз. Следовательно, факторы, влияющие на увеличение реакционной поверхности (измельчение твердых тел или спрессовывание раздробленных реагентов), вызывают быстрое достижение полного протекания реакции. Однако по мере прохождения реакции на поверхности соприкосновения фаз возрастает слой образующегося продукта и медленная диффузия исходного вещества через этот слой к месту реакции лимитирует скорость превращения. В этих условиях повышение степени измельчения фаз также обеспечивает ускорение превращения вследствие того, что уменьшается путь диффузии в твердой фазе.
Следовательно, факторы, влияющие на увеличение реакционной поверхности (измельчение твердых тел или спрессовывание раздробленных реагентов), вызывают быстрое достижение полного протекания реакции. Однако по мере прохождения реакции на поверхности соприкосновения фаз возрастает слой образующегося продукта и медленная диффузия исходного вещества через этот слой к месту реакции лимитирует скорость превращения. В этих условиях повышение степени измельчения фаз также обеспечивает ускорение превращения вследствие того, что уменьшается путь диффузии в твердой фазе.
Тензор напряжений в дисперсной фазе Хд можно полагать равным нулю лишь в том случае, если частицы взаимодействуют между собой только через посредство сплошной фазы, а непосредственные взаимодействие частиц (столкновения) отсутствуют. Считается, что такая ситуация имеет место при движениях твердых частиц, капель или пузырей в жидкостях. При движении твердых частиц в газе и больших объемных содержаниях твердой фазы через поверхность вьщеленного объема смеси посредством столкновений частиц передается дополнительный импульс, связанный с их хаотическим движением.
Предельно-необратимый процесс, протекающий при постоянном давлении и полном отсутствии полезной работы А (непосредственное взаимодействие серебра и хлора) [c.528]
Обосновать невозможность получення оксидов хлора непосредственным взаимодействием хлора G кислородом.
Какими свойствами обладает азидоводород (азотистоводородная кислота) и его соли Можно лн получить HN3 непосредственным взаимодействием азота и водорода Ответ мотивировать. [c.229]
При добавлении щелочи происходит непосредственное взаимодействие катионов с ионами гидроксила [c.79]
Соединения. Хлор, бром, иод непосредственно взаимодействуют с водородом, образуя галогенводороды [c.475]
Рис, 1,8. Химико-технологическая система периодического действия с непосредственным взаимодействием аппаратов (а) и временные графики ее функционирования [c.
Формулы (1.4), (1.5) справедливы для систем последовательной структуры при непосредственном взаимодействии аппаратурных стадий. При наличии между стадиями промежуточных емкостей система функционирует в квазинепрерывном режиме, и ее технологический цикл определяется продолжительностью цикла последнего аппарата, деленной на число параллельных аппаратов, образующих эту стадию [c.34]
Бинарные соединения брома (I) и иода (I), кроме ВГ2О, образуются при непосредственном взаимодействии простых веществ. За исключением I (AWjgf = —12 кДж/моль), они очень неустойчивы. [c.303]
В обычных условиях азот непосредственно взаимодействует лишь с литием с образованием LigN. При активации молекул N2 (нагре-ваниш, действием электроразряда или ионизирующих излучений) азот )бычно выступает как окислитель и лишь при взаимодействии с фтором и кислородом — как восстановитель.
Хлорид ртути 1), или сулема, Hg i2 может быть получен непосредственным взаимодействием ртути с хлором. Это бесцветное вещество, сравнительно мало растворимое в холодной воде (6,6 г в 100 г воды при 20 С). Однако с повышением температуры [c.627]
Это бесцветное вещество, сравнительно мало растворимое в холодной воде (6,6 г в 100 г воды при 20 С). Однако с повышением температуры [c.627]
Сульфгды образуются непосредственным взаимодействием простых вещести или действием сероводорода на растворимые соединения Э(1П) в кнелей среде
Регенерация реагентов. Часто в систему необходимо вводить вспомогательные исходные вещества, например, когда новый ход процесса будет более выгодным, чем при непосредственном взаимодействии основных исходных веществ, или даже единственно возможным. В этом случае нужно так организовать производственный цикл, чтобы вспомогательное исходное вещество можно было регенерировать. После регенерации это вещество возвращается в цикл, и его расход ограничивается только потерями. Такой метод широко используется в химической технологии. Отметим, что он отличается от рециркуляции реагента, олисанной на стр. 356. Обычно возвращаемое в цикл вспомогательное йсходное вещество регенерируется в результате химического превращения, а не выделяется из смеси физическими методами. Примером может служить использование концентрированной гидроокиси натрия для разложения боксита в производстве окиси алюминия методом Байера, сохранение в цикле окислов азота при башенном способе получения серной кислоты или введение в цикл аммиака при производстве соды методом Сольвея. В последнем случае процесс не может проводиться при, непосредственном взаимодействии основных исходных веществ по уравнению [c.377]
Примером может служить использование концентрированной гидроокиси натрия для разложения боксита в производстве окиси алюминия методом Байера, сохранение в цикле окислов азота при башенном способе получения серной кислоты или введение в цикл аммиака при производстве соды методом Сольвея. В последнем случае процесс не может проводиться при, непосредственном взаимодействии основных исходных веществ по уравнению [c.377]
При обычной температуре элементарный углерод весьма инертен. При высоких же температурах он непосредственно взаимодействует с многими металлами и неметаллами. Углерод проявляет восстановительные свойства, что широко используется в металлургии. Окислительные свойства углерода выражены слабо. Вследствие различия в структуре алмаз, графит и карбин по-разному ведут себя в химических реакциях. Для графита характерны реакции образования кристаллических соединений, в которых макромолекулярные слои С200 играют роль самостоятельных радикалов. [c.394]
Г а. л и д ы SiHalj можно получить непосредственным взаимодействием простых веществ. Тетрафторид обычно получают действием концентрированной серной кислоты на смесь SiOa + aF [c.413]
л и д ы SiHalj можно получить непосредственным взаимодействием простых веществ. Тетрафторид обычно получают действием концентрированной серной кислоты на смесь SiOa + aF [c.413]
Можно ли получить Н2О2 непосредственным взаимодействием водорода и кислорода Ответ обосновать. [c.220]
В обычных условиях бор (подобно кремнию) весьма инертен и непосредственно взаимодействует только со фтором при нагревании (400—700 С) окисляется кислородом, серой, хлором и даже азотом (выше 1200″С). С водородом бор не взаимодействует. При сильном нагревании восстановительная активность бора проявляется и в отно-HjeHHH таких устойчивых оксидов, как Si02, Р2О5 и др. [c.436]
С i л ь ф и д алюминия А1,83 получают непосредственным взаимодействием простых веществ при темпе 1атуре красного каления. Это кристаллическое вещество (т. пл. 1()0″С), полностью гидролизующееся да>1 е следами влаги в воздухе. При сплавлении АЬЗз с основными сульф 1Дами образуются полимерные сульфидоалюминаты состава [c. 457]
457]
Карбиды и нитриды подгруппы титана образуются непосредственным взаимодействием простых веществ при высокой температуре. Соединения 3N и ЭС (переменного состава) — кристаллические вещества, очень твердые, тугоплавкие (3000—4000° С), хорошо проводят электрический ток и химически инертны. Аналогичными свойствами обладают силициды 3Si 2, бориды ЭВ, ЭВг, Все они, конечно, обладают переменным составом. Соединения Ti , TiN, TiO, ZrN, Zr , Hf имеют структуру типа Na l 11 друг с другом образуют твердые растворы. [c.532]
С кислородом непосредственно взаимодействует только медь. При температуре красного каления образуется СиО, а при более высокой температуре ujO с серой непосредственно взаимодействуют Си и Ag [c.622]
Авторы работы [41] предположили, что обмен протекает с участием фенильных радикалов, а реакция присоединения идет при непосредственном взаимодействии молекул и.) (или двух атомов В) с бензолом. Интересно отметить, что экспериментальЕше данные хорошо согласуются с предположением о том, что Вз и Сс,Нв адсорбируются на участках поверхности без конкуренции и лимитирующей стадией обмена является реакция РЬ-Н-З — 8 РЬ-8- -Н-8. Аналогичный механизм в случае этилена хорошо объясняет реакцию присоединения (но не обмена). [c.550]
Аналогичный механизм в случае этилена хорошо объясняет реакцию присоединения (но не обмена). [c.550]
В таблицу не помещена реакция между твердыми фазами, которая во многих случаях проходит с участием жидкой или газовой фаз, образующихся во время реакции. При ояисании таких превращений необходимо учитывать химическую реакцию, диффузию возникновение газовой фазы в результате диссоциации или йена рения, появление жидкой фазы (плавление) и кристаллизацию Поскольку участие жидкой и газовой фаз в реакциях между твер дыми фазами имеет, как обнаружилось, основное значение для вы яснения их механизма, эти реакции можно подразделить на четыре группы 1) проходящие при непосредственном взаимодействии реагентов в твердой фазе 2) проходящие с участием газовой фазы [c.243]
Как и при любых других самопроизвольно идущих процессах, реакция, протекающая в гал1.ваническом элементе, сопровождается уменьшением энергии Гиббса. Но это означает, что при непосредственном взаимодействии реагирующих веществ реакция будет протекать в том же направлении. Таким образом, сопоставляя электродные потенциалы соответствующих систем, можно заранее определять направление, в котором будет протекать окислительно-восстановительная реакция. [c.185]
Таким образом, сопоставляя электродные потенциалы соответствующих систем, можно заранее определять направление, в котором будет протекать окислительно-восстановительная реакция. [c.185]
В отличне от водородных соединений неметаллов VI и УИ груп[(, аммиак не обладает кислогнымн свойствами. Однако атомы водорода в его молекуле могут замещаться атомами металлов. При полном замещении водорода металлом образуются соединения, называемые нитридами. Некоторые из них, например нитриды кальция и магния, получаются ири непосредственном взаимодействии азога с металлами при высокой температуре [c.400]
Результаты последних исследований [32, 28] позволяют с уве-ренностыр сказать, что антагонистическое действие сероорганиче- ских соединений является следствием реакций, протекающих в газовой фазе в предпламенных стадиях процесса сгорания. Непосредственное взаимодействие некоторых соединений при низких температурах хранения и применения если и протекает, то очень медленно и в общем антагонистическом эффекте сероорганических соединений имеет второстепенное значение. Снижение детонационной стойкости в предпламенный период можно представить следующим образом. [c.139]
Снижение детонационной стойкости в предпламенный период можно представить следующим образом. [c.139]
Таким образом, герметик размягчается при наличии в топливе первичных продуктов окисления—гидропероксидов. Гидропероксиды могут разрушать герметик непосредственно взаимодействуя с ним, либо реагируя со свободными радикалами, которые появляются при распаде гидроперокеида. Это было установлено в следующих опытах. [c.238]
Если не касаться спорного вопроса о наличии внутриионизированных структур олефина [14], то и в этом случае остаются возражения против предложенного механизма, которые уже упоминались выше (если в реагирующей среде одновременно находятся разнаполярные ионы, в продуктах реакции должны присутствовать продукты их непосредственного взаимодействия). [c.14]
Почему при непосредственном взаимодействии хлора, брома и йода с железом образуются соединения с различным валентным состоянием железа РеС1з, РеВг2 и РеЬ [c.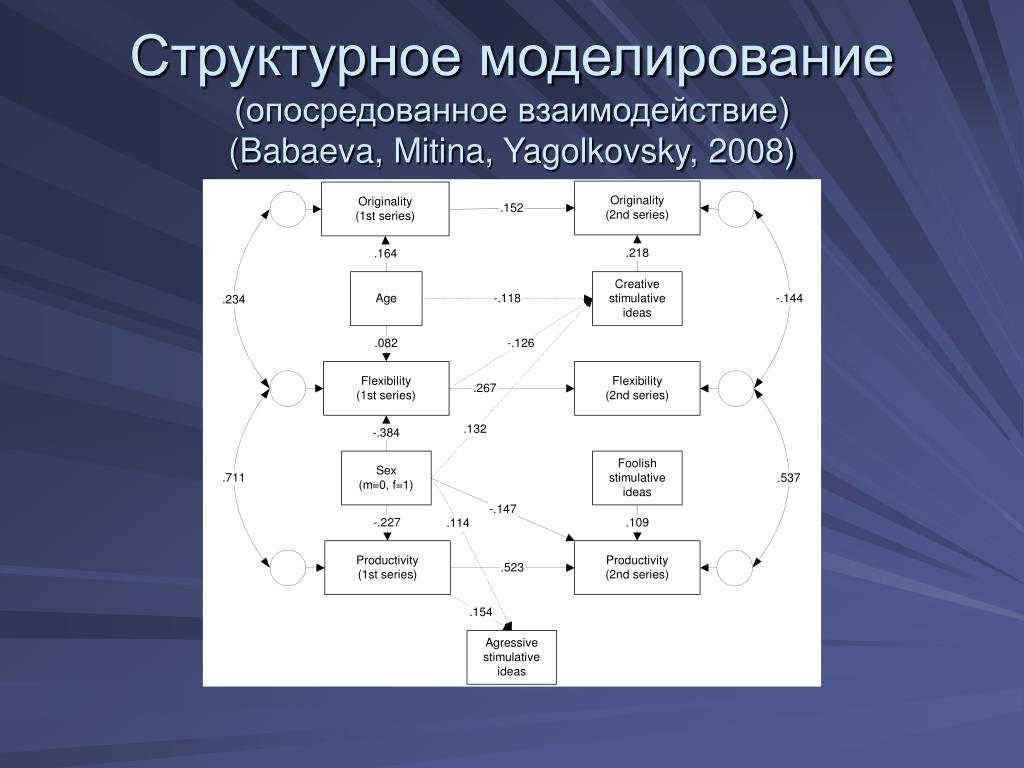 158]
158]
Ксенон непосредственно взаимодействует только с фтором и некоторыми фторидами, например Р1Ре. Фториды ксенона,служат исходными веществами для получения других его соединений. [c.487]
На рис. 1,8, а изобралнепосредственным взаимодействием аппаратурных стадий, т, е. такая, в которой реакционная масса из аппарата предыдущей стадии непосредственно поступает в аппарат следующей стадии. [c.32]
Здесь мы привели примеры моделей синтеза совмещенных ХТС с непосредственным взаимодействием аппаратурных стадий. Аналогичные зависимости могут быть получены, когда между основными аппаратами системы находятся промежуточные гмкости, согласующие их работу. Чтобы сформировать эти модели, необходимо в систему ограничений задачи синтеза одно-тродуктовой системы с промежуточными емкостями (см. предыдущий раздел) внести индекс г, соответствующий номеру продукта. Если продукты при этом производятся последовательно, то в условии выполнения плана появится знак суммы. [c.208]
[c.208]
осуществляет непосредственное взаимодействие с — Translation into English — examples Russian
These examples may contain rude words based on your search.
These examples may contain colloquial words based on your search.
Совет вновь подтвердил, что НОО является той частью институциональной инфраструктуры МЧР, которая осуществляет непосредственное взаимодействие с участниками проектов.
The Board reiterated that a DOE is part of the institutional infrastructure of the CDM, directly interacting with project participants.Suggest an example
В некоторых странах были созданы специальные парламентские комитеты для непосредственного взаимодействия с поставщиками помощи.
In some countries, specialized parliamentary committees have been set up to interact directly with the providers of aid.
Эти должности характеризуются их непосредственным взаимодействием с получателями помощи.
Мы занимались урегулированием этой проблемы в непосредственном взаимодействии с автором методики.
We worked on the problem directly with the author of the methodology.Цель нашей работы — создать возможность непосредственного взаимодействия с океанами из любого места на Земле.
This — I’ve shown you sea floor — but so the goal here is real time interaction with the oceans from anywhere on earth. Данное событие предоставляет нам возможность для непосредственного взаимодействия с молодежью и обмена информацией о наших достижениях, трудностях, надеждах и опасениях.
Они призваны дать посетителям возможность самостоятельного исследования вопросов науки путем непосредственного взаимодействия с познавательными экспонатами.
The exhibitions seek to provide knowledge-oriented, scientific and interactive exhibits and allow visitors to explore science in a hands-on experience.Это вызывает внутреннюю напряженность между его ролью в качестве глобального, базирующегося в Центральных учреждениях подразделения и его непосредственным взаимодействием с национальными правительствами.
This has created an inherent tension between its role as a global, Headquarters-based entity and its direct engagement with national Governments.
Одним из ключевых аспектов этого процесса является непосредственное взаимодействие с правительствами и другими заинтересованными сторонами, действующими на национальном уровне.
Помимо непосредственного взаимодействия с правительствами, мы поддерживаем идею большей национальной ответственности и подотчетности государств перед своими гражданами путем укрепления парламентов и оказания поддержки организациям гражданского общества.
As well as working directly with governments, we support greater country ownership and accountability between States and their citizens by strengthening parliaments and supporting civil society organizations. Увеличение количества посещений стран свидетельствует о том, что Комитет усилил акцент на непосредственном взаимодействии с государствами.
Подготовка каждой рекомендации по вопросу о восстановлении в списке требует проведения множества обзоров в непосредственном взаимодействии с другими департаментами.
Advice for each reinstatement matter requires multiple reviews and close liaison with other departments.Министерство иностранных дел выступает в роли центрального органа, осуществляющего непосредственное взаимодействие с центральными органами других государств.
The central authority is the Ministry of Foreign Affairs, which communicates directly with the central authorities of other States. Эта задача может решаться в партнерстве с местными властями и НПО с учетом их непосредственного взаимодействия с общинами на низовом уровне.
Такая схема обеспечивает непосредственное взаимодействие с населением, а также более широкий диапазон и более высокую эффективность просветительской работы.
Непосредственное взаимодействие с бюро функциональных комиссий также расширяется, что способствует обеспечению более комплексного подхода к последующей деятельности по итогам конференций.
Direct interaction with the bureaux of the functional commissions is also increasing, which contributes to a more integrated follow-up to conferences. Так как спецификация OLEDB не позволяет выполнение команд неинициализированного источника данных, этот запрос возможен только при непосредственном взаимодействии с IBProvider’ом.
В частности, благодаря этому МСП могут налаживать непосредственное взаимодействие с другими МСП в своих и других регионах.
In particular, it allows SMEs to work directly with other SMEs in their regions and in other regions.Их деятельность и размещение предполагали непосредственное взаимодействие с местным населением, от которого их в лучшем случае отделяла колючая проволока.
Their work and accommodations involved direct contact with the local population, separated at most by strands of barbed wire. Они выразили обеспокоенность по поводу сохранения тяжелых условий жизни рома по всей стране и призвали Венгрию продолжать непосредственно взаимодействие с местными НПО и лидерами гражданского общества в деле решения существующих проблем.
Что такое дистанционное обучение? — Dropbox Business
Что такое дистанционное обучение?
Проще говоря, дистанционное обучение — это процесс удаленного получения знаний, не требующий физического присутствия в классе. Исторически понятие «дистанционное обучение» в основном относилось к программам обучения в колледжах, которые позволяли студентам обучаться вне стен учебного заведения. Теперь же дистанционное обучение касается не только студентов колледжей, но и учащихся начальной, средней и старшей школы.
Типы дистанционного обучения
Есть несколько различных типов дистанционного обучения:
Видеоконференции
Самый распространенный формат практических занятий и лекций — видеоконференция или вебинар через Zoom. Иногда такие занятия ведет один учитель, а другой отвечает на вопросы учеников и наблюдает за ними.
Иногда такие занятия ведет один учитель, а другой отвечает на вопросы учеников и наблюдает за ними.
Такой способ обеспечивает прямое взаимодействие между учеником и учителем, благодаря чему этот тип обучения становится наиболее близким к традиционному классному обучению.
Асинхронное обучение
При асинхронном обучении студенты работают автономно. Нет никаких групповых занятий — только еженедельные задания и индивидуальные сроки сдачи. Такой метод самообучения обеспечивает студентам гибкость в организации учебного процесса и составлении расписания, но сводит к минимуму групповое взаимодействие и непосредственное общение с преподавателем.
Свободный график
Учебные курсы со свободным графиком похожи на курсы асинхронного обучения и при этом имеют еще меньше ограничений. Этот вид обучения не предполагает еженедельных сроков сдачи, что позволяет студентам работать в любое удобное время и лучше находить баланс между учебой и обязанностями помимо нее. Это предпочтительный вариант для людей, занятых полный рабочий день, родителей и тех, кто хочет учиться, но не может выделить время для традиционного графика занятий.
Это предпочтительный вариант для людей, занятых полный рабочий день, родителей и тех, кто хочет учиться, но не может выделить время для традиционного графика занятий.
В каждом учебном заведении есть собственные методы и система организации удаленного обучения, которая наверняка соответствует одной из этих категорий или представляет собой их сочетание. Асинхронное обучение и обучение со свободным графиком чаще используются в сфере высшего образования.
Преимущества дистанционного обучения
Учебные курсы со свободным графиком позволяют более гибко выбирать не только место, но и время обучения, что отлично подходит людям, которые работают полный рабочий день. Для многих возможность работать полный рабочий день и учиться дистанционно может быть вариантом, обеспечивающим доступ к образованию.
Что касается затрат, то курсы дистанционного обучения обходятся дешевле, чем обучение на территории учебного заведения, поскольку в этом случае снижается потребность в ресурсах и инфраструктуре.
Иногда курсы дистанционного обучения позволяют изучать предметы или получать квалификацию, которые могут быть недоступны на местном уровне, поскольку можно осваивать курсы колледжей в любой точке мира, не покидая своего дома.
Недостатки дистанционного обучения
Оценивая эффективность дистанционного обучения, важно учитывать индивидуальные особенности студентов. Разным людям удобнее воспринимать информацию по-разному: визуально или вербально, кинестетически или логически, при этом кому-то легче работать в коллективе, а кому-то — индивидуально.
Проводя занятия для целого класса, учителя должны доносить знания с учетом разных типов восприятия. Этого можно добиться в классе, но при дистанционном обучении бывает сложно достучаться до тех, кто воспринимает информацию кинестетически или кому нужен коллектив. Без правильных инструментов совместное обучение также может быть неполноценным.
При дистанционном обучении учителя могут столкнуться с проблемой дисциплины. Во время видеоконференции озорство проявляется также, как и в классе, только на расстоянии его труднее сдерживать.
Во время видеоконференции озорство проявляется также, как и в классе, только на расстоянии его труднее сдерживать.
Дома учащихся может окружать гораздо больше отвлекающих факторов, чем в школе. Во время занятий тяжелее контролировать и пресекать болтовню между учениками, учитывая, что ее можно и не слышать.
Учащимся может быть трудно приспособиться к новой организации обучения, и они легко могут почувствовать себя изолированными и потерянными без непосредственного взаимодействия с учителями и одноклассниками. Серьезной проблемой дистанционного обучения является психическое здоровье: для развития молодых людей необходимы взаимодействие и контакт, и в этом дистанционное обучение может оказаться преградой.
Советы по дистанционному обучению для учащихся:
- Составьте расписание и соблюдайте его. Преимуществом дистанционного обучения является то, что часто вы сами решаете, где и когда учиться, но это может иметь пагубные последствия в случае несоблюдения единого расписания.
 Выделите определенное время для занятий и максимально его придерживайтесь. Если у вас нет расписания, то очень быстро вы можете потерять концентрацию и мотивацию.
Выделите определенное время для занятий и максимально его придерживайтесь. Если у вас нет расписания, то очень быстро вы можете потерять концентрацию и мотивацию. - Выделите место для учебы. По возможности выделите дома место для учебы, отдельное от других жилых зон. Это поможет разделить работу и развлечения. И когда вы будете в этом месте, то будете знать, что находитесь в «классе» и должны вести себя соответствующим образом.
- Подготовьтесь к предстоящему дню. Подготовьтесь утром к предстоящему дню так, как будто вы идете в школу. Вы не сможете мысленно настроиться на школу, если весь день будете сидеть в пижаме. Правильный утренний распорядок задает тон на весь день.
- Ознакомьтесь с инструментами. Если вы не хотите отставать из-за технических трудностей, обязательно выделите время до занятий онлайн, чтобы ознакомиться с технологиями дистанционного обучения.
- Делайте перерывы. Составляя свое расписание, не забудьте о перерывах. Непрерывная учеба часами напролет может ухудшать восприятие, ведь вам нужно время для усвоения изученного материала.
- Будьте в курсе стоящих перед вами задач. В отличие от занятий в классе, перед вами нет учителя, который будет напоминать о предстоящих сроках сдачи заданий или зачетах. Программа дистанционного обучения предполагает, что вы будете в курсе стоящих перед вами задач. У вас должен быть точный календарь, где установлены напоминания, помогающие отслеживать, что должно быть сделано.
- Оставайтесь на связи с одноклассниками. Если виртуальные занятия не предполагают особого взаимодействия, подумайте о создании учебной группы, чтобы оставаться на связи с одноклассниками и быть уверенными, что вы все на правильном пути. Воспользуйтесь преимуществами инструментов для совместного обучения, таких как Dropbox Paper, чтобы легко делиться заметками, файлами проектов, курсовыми работами и улучшить групповое общение.
 Выделите определенное время для занятий и максимально его придерживайтесь. Если у вас нет расписания, то очень быстро вы можете потерять концентрацию и мотивацию.
Выделите определенное время для занятий и максимально его придерживайтесь. Если у вас нет расписания, то очень быстро вы можете потерять концентрацию и мотивацию. Непрерывная учеба часами напролет может ухудшать восприятие, ведь вам нужно время для усвоения изученного материала.
Непрерывная учеба часами напролет может ухудшать восприятие, ведь вам нужно время для усвоения изученного материала.Советы по дистанционному обучению для учителей:
- Сделайте образовательные технологии своим помощником. Переход к обучению онлайн пугает, но вы быстро справитесь со своим страхом, познакомившись с цифровой средой. Воспользуйтесь преимуществами приложений, инструментов и служб, которые помогают в обучении онлайн.
- Поощряйте совместную работу. Мыслите нестандартно, чтобы разрабатывать совместные проекты, способствующие социальному обучению. Используйте онлайн-инструменты, такие как Dropbox Paper, чтобы упростить групповую совместную работу и оценку.
- Общайтесь с учащимися. Если можете, общайтесь с каждым учащимся напрямую. Конечно, это будет непросто, особенно если ваш класс больше обычного, но если вам это удастся, то такое личное взаимодействие будет мотивировать ребят и поможет вам выявить стоящие перед вами задачи. Если личное общение невозможно, обязательно свяжитесь с вашими учениками по электронной почте или отправьте им мгновенные сообщения с напоминанием, что вы готовы дать индивидуальную консультацию.
- Регулярно оставляйте отзывы. Отзыв помогает мотивировать учащихся и дает им возможность распознавать свои сильные и слабые стороны, способствуя росту и развитию. Это особенно важно при удаленном обучении, когда учащиеся могут чувствовать себя изолированными и не знать о своей успеваемости.
- Общайтесь с другими учителями. Так же, как родители должны общаться с родителями, а учащиеся с учащимися, так и учителя должны общаться друг с другом и делиться своими впечатлениями, советами и переживаниями.
- Будьте последовательными. Как только вы определили систему работы, придерживайтесь ее и старайтесь быть последовательными в отношении используемых инструментов и планов занятий. Поначалу виртуальное обучение может показаться хаотичным, но последовательный подход поможет нормализовать ситуацию для учащихся и сформировать у них правильный настрой.
- Обращайтесь за отзывом. Получить отзыв так же важно, как и оставить его, особенно при испытании новой системы. Чтобы узнать, что подходит вашим ученикам, нужно получить честный отзыв о том, нравятся ли им используемые вами инструменты и приносят ли они пользу.
 Переход к обучению онлайн пугает, но вы быстро справитесь со своим страхом, познакомившись с цифровой средой. Воспользуйтесь преимуществами приложений, инструментов и служб, которые помогают в обучении онлайн.
Переход к обучению онлайн пугает, но вы быстро справитесь со своим страхом, познакомившись с цифровой средой. Воспользуйтесь преимуществами приложений, инструментов и служб, которые помогают в обучении онлайн. Отзыв помогает мотивировать учащихся и дает им возможность распознавать свои сильные и слабые стороны, способствуя росту и развитию. Это особенно важно при удаленном обучении, когда учащиеся могут чувствовать себя изолированными и не знать о своей успеваемости.
Отзыв помогает мотивировать учащихся и дает им возможность распознавать свои сильные и слабые стороны, способствуя росту и развитию. Это особенно важно при удаленном обучении, когда учащиеся могут чувствовать себя изолированными и не знать о своей успеваемости.
Существует множество нюансов и последствий, которые трудно предвидеть, поскольку дистанционное обучение пока только развивается. Может, нас ждет будущее, в котором обучение на дому будет таким надежным, что нам не придется переживать из-за закрытия школ в снегопад. Являются ли цифровые диорамы будущим научных проектов? Время покажет, как развиваются форматы, но если вы прислушаетесь к вышесказанному, вам будет легче адаптироваться.
Типовые ситуации | Порядок урегулирования конфликта интересов |
| Выполнение отдельных функций государственного управления в отношении родственников и/или иных лиц, с которыми связана личная заинтересованность государственного служащего | |
Государственный служащий участвует в осуществлении отдельных функций государственного управления и/или в принятии кадровых решений в отношении родственников и/или иных лиц, с которыми связана личная заинтересованность государственного служащего, например:
| Государственному служащему следует уведомить о наличии личной заинтересованности представителя нанимателя и непосредственного начальника в письменной форме. Представителю нанимателя рекомендуется отстранить государственного служащего от исполнения должностных обязанностей, предполагающих непосредственное взаимодействие с родственниками и/или иными лицами, с которыми связана личная заинтересованность государственного служащего. Например, рекомендуется временно вывести государственного служащего из состава конкурсной комиссии, если одним из кандидатов на замещение вакантной должности государственной службы является его родственник. При этом далеко не любое выполнение функций государственного управления в отношении родственников влечет конфликт интересов. Если государственный служащий предоставляет государственные услуги, получение которых одним заявителем (родственником) не влечет отказа в предоставлении услуги другим заявителям, и при этом он не обладает полномочиями, позволяющими оказывать кому-либо предпочтение, вероятность возникновения конфликта интересов в большинстве случаев является незначительной. |
| Выполнение иной оплачиваемой работы | |
| Государственному служащему следует уведомить о наличии личной заинтересованности представителя нанимателя и непосредственного начальника в письменной форме, отказаться от выполнения иной оплачиваемой работы в организации, в отношении которой государственный служащий осуществляет отдельные функции государственного управления. В случае если на момент начала выполнения отдельных функций государственного управления в отношении организации родственники государственного служащего выполняют в ней оплачиваемую работу, следует уведомить о наличии личной заинтересованности представителя нанимателя и непосредственного начальника в письменной форме. В случае если государственный служащий самостоятельно не предпринял мер по урегулированию конфликта интересов, представителю нанимателя рекомендуется отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении организации, в которой государственный служащий или его родственники выполняют иную оплачиваемую работу. |
| В уведомлении государственному служащему следует полно и подробно изложить, в какой степени выполнение им этой работы связано с его должностными обязанностями. Рекомендуется отказаться от выполнения иной оплачиваемой работы в этой организации. В случае если на момент начала выполнения отдельных функций государственного управления в отношении организации, получающей платные услуги, родственники государственного служащего уже выполняли оплачиваемую работу в организации, оказывающей платные услуги, следует уведомить о наличии личной заинтересованности представителя нанимателя и непосредственного начальника в письменной форме. Представителю нанимателя рекомендуется подробно рассмотреть обстоятельства выполнения государственным служащим иной оплачиваемой работы. Особое внимание следует уделять фактам, указывающим на возможное использование государственным служащим своих полномочий для получения дополнительного дохода, например:
При обнаружении подобных фактов представителю нанимателя рекомендуется принять решение о том, что выполнение иной оплачиваемой работы влечет конфликт интересов и отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении организации, получающей платные услуги. |
| Представителю нанимателя рекомендуется указать государственному служащему, что выполнение подобной иной оплачиваемой работы влечет конфликт интересов. В случае если государственный служащий не предпринимает мер по урегулированию конфликта интересов и не отказывается от личной заинтересованности, рекомендуется рассмотреть вопрос об отстранении государственного служащего от замещаемой должности. |
| При направлении представителю нанимателя предварительного уведомления о выполнении иной оплачиваемой работы государственному служащему следует полно изложить, каким образом организация, в которой он собирается выполнять иную оплачиваемую работу, связана с организациями, в отношении которых он осуществляет отдельные функции государственного управления. При этом рекомендуется отказаться от выполнения иной оплачиваемой работы в материнских, дочерних и иным образом аффилированных организациях. В случае если на момент начала выполнения отдельных функций государственного управления в отношении организации родственники государственного служащего уже выполняли оплачиваемую работу в аффилированной организации, следует уведомить о наличии личной заинтересованности представителя нанимателя и непосредственного начальника в письменной форме. Представителю нанимателя рекомендуется отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении организации, являющейся материнской, дочерней или иным образом аффилированной с той организацией, в которой государственный служащий выполняет иную оплачиваемую работу. |
| Принятие решений о закупке товаров, выполнении услуг | |
Государственный служащий участвует в принятии решения о закупке государственным органом товаров, являющихся результатами интеллектуальной деятельности, исключительными правами на которые обладает он сам, его родственники или иные лица, с которыми связана личная заинтересованность государственного служащего. | Государственному служащему следует уведомить о наличии личной заинтересованности представителя нанимателя и непосредственного начальника в письменной форме. При этом рекомендуется отказаться от участия в соответствующем конкурсе. Представителю нанимателя рекомендуется вывести государственного служащего из состава комиссии по размещению заказа на время проведения конкурса. |
| Владение ценными бумагами, банковскими вкладами | |
| В случае если государственный служащий Государственный служащий (его родственники) владеет ценными бумагами организации, в отношении которой он осуществляет отдельные функции государственного управления, он обязан уведомить представителя нанимателя и непосредственного начальника о наличии личной заинтересованности в письменной форме, а также передать (рекомендовать родственникам передать) ценные бумаги в доверительное управление либо рассмотреть вопрос об их отчуждении. До принятия мер по урегулированию конфликта интересов представителю нанимателя рекомендуется отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении организации, ценными бумагами которой владеет государственный служащий или его родственники. |
| Государственному служащему следует уведомить о наличии личной заинтересованности представителя нанимателя и непосредственного начальника в письменной форме. Представителю нанимателя рекомендуется до принятия государственным служащим мер по урегулированию конфликта интересов отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении банков и кредитных организаций, вкоторых сам государственный служащий, его родственники или иные лица, с которыми связана его личная заинтересованность, имеют вклады либо взаимные обязательства, связанные с оказанием финансовых. |
| Получение подарков и услуг | |
| Государственному служащему следует уведомить представителя нанимателя и непосредственного начальника в письменной форме о наличии личной заинтересованности. Следует оценить, действительно ли оказание услуг может привести к необъективному исполнению государственным служащим его должностных обязанностей. Если вероятность конфликта интересов высока, рекомендуется отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении указанных физических лиц или организаций. |
| Государственному служащему и его родственникам рекомендуется не принимать подарки от организаций, в отношении которых государственный служащий осуществляет или ранее осуществлял отдельные функции государственного управления, вне зависимости от стоимости этих подарков и поводов дарения. Представителю нанимателя, в случае если ему стало известно о получении государственным служащим такого подарка, необходимо оценить, насколько полученный подарок связан с исполнением государственным служащим его должностных обязанностей. Если подарок связан с исполнением должностных обязанностей, то в отношении государственного служащего должны быть применены меры дисциплинарной ответственности, учитывая характер совершенного государственным служащим коррупционного правонарушения, его тяжесть, обстоятельства, при которых оно совершено, соблюдение государственным служащим других ограничений и запретов, требований о предотвращении или об урегулировании конфликта интересов и исполнение им обязанностей, установленных в целях противодействия коррупции, а также предшествующие результаты исполнения государственным служащим своих должностных обязанностей. Если подарок не связан с исполнением должностных обязанностей, то государственному служащему рекомендуется указать на то, что получение подарков от заинтересованных физических лиц и организаций может нанести урон репутации государственного органа, и поэтому является нежелательным вне зависимости от повода дарения. В случае если представитель нанимателя обладает информацией о получении родственниками государственного служащего подарков от физических лиц и/или организаций, в отношении которых государственный служащий осуществляет или ранее осуществлял отдельные функции государственного управления, рекомендуется:
|
| Государственному служащему рекомендуется не принимать подарки от непосредственных подчиненных вне зависимости от их стоимости и повода дарения. Особенно строго следует подходить к получению регулярных подарков от одного дарителя. Представителю нанимателя, которому стало известно о получении государственным служащим подарков от непосредственных подчиненных, следует указать государственному служащему на то, что подобный подарок может рассматриваться как полученный в связи с исполнением должностных обязанностей, в связи с чем подобная практика может повлечь конфликт интересов, а также рекомендовать государственному служащему вернуть полученный подарок дарителю в целях предотвращения конфликта интересов. |
| Конфликт интересов, связанный с имущественными обязательствами и судебными разбирательствами | |
| Государственному служащему и его родственникам рекомендуется урегулировать имеющиеся имущественные обязательства (выплатить долг, расторгнуть договор аренды и т.д.). При невозможности сделать это, государственному служащему следует уведомить представителя нанимателя и непосредственного начальника о наличии личной заинтересованности в письменной форме. Представителю нанимателя рекомендуется по крайней мере до урегулирования имущественного обязательства отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении организации, перед которой сам государственный служащий, его родственники или иные лица, с которыми связана личная заинтересованность государственного служащего, имеют имущественные обязательства. |
| Государственному служащему следует уведомить представителя нанимателя и непосредственного начальника о наличии личной заинтересованности в письменной форме. Представителю нанимателя рекомендуется отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении кредиторов организации, владельцами или сотрудниками которых являются родственники государственного служащего или иные лица, с которыми связана личная заинтересованность государственного служащего. |
| Государственному служащему следует уведомить представителя нанимателя и непосредственного начальника в письменной форме о наличии личной заинтересованности. Представителю нанимателя рекомендуется по крайней мере до урегулирования имущественного обязательства отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении организации, которая имеет имущественные обязательства перед государственным служащим, его родственниками, или иными лицами, с которыми связана личная заинтересованность государственного служащего. |
| Государственному служащему следует уведомить представителя нанимателя и непосредственного начальника в письменной форме о наличии личной заинтересованности. Представителю нанимателя рекомендуется отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении физических лиц и организаций, которые находятся в стадии судебного разбирательства с государственным служащим, его родственниками или иными лицами, с которыми связана личная заинтересованность государственного служащего. |
| Взаимодействие с бывшим работодателем и трудоустройство после увольнения с государственной службы | |
| Государственному служащему в случае поручения ему отдельных функций государственного управления в отношении организации, в которой он работал до поступления на государственную службу, рекомендуется уведомить об этом представителя нанимателя и непосредственного начальника в письменной форме. Представителю нанимателя рекомендуется оценить, могут ли взаимоотношения государственного служащего с бывшим работодателем повлиять на объективное исполнение должностных обязанностей и повлечь конфликт интересов. В случае если эта вероятность есть, рекомендуется отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении бывшего работодателя. |
| Государственному служащему рекомендуется воздерживаться от ведения переговоров о последующем трудоустройстве с организациями, в отношении которых он осуществляет отдельные функции государственного управления, до момента увольнения с государственной службы. Если указанные переговоры начались, государственному служащему следует уведомить представителя нанимателя и непосредственного начальника в письменной форме о наличии личной заинтересованности. Представителю нанимателя рекомендуется отстранить государственного служащего от исполнения должностных (служебных) обязанностей в отношении организации, с которой он ведет переговоры о трудоустройстве после увольнения с государственной службы. Возможны ситуации, которые могут повлечь конфликт интересов и нанести ущерб репутации государственного органа, но при этом не могут быть урегулированы в рамках действующего законодательства:
|
| Нарушения запретов, установленных законодательством | |
| В соответствии с п. 11 ч. 1 ст. 17 Федерального закона № 79-ФЗ гражданскому служащему запрещается принимать без письменного разрешения представителя нанимателя награды, почетные и специальные звания иностранных государств, международных организаций, а также политических партий, других общественных объединений и религиозных объединений, если в его должностные обязанности входит взаимодействие с указанными организациями и объединениями. Представителю нанимателя при принятии решения о предоставлении или непредоставлении разрешения рекомендуется уделить особое внимание основанию и цели награждения, а также тому, насколько получение гражданским служащим награды, почетного и специального звания может породить сомнение в его беспристрастности и объективности. |
| В соответствии с п. 17 ч. 1 ст. 17 Федерального закона № 79-ФЗ гражданскому служащему запрещается заниматься без письменного разрешения представителя нанимателя оплачиваемой деятельностью, финансируемой исключительно за счет средств иностранных государств, международных и иностранных организаций, иностранных граждан и лиц без гражданства, если иное не предусмотрено международным договором Российской Федерации или российским законодательством. Представителю нанимателя при принятии решения об указанном разрешении рекомендуется выяснить, насколько выполнение гражданским служащим иной оплачиваемой работы может породить сомнение в его беспристрастности и объективности, а также «выяснить» какую именно работу он там выполняет. |
| Государственному служащему при выявлении нарушений законодательства в ходе контрольно-надзорных мероприятий рекомендуется воздержаться от дачи советов относительно того, какие организации могут быть привлечены для устранения этих нарушений. |
| Государственному служащему запрещается разглашать или использовать в целях, не связанных с государственной службой, сведения, отнесенные в соответствии с федеральным законом к сведениям конфиденциального характера, или служебную информацию, ставшие ему известными в связи с исполнением должностных обязанностей. Указанный запрет распространяется в том числе и на использование неконфиденциальной информации, которая лишь временно недоступна широкой общественности. В связи с этим государственному служащему следует воздерживаться от использования в личных целях сведений, ставших ему известными в ходе исполнения служебных обязанностей, до тех пор, пока эти сведения не станут достоянием широкой общественности. Представителю нанимателя, которому стало известно о факте использования государственным служащим информации, полученной в ходе исполнения служебных обязанностей и временно недоступной широкой общественности, для получения конкурентных преимуществ при совершении коммерческих операций, рекомендуется рассмотреть вопрос о применении к государственному служащему мер дисциплинарной ответственности за нарушение запретов, связанных с государственной службой, учитывая характер совершенного государственным служащим коррупционного правонарушения, его тяжесть, обстоятельства, при которых оно совершено, соблюдение государственным служащим других ограничений и запретов, требований о предотвращении или об урегулировании конфликта интересов и исполнение им обязанностей, установленных в целях противодействия коррупции, а также предшествующие результаты исполнения государственным служащим своих должностных обязанностей. В случае установления признаков дисциплинарного проступка либо факта совершения государственным служащим деяния, содержащего признаки административного правонарушения или состава преступления, данная информация представляется руководителю государственного органа для решения вопроса о проведении служебной проверки и применении мер ответственности, предусмотренных нормативными правовыми актами Российской Федерации, либо передается в правоохранительные органы по подведомственности. |
 При этом одним из кандидатов на вакантную должность в этом государственном органе является родственник государственного служащего;
При этом одним из кандидатов на вакантную должность в этом государственном органе является родственник государственного служащего;







 ).
).








 Следует также учитывать требования статья 12 Федерального закона от 25.12.2008 № 273-ФЗ «О противодействии коррупции».
Следует также учитывать требования статья 12 Федерального закона от 25.12.2008 № 273-ФЗ «О противодействии коррупции».




Страховые представители на страже интересов пациентов » ТФОМС
На сегодняшний день высока ответственность страховой медицинской организации по охране прав своих застрахованных лиц. Для доступной адресной консультативной помощи в сфере обязательного медицинского страхования в регионе сформирована и успешно функционирует служба страховых представителей. На защите наших граждан стоят профессионалы 3-х уровней.
Страховой представитель – это сотрудник компании, в задачу которого входит непосредственное взаимодействие с гражданами и помощь при получении медицинской помощи в рамках программы обязательного медицинского страхования.
Если Вы позвоните по телефону горячей линии Контакт-центра 8-800-100-73-09, то услышите страхового представителя 1 уровня. Он примет и зарегистрирует обращение, ответит на вопросы справочно-консультационного характера. Специалист подробно расскажет:
— где и кому можно получить полис ОМС, какие документы предъявить;
— в каких случаях необходимо уведомить свою страховую компанию;
— адреса и номера телефонов ближайших офисов страховых организаций;
— что делать с полисом ОМС военнослужащим;
— какие медицинские организации работают в сфере ОМС в нашем регионе;
— в каких случаях пациент имеет право выбрать лечебное учреждение и врача;
— какая медицинская помощь пациенту положена по полису ОМС;
— какие обследования проводятся в рамках диспансеризации в поликлинике.
Если вопрос более конкретный, требующий страховой идентификации гражданина или работы по фактам нарушения оказания медицинской помощи, несоблюдении сроков или отказа в ней в определенном лечебном учреждении, Ваш звонок будет переадресован на страхового представителя 2 уровня. Эти «поверенные» прошли соответствующую подготовку по знанию порядков и стандартов медицинской помощи. В их функцию входит организация работы с гражданами по информированию и сопровождению при оказании медицинской помощи, по защите законных интересов в сфере ОМС.
Эти «поверенные» прошли соответствующую подготовку по знанию порядков и стандартов медицинской помощи. В их функцию входит организация работы с гражданами по информированию и сопровождению при оказании медицинской помощи, по защите законных интересов в сфере ОМС.
Высшее звено института страховых поверенных – страховые представители 3 уровня. Это специалисты с высшим медицинским образованием, стажем работы во врачебной специальности и соответствующей подготовкой.
Все спорные и сложные случаи на основе письменных обращений, связанных с доступностью и качеством медицинской помощи, ложатся на их стол. Они проводят экспертизы на основании жалоб граждан с последующим информированием их о результатах и выявленных нарушениях. При необходимости организуют правовую поддержку интересов пациентов в рамках досудебного и судебного разрешения спорных вопросов.
Эксперты осуществляют тематические экспертизы на предмет выявления нарушений условий оказания медицинской помощи, в том числе сроков её ожидания, пациентам со злокачественными новообразованиями, пациентам, взятым на диспансерный учет по результатам проведенной диспансеризации. В случае нарушения сроков либо отказе в госпитализации проводят экспертные мероприятия для выяснения причин и восстановления нарушенных прав граждан.
В случае нарушения сроков либо отказе в госпитализации проводят экспертные мероприятия для выяснения причин и восстановления нарушенных прав граждан.
Страховые представители находятся на постах непосредственно в медицинских организациях, при организации мобильных представительств, выездных акций и поездов здоровья.
Подробную информацию о работе страховых представителей можно получить в страховой компании, выдавшей Вам полис ОМС.
Коронавирус: как поменяется работа парикмахерских после карантина
Автор фото, Getty Images
Подпись к фото,Парикмахерские стали первыми заведениями, открывшимися после смягчения карантина
Карантинные меры в некоторых странах постепенно ослабляют, парикмахерские и салоны красоты скоро вновь откроют двери. Как изменятся правила работы с клиентами в условиях необходимости соблюдать социальную дистанцию — разбираемся на примере Германии, где парикмахерские уже заработали.
Германия ослабила карантин, позволив открыться парикмахерским, которые здесь, как и по всей Европе, только лишь примеряются к новым условиям работы.
Никакого ожидания с перелистыванием глянцевых журналов, никакой стрижки на сухие волосы, и всё — только в маске, причем и для клиента, и для мастера.
По нормам, установленным германским министерством экономики и социальных проблем, между клиентами должно сохраняться расстояние в полтора метра и в идеале — никакой укладки.
Парикмахер должен находиться в перчатках до того момента, как волосы клиента помыты (чтобы были смыты все бактерии), но, чтобы сделать стрижку, перчатки можно снимать.
И придется обойтись без привычных сплетен: из-за маски все равно ничего не разберешь. Лицом к лицу разговаривать нельзя: все переговоры по поводу цвета волос и длины надо вести только через зеркало, да и то по минимуму.
Первая проба ножниц
В министерстве говорят, что эти нормы необходимы, потому что по своей природе стрижка — вещь интимная, подразумевающая непосредственное взаимодействие с клиентом.
Здесь контакт гораздо ближе, чем в магазине между продавцом и покупателем.
Автор фото, Getty Images
Подпись к фото,Эксперты полагают, что цены на парикмахерские услуги неизбежно возрастут
Салоны также обязаны записывать имена и телефоны своих клиентов, чтобы можно было их проинформировать, если вдруг кто-то заболеет.
По всей Европе парикмахерские салоны стали первыми, кому разрешено возобновить свой бизнес.
В Испании салоны также открылись на этой неделе, 4 мая, но их загруженность не должна превышать 30% от обычного клиентского потока, и принимать они могут только по предварительной записи.
Когда в Дании 17 апреля было объявлено о начале работы парикмахерских салонов, то сразу рухнул крупнейший в стране интернет-портал по бронированию услуг: как написала одна местная газета, от слишком большого наплыва лохматых граждан, жаждущих постричься.
Какие правила в салонах введены в Германии:
- Стрижка только по записи, никакого ожидания в салоне
- Клиенты и мастера должны соблюдать дистанцию в 1,5 метра, но не во время собственно стрижки
- Клиенты и сотрудники салонов должны быть в масках
- Прямое общение не допускается — разговаривать о прическе, цвете волос и т. п. можно только через зеркало и по минимуму
- По возможности обходиться без укладки
- В салоне должен быть свежий воздух в объеме 100 кубометров в час на каждого парикмахера;
- Ножницы и другие приборы должны дезинфицироваться между клиентами, как и кресла;
- Фартуки и накидки парикмахеров должны стираться после каждого использования или же поверх них необходимо надевать одноразовую накидку;
- Клиенты, приходя в салон, обязаны мыть руки с мылом.
 п. можно только через зеркало и по минимуму
п. можно только через зеркало и по минимумуИсточник: Berufsgenossenschaft für Gesundheitsdienst und Wohlfahrtspflege
Корреспондент Би-би-си в Берлине Дамиен Макгиннес поинтересовался у совладельца берлинского салона Harms Рено Хармса, в каком виде к нему приходят сейчас его клиенты.
«В довольно смешном. Многие сами себе стригли волосы. Некоторые даже и красились, так что мы сейчас в основном проводим работу над ошибками», — говорит Рено.
За время шестинедельного локдауна многие берлинцы купили себе ваучеры с 50%-й скидкой, чтобы использовать их, когда салоны откроются, а это фактически означает, что Рено и многие его коллеги работают в первые дни бесплатно, поскольку клиенты отоваривают свои ваучеры.
«Снимите завязки с ушей»
Но насколько легко в целом будет следовать новым правилам, и как они могут повлиять на наши с вами прически?
Автор фото, Getty Images
Подпись к фото,Общаться в мастером можно только через зеркало. Или телепатически
Алисон Бадрик, парикмахер из английского графства Бакингэмпшир, работающая как индивидуальный предприниматель, все еще ждет каких-либо разъяснений от британских властей по поводу того, когда она сможет вернуться к работе и начать снова ездить к клиентам.
Ее волнует, что это может произойти нескоро. По ее словам, все перечисленные выше европейские правила разумны, но им трудно следовать, особенно в том, что касается подстригания волос, когда у клиента медицинская маска на лице.
Как она поясняет, дело в том, завязки или резинки от маски будут мешать создавать многослойную стрижку или же стричь за ушами.
«Я не знаю, как это будет возможно, если только они не придумают какую-нибудь такую маску, которая бы просто крепилась на лицо», — говорит Алисон.
Глава Ассоциации парикмахеров Германии в интервью телепрограмме Tagesschau пояснил, что клиенты смогут на короткое время снять завязки с ушей и просто держать маску у лица.
Как быть мужчинам, которые захотят привести в порядок растительность на лице, пока совершенно непонятно.
Какой бы для вас ни была процедура посещения салона до карантина — расслабляющей или стрессовой — после все будет казаться иначе.
Новые нормы социального дистанцирования означают, что салоны не смогут принимать прежнее количество клиентов в день.
По прикидкам лондонской сети парикмахерских Blue Tit, их салоны будут работать вполовину от прежнего клиентопотока.
Эта сеть также рассчитывает принимать клиентов с интервалом в 15 минут — чтобы иметь возможность всё тщательно вымыть, — а также увеличить часы работы и работать в две смены.
Они надеются, что смогут предложить и некоторые «излишества» типа массажа головы и горячих напитков.
Но ясно, что каковы бы ни были новые правила, цена на услуги неминуемо возрастет.
«Более продолжительное обслуживание, как и расходы на средства защиты, повлекут за собой большие расходы с нашей стороны, но безопасность наших клиентов и сотрудников — для нас самое главное», — говорят в сети салонов Blue Tit.
А как насчет потрепаться?
«Все дело в том, что парикмахеры любят поговорить — это одно из главных качеств парикмахера, — признает Алисон Бадрик. — Разговор по душам — это часть парикмахерской услуги. Поэтому интересно посмотреть, как все это будет происходить».
Обучение с применением дистанционных образовательных технологий — Образование
Обучение с применением дистанционных образовательных технологийВ связи с переходом учителей школ на удаленную работу из дома в соответствии с Постановлением Правительства СПб №182 от 03. 04.2020 записи в электронном дневнике временно будут отсутствовать. Ведется работа по удаленному доступу учителей СПб к электронному журналу. О восстановлении регулярного заполнения электронного дневника будет объявлено дополнительно.
04.2020 записи в электронном дневнике временно будут отсутствовать. Ведется работа по удаленному доступу учителей СПб к электронному журналу. О восстановлении регулярного заполнения электронного дневника будет объявлено дополнительно.
Прокофьева Наталья Анатольевна, +7 (911) 224-00-27
Андреева Светлана Олеговна, +7 (921) 649-58-74
Дистанционные технологии обучения (образовательного процесса) представляют собой совокупность методов, средств обучения и администрирования учебных процедур, обеспечивающих проведение учебного процесса на расстоянии на основе использования современных информационных и телекоммуникационных технологий. Использование дистанционных технологий предполагает специальную организацию образовательного процесса, базирующуюся на принципе самостоятельного обучения. Среда обучения характеризуется тем, что учащиеся в основном, а часто и совсем, отдалены от преподавателя в пространстве и (или) во времени, в то же время они имеют возможность в любой момент поддерживать диалог с помощью средств телекоммуникации.
Среда обучения характеризуется тем, что учащиеся в основном, а часто и совсем, отдалены от преподавателя в пространстве и (или) во времени, в то же время они имеют возможность в любой момент поддерживать диалог с помощью средств телекоммуникации.
Основное достоинство дистанционных технологий в обучении детей с ограниченными возможностями состоит в отсутствии строгой привязки к месту и времени проведения занятий.
Во-вторых, появляется возможность организовать щадящий режим обучения, сокращая количество часов учебной нагрузки, нормируя количество времени, проводимого за компьютером, многократно возвращаясь к изучаемому материалу при необходимости.
В-третьих, дистанционные технологии в определенной степени разрешают основную проблему «особых» детей, которая заключается в недостатке общения с другими людьми и, в особенности, со сверстниками. Несмотря на физическую удаленность субъектов обучения друг от друга, существует реальная возможность взаимного общения детей в рамках курсовых и тематических совместных занятий как по вертикали (педагог — обучающийся), так и по горизонтали (между обучающимися, в режиме электронной почты, конференций, чата, виртуальных семинаров и т. п.).
п.).
Обучение с применением дистанционных образовательных технологий не следует рассматривать как единственную форму проведения учебных занятий, полностью исключающую непосредственное взаимодействие обучающегося с учителем (учителями). Оптимальным является сочетание дистанционной формы обучения с посещением обучающихся на дому учителем. Для детей, состояние здоровья которых допускает возможность периодического посещения ими образовательного учреждения, наряду с дистанционным обучением и занятиями на дому целесообразно организовывать занятия в помещениях образовательного учреждения.
Особенности организации образовательного процесса для каждого обучающегося, включают объем его учебной нагрузки, а также соотношение объема проведенных занятий с использованием дистанционных образовательных технологий или путем непосредственного взаимодействия учителя с обучающимся, определяются индивидуально на основании рекомендаций специалистов.
Нормативно-правовые документы
Расписание на период обучения с применением дистанционных образовательных технологий
Отчеты о реализации обучения с применением дистанционных образовательных технологий и электронного обучения
Основными формами дистанционного обучения школьников являются:
- использование образовательных интернет-ресурсов
- использование ресурсов, созданных учителями школы
- WEB-консультации
- общение с учителем через электронную почту
- использование специализированных порталов дистанционного обучения, например:
Информационные ресурсы для обучающихся
Ресурсы дистанционных форм обучения:
Образовательные порталы:
- http://www. gramota.ru – Cправочно-информационый портал «Грамота.ru».
- http://www.ucheba.com – Образовательный портал «УЧЕБА”.
- http://www.alledu.ru – «Всё образование в интернет». Образовательный информационный портал. Каталог ссылок, новости, детская общественная приёмная, информация о лидерах образования, анонсы, объявления о работе.
- http://www.distance-learning.ru – Информационный портал «Дистанционное образование».
- http://college.ru – первый в России образовательный Internet-портал, включающий обучение школьников (математика, физика, астрономия, химия, биология и другие предметы) и курсы для профессионального образования.
- href=http://ya-i-mir.ru/ — Конкурс дистанционных проектов «Я познаю мир».
 gramota.ru – Cправочно-информационый портал «Грамота.ru».
gramota.ru – Cправочно-информационый портал «Грамота.ru».Информационные ресурсы для педагогических работников
Система дистанционного обучения ИМЦ Адмиралтейского района >>>
Дистанционные курсы повышения квалификации и профессиональной переподготовки. Конспекты уроков, презентации, тесты, видеоуроки и другие материалы по предметам школьной программы.
 https://infourok.ru/
https://infourok.ru/Курсы повышения квалификации СПбЦОКОиИТ >>>>
Портал дистанционного обучения СПбЦОКОиИТ >>>>
Для учителей запущен бесплатный ресурс по дистанционному обучению
На портале «Учись дома онлайн» открыт бесплатный онлайн-курс «Дистанционное обучение: организация процесса и использование бесплатных приложений, курсов, видеолекций».
Цель курса – научить педагогов школ и вузов пользоваться дистанционными форматами обучения, показать, как они могут создать образовательный контент, показать лучшие бесплатные ресурсы, которые можно использовать для работы со студентами и школьниками.
Модули курса:
— Образовательные ресурсы для педагога: сайты, приложения, видеолекции, курсы и интерактивные приложения;
— Технологии проведения дистанционного образования;
— Как создать свой контент для дистанционного образования: методики и векторы развития онлайн образования.
На сайте также есть «банк», в котором размещены полезные ресурсы и курсы, которые педагоги могут использовать в образовательном процессе. С уважением,методисты.
Безопасность | Стеклянная дверь
Мы получаем подозрительную активность от вас или от кого-то, кто использует вашу интернет-сеть. Подождите, пока мы убедимся, что вы настоящий человек. Ваш контент появится в ближайшее время. Если вы продолжаете видеть это сообщение, напишите нам чтобы сообщить нам, что у вас проблемы.
Nous aider à garder Glassdoor sécurisée
Nous avons reçu des activités suspectes venant de quelqu’un utilisant votre réseau internet. Подвеска Veuillez Patient que nous vérifions que vous êtes une vraie personne.Вотре содержание apparaîtra bientôt. Si vous continuez à voir ce message, veuillez envoyer un электронная почта à pour nous informer du désagrément.
Unterstützen Sie uns beim Schutz von Glassdoor
Wir haben einige verdächtige Aktivitäten von Ihnen oder von jemandem, der in ihrem
Интернет-Netzwerk angemeldet ist, festgestellt. Bitte warten Sie, während wir
überprüfen, ob Sie ein Mensch und kein Bot sind. Ihr Inhalt wird в Kürze angezeigt.
Wenn Sie weiterhin diese Meldung erhalten, informieren Sie uns darüber bitte по электронной почте:
.
Bitte warten Sie, während wir
überprüfen, ob Sie ein Mensch und kein Bot sind. Ihr Inhalt wird в Kürze angezeigt.
Wenn Sie weiterhin diese Meldung erhalten, informieren Sie uns darüber bitte по электронной почте:
.
We hebben verdachte activiteiten waargenomen op Glassdoor van iemand of iemand die uw internet netwerk deelt. Een momentje geduld totdat, мы исследовали, что u daadwerkelijk een persoon bent. Uw bijdrage zal spoedig te zien zijn. Als u deze melding blijft zien, электронная почта: om ons te laten weten dat uw проблема zich nog steeds voordoet.
Hemos estado detectando actividad sospechosa tuya o de alguien con quien compare tu red de Internet. Эспера mientras verificamos que eres una persona real.Tu contenido se mostrará en breve. Si Continúas recibiendo este mensaje, envía un correo electrónico a para informarnos de que tienes problemas.
Hemos estado percibiendo actividad sospechosa de ti o de alguien con quien compare tu red de Internet. Эспера
mientras verificamos que eres una persona real. Tu contenido se mostrará en breve. Si Continúas recibiendo este
mensaje, envía un correo electrónico a
para hacernos saber que
estás teniendo problemas.
Tu contenido se mostrará en breve. Si Continúas recibiendo este
mensaje, envía un correo electrónico a
para hacernos saber que
estás teniendo problemas.
Temos Recebido algumas atividades suspeitas de voiceê ou de alguém que esteja usando a mesma rede.Aguarde enquanto confirmamos que Você é Uma Pessoa de Verdade. Сеу контексто апаресера эм бреве. Caso продолжить Recebendo esta mensagem, envie um email para пункт нет informar sobre o проблема.
Abbiamo notato alcune attività sospette da parte tua o di una persona che condivide la tua rete Internet. Attendi mentre verifichiamo Che sei una persona reale. Il tuo contenuto verrà visualizzato a breve. Secontini Visualizzare questo messaggio, invia un’e-mail all’indirizzo per informarci del проблема.
Пожалуйста, включите куки и перезагрузите страницу.
Это автоматический процесс. Ваш браузер в ближайшее время перенаправит вас на запрошенный контент.
Подождите до 5 секунд…
Перенаправление…
Заводское обозначение: CF-102 / 62055b93ab87900f.
Прямое взаимодействие | Фонд дизайна взаимодействия (IxDF)
Глубоко погрузитесь в прямое взаимодействие с нашим курсом Пользовательский опыт: руководство для начинающих .
Пользовательский опыт, или UX, был модным словом примерно с 2005 года, и, по данным исследовательской компании Gartner, фокус на цифровом опыте больше не ограничивается цифровыми компаниями. Скорее всего, вы слышали об этом термине или даже имеете его в своем портфолио. Но, как и большинство из нас, есть большая вероятность, что вы иногда не уверены в том, что на самом деле охватывает термин «пользовательский опыт».
[Пользовательский опыт] используется людьми, чтобы сказать: ‘ I ’ m дизайнер взаимодействия с пользователем, я разрабатываю веб-сайты ’ или ‘ Я разрабатываю приложения. ’ [ … ], и они думают, что это простое устройство, веб-сайт или приложение, или неизвестно что. Нет! Это ‘ s все — s ‘ s то, как вы воспринимаете мир, это ‘ s то, как вы переживаете свою жизнь, это ‘ s способ вы испытываете сервис.Или, да, приложение или компьютерная система. Но это ’ s система , что ’ s все. ”— Дон Норман, пионер и изобретатель термина « пользовательский опыт », в интервью NNGroup
Как указывает Дон Норман, User Experience — это зонтик термин, охватывающий множество различных областей . Когда вы работаете с пользовательским интерфейсом, очень важно хорошо понимать, что это за области, чтобы знать, какие инструменты вам доступны.
Когда вы работаете с пользовательским интерфейсом, очень важно хорошо понимать, что это за области, чтобы знать, какие инструменты вам доступны.
В ходе этого курса вы, , получите полное представление о различных принципах дизайна, которые вместе создают впечатление пользователя при использовании продукта или услуги . По мере продвижения вы узнаете, какую ценность дизайн пользовательского интерфейса привносит в проект, и какие области следует учитывать, когда вы хотите создать удобный пользовательский интерфейс. Поскольку пользовательский опыт — это развивающийся термин, мы не можем дать вам определение термина «пользовательский опыт», чтобы положить конец всем обсуждениям, но мы дадим вам четкое представление о различных аспектах взаимодействия с пользователем, чтобы оно стало ясным для вас. что нужно для создания отличного UX-дизайна.
Если вы новичок в Interaction Design Foundation, этот курс — отличное место для начала , потому что он объединяет материалы из многих других наших курсов. Это дает вам как отличное введение в пользовательский опыт, так и предварительный просмотр курсов, которые мы можем предложить, чтобы помочь вам в развитии вашей будущей карьеры. После каждого урока мы познакомим вас с курсами, которые вы можете пройти, если ваше внимание привлекла конкретная тема. Так вам будет легко продолжить свое обучение.
Это дает вам как отличное введение в пользовательский опыт, так и предварительный просмотр курсов, которые мы можем предложить, чтобы помочь вам в развитии вашей будущей карьеры. После каждого урока мы познакомим вас с курсами, которые вы можете пройти, если ваше внимание привлекла конкретная тема. Так вам будет легко продолжить свое обучение.
Прямое взаимодействие между белком сборки нуклеосомы 1 и белками папилломавируса E2, участвующими в активации транскрипции
РЕФЕРАТ
Используя дрожжевой двухгибридный скрининг, мы идентифицировали белок сборки нуклеосомы 1 человека (hNAP-1) как белок, взаимодействующий с активацией домен активатора транскрипции, кодируемый папилломавирусами (ПВ), белок Е2. Мы показали, что взаимодействие между E2 и hNAP-1 является прямым, а не просто опосредуется транскрипционным коактиватором p300, который связывается обоими белками.Коэкспрессия hNAP-1 сильно усиливает активацию E2, что также указывает на функциональное взаимодействие. E2 связывается по крайней мере с двумя отдельными доменами в hNAP-1, одним внутри С-конца и внутренним доменом. Связывание E2 с hNAP-1 необходимо для взаимодействия факторов. Более того, 91 N-концевая аминокислота имеет решающее значение для транскрипционной активности hNAP-1, поскольку делеционные мутанты, лишенные этой N-концевой части, неспособны кооперироваться с E2. Мы предоставляем доказательства того, что hNAP-1, E2 и p300 могут образовывать тройной комплекс, эффективный при активации транскрипции.Мы также показываем, что р53 напрямую взаимодействует с hNAP-1, указывая на то, что активаторы транскрипции в дополнение к PV E2 взаимодействуют с hNAP-1. Эти результаты предполагают, что связывание специфичных для последовательности ДНК-связывающих белков с hNAP-1 может быть важным шагом, способствующим активации транскрипции.
E2 связывается по крайней мере с двумя отдельными доменами в hNAP-1, одним внутри С-конца и внутренним доменом. Связывание E2 с hNAP-1 необходимо для взаимодействия факторов. Более того, 91 N-концевая аминокислота имеет решающее значение для транскрипционной активности hNAP-1, поскольку делеционные мутанты, лишенные этой N-концевой части, неспособны кооперироваться с E2. Мы предоставляем доказательства того, что hNAP-1, E2 и p300 могут образовывать тройной комплекс, эффективный при активации транскрипции.Мы также показываем, что р53 напрямую взаимодействует с hNAP-1, указывая на то, что активаторы транскрипции в дополнение к PV E2 взаимодействуют с hNAP-1. Эти результаты предполагают, что связывание специфичных для последовательности ДНК-связывающих белков с hNAP-1 может быть важным шагом, способствующим активации транскрипции.
Активация транскрипции генов, репрессированных нуклеосомами, требует присутствия активаторов, специфически связывающих последовательность. Они повышают эффективность сборки транскрипционно компетентного преинициативного комплекса (PIC) и противодействуют репрессивным эффектам хроматина за счет привлечения комплексов ремоделирования хроматина и гистоновых ацетилтрансфераз (HAT) (2, 15, 30, 31, 40, 52, 53, 64). Ремоделирование хроматина осуществляется с помощью больших АТФ-зависимых ремоделирующих комплексов хроматина, которые изменяют структуру хроматина путем временного нарушения взаимодействий гистон-ДНК (см. Ссылки 6, 30 и 63). Посттрансляционные модификации хроматина, такие как ацетилирование транскрипционными коактиваторами, также вносят вклад в регуляцию генов (22). Считается, что HAT катализируют добавление ацетильных групп к N-концевым хвостам гистонов ядра, процесс, который обычно коррелирует с активацией транскрипции (55).Коактиватор транскрипции p300 и его гомолог CREB-связывающий белок (CBP) обладают HAT-активностью, и оба участвуют в регуляции транскрипции с помощью большого количества последовательностей-специфичных белков-активаторов (см. Ссылки 11, 19 и 21). p300 и CBP связаны с другими HAT, такими как p / CAF, ACTR и SRF, в мультибелковом комплексе. Функциональные исследования показали, что коактиваторная функция p300 и CBP требует их ацетилтрансферазной активности (32, 33, 41).
Ремоделирование хроматина осуществляется с помощью больших АТФ-зависимых ремоделирующих комплексов хроматина, которые изменяют структуру хроматина путем временного нарушения взаимодействий гистон-ДНК (см. Ссылки 6, 30 и 63). Посттрансляционные модификации хроматина, такие как ацетилирование транскрипционными коактиваторами, также вносят вклад в регуляцию генов (22). Считается, что HAT катализируют добавление ацетильных групп к N-концевым хвостам гистонов ядра, процесс, который обычно коррелирует с активацией транскрипции (55).Коактиватор транскрипции p300 и его гомолог CREB-связывающий белок (CBP) обладают HAT-активностью, и оба участвуют в регуляции транскрипции с помощью большого количества последовательностей-специфичных белков-активаторов (см. Ссылки 11, 19 и 21). p300 и CBP связаны с другими HAT, такими как p / CAF, ACTR и SRF, в мультибелковом комплексе. Функциональные исследования показали, что коактиваторная функция p300 и CBP требует их ацетилтрансферазной активности (32, 33, 41). Более того, дополнительные функции этих кофакторов, такие как взаимодействие с компонентами ПОС, необходимы для их стимулирующей активности (3, 45, 51).
Более того, дополнительные функции этих кофакторов, такие как взаимодействие с компонентами ПОС, необходимы для их стимулирующей активности (3, 45, 51).
Сборка нуклеосом тесно связана с репликацией ДНК. Голые дочерние нити вновь реплицированной ДНК быстро собираются в хроматин с помощью многоступенчатого процесса. Фактор сборки хроматина 1 и фактор сборки, связанный с репликацией / белок функции подавления молчания 1 (ASF1) действуют как шапероны гистонов для депонирования гистонов h4 и h5. Белок сборки нуклеосомы (NAP) представляет собой шаперон гистона, ответственный за включение двух димеров гистона h3A-h3B для завершения нуклеосомы (см. Ссылку 62).NAP-1 может действовать как нуклеоцитоплазматический челночный белок, который доставляет димеры h3A-h3B из цитоплазмы в механизм сборки хроматина в ядре (47). Помимо своей функции в сборке хроматина, NAP-1 также может играть роль в развитии клеточного цикла. Генетические эксперименты с дрожжами показали, что NAP-1 играет роль в развитии клеточного цикла во время фазы G 1 и митоза. NAP-1 связывается с циклином B (29) и киназой Gin4p (1).
NAP-1 связывается с циклином B (29) и киназой Gin4p (1).
Более того, гистоновые шапероны, по-видимому, способствуют активации транскрипции за счет своей модифицирующей хроматин активности.Недавние данные свидетельствуют о том, что комплексы HAT, а также АТФ-зависимые комплексы ремоделирования хроматина взаимодействуют с шаперонами гистонов в изменении структуры хроматина во время активации транскрипции. Было обнаружено, что ASF1 функционально взаимодействует с АТФ-зависимым комплексом ремоделирования хроматина Brahma (SWI / SNF), участвующим в активации транскрипции (48). Сообщалось также о функциональном взаимодействии между p300 / CBP и NAP (4, 27, 58). Было продемонстрировано, что ацетилирование гистонов с помощью p300 облегчает перенос гистонов h3A и h3B на NAP-1 in vitro.Таким образом, структура гистонов может быть изменена ацетилированием гистонов, что способствует потере димеров h3A-h3B, которые были реконструированы под действием АТФ-зависимых ремоделирующих комплексов хроматина (27). Эта модель подтверждается наблюдением, что NAP могут усиливать активацию за счет факторов, которые используют p300 в качестве коактиватора (58). Кроме того, было показано, что NAP-1 стимулирует связывание факторов транскрипции с их сайтами связывания, процесс, который сопровождается разрушением октамера гистонов (65).Хотя накапливаются намеки на роль гистоновых шаперонов, таких как NAP-1, в активации транскрипции, неизвестно, каким образом гистоновые шапероны могут быть нацелены на активно транскрибирующие промоторные области.
Эта модель подтверждается наблюдением, что NAP могут усиливать активацию за счет факторов, которые используют p300 в качестве коактиватора (58). Кроме того, было показано, что NAP-1 стимулирует связывание факторов транскрипции с их сайтами связывания, процесс, который сопровождается разрушением октамера гистонов (65).Хотя накапливаются намеки на роль гистоновых шаперонов, таких как NAP-1, в активации транскрипции, неизвестно, каким образом гистоновые шапероны могут быть нацелены на активно транскрибирующие промоторные области.
Белок E2 папилломавируса (PV) представляет собой фактор транскрипции, который регулирует транскрипцию PV, а также необходим для репликации вирусной ДНК (14, 18, 46). Для бычьего PV типа 1 (BPV1) E2 сильно активирует несколько промоторов BPV1, связываясь с его высокоаффинной палиндромной последовательностью распознавания, ACCGN 4 CGGT, присутствующей в нескольких копиях в геноме PV (60).Активация транскрипции вирусными белками E2 включает связывание домена активации (AD) с факторами транскрипции TFIIB и TBP и кофакторами AMF1 / Gps2 и p / CAF. Все эти взаимодействия кажутся важными для активации E2 (7, 9, 36, 44, 70). Кроме того, E2 взаимодействует с p300 / CBP (37, 49, 56). Было показано, что коэкспрессия p300 может усиливать активацию с помощью E2, указывая на то, что связывание с p300 может быть этапом, ограничивающим скорость активации с помощью E2 (49).
Все эти взаимодействия кажутся важными для активации E2 (7, 9, 36, 44, 70). Кроме того, E2 взаимодействует с p300 / CBP (37, 49, 56). Было показано, что коэкспрессия p300 может усиливать активацию с помощью E2, указывая на то, что связывание с p300 может быть этапом, ограничивающим скорость активации с помощью E2 (49).
Здесь мы описываем дрожжевую двугибридную систему, которая использует транскрипционно компетентный белок Е2 BPV1.Эта система позволила идентифицировать человеческий NAP-1 (hNAP-1) как партнера по взаимодействию для E2. Мы представляем данные, демонстрирующие, что прямое взаимодействие между E2 и hNAP-1 важно для активации транскрипции с помощью BPV1 E2. Однако требуются дополнительные функции, расположенные на N-конце hNAP-1. Мы показываем, что hNAP-1 и E2 могут одновременно связываться с p300 и что этот стабильный тройной комплекс эффективно способствует активации транскрипции. Кроме того, р53 также напрямую связывается с hNAP-1.Наши данные предполагают, что связывание с NAP-1 может быть важным шагом для последовательностных активаторов транскрипции, которые используют p300 в качестве коактиватора для активации транскрипции.
МАТЕРИАЛЫ И МЕТОДЫ
Конструкции плазмид. Все дрожжевые плазмиды были челночными векторами, и клонирование выполняли с использованием штамма Escherichia coli XL1Blue. Репортерная конструкция дрожжей, экспрессирующая ген lacZ под контролем синтетического промотора, содержащего четыре сайта связывания E2 перед ТАТА-боксом человеческого PV (HPV) типа 18 (HPV18) P 105 , была получена в несколько этапов.Фрагмент промотора Sal I- Bgl II был выделен из конструкции p4E2 23 T 105 (24), лигирован с олигонуклеотидом, кодирующим соединение между промотором и геном lacZ , за которым следует Bsp. EI сайта рестрикции и клонировали в сайт рестрикции Sal I- Bsp EI pBR322. Впоследствии этот фрагмент промотора был вставлен в сайты рестрикции Xho I- Bsp EI pΔUAS (34), что привело к замещению промотора CYC промотором 4E2 23 P 105 .Наконец, фрагмент промотора Hin dIII- Bam HI, выделенный из этой конструкции, был вставлен во фрагмент вектора Hin dIII- Bam HI placZi (Clontech) с образованием 4E2 23 P 105 lacZi. 5E2-pHIS min был сконструирован путем вставки олигонуклеотидов, кодирующих сайты связывания E2, в сайт связывания Xba I pHIS-i1 (Clontech). Вектор экспрессии дрожжевого E2 pGBT9-E2 был получен путем удаления ДНК-связывающего домена GAL4 (DBD) из pGBT9 (Clontech) путем переваривания с помощью Bam HI и частичного переваривания с помощью Hin dIII.Кодирующий E2 фрагмент Hin dIII- Bam HI был выделен из pTZE2mHIII (34) и клонирован в pGBT9. Фрагмент Hin dIII- Bam HI, полученный из pTZE2Δ192-282 (42), был вставлен аналогичным образом для получения pGBT9-E2Δ192-282.
Для экспрессии AD GAL4, слитого с DBD E2, Stu I- Bam HI-фрагмент E2, полученный из pTZE2mHIII (42), клонировали в вектор pGAD424 (Clontech). Полноразмерный клон hNAP-1 был выделен из библиотеки кДНК HaCaT линии клеток кератиноцитов (экспрессирующих белки, кодируемые кДНК; Clontech) путем амплификации со специфическими праймерами и клонирован в рамке с меткой гемагглютинина (HA) в векторы pXJ41 (68) и pcDNA3.1 (Invitrogen). Плазмиды, экспрессирующие слитый белок глутатион S -трансфераза (GST) — hNAP-1, были получены путем клонирования продуктов ПЦР, кодирующих полноразмерный hNAP-1, в pGEX-5X-2 и pGEX-2T (Pharmacia Biotech) или делеционных мутантов в pGEX. -2Т. Некоторые из делеционных конструкций hNAP-1 были впоследствии перенесены в эукариотический вектор pXJ41 для их экспрессии, слитой с меткой НА. Для экспрессии белков E2 BPV1, HPV8 и HPV18, слитых с тегом FLAG, соответствующие открытые рамки считывания были амплифицированы с помощью ПЦР с последующим клонированием в вектор pCMV2-FLAG (Kodak), как и BPV1 E2 без AD и E2, экспрессирующие AD.Вектор экспрессии pC59, экспрессирующий BPV1 E2 под контролем раннего промотора обезьяньего вируса 40 (SV40), был описан ранее (69). Векторы экспрессии для HPV8 E2 и HPV18 E2 и репортерная конструкция некодирующей области HPV8-люциферазы также были описаны ранее (49), как и вектор экспрессии для p300 (13). Регуляторная область BPV1 от нуклеотидов 6958 до 475 (длинная контрольная область [LCR]) была амплифицирована с помощью ПЦР и клонирована в pALuc (12) для получения BPV1 LCR-Luc. Вектор экспрессии p53 pCp53wt был описан Nigro et al.(54), а синтетическая р53-чувствительная репортерная конструкция была описана Funk et al. (17). Репортерная конструкция pG5-Luc и вектор, экспрессирующий DBD GAL4 (pM), были получены из двугибридной системы млекопитающих (Clontech). Продукты ПЦР, кодирующие AD TEF-1 или E2 HPV8, клонировали в вектор pM.
Дрожжевая двугибридная система. Поддержание дрожжей, трансформацию и хранение выполняли в соответствии с инструкциями производителя дрожжевой двухгибридной системы (Clontech). Штамм дрожжей YM954 (67) (генотип: MAT a ura3-52 his3-200 ade2-101 lys2-801 leu2-3 , 112 trp1-901 gal4-542 gal80-538 ) любезно предоставлены P.Bartel, Stony Brook, N.Y. Активность β-галактозидазы (β-Gal) определяли с помощью анализа подъема фильтра и анализа жидкого β-Gal в соответствии с протоколами от Clontech.
Культура клеток и трансфекция. RTS3b, линия клеток кератиноцитов, полученная из HPV-отрицательного поражения кожи реципиента почечного трансплантата (57), культивировали в среде E (43). Для временной трансфекции использовали 10 5 клеток RTS3b для засева одного шестилуночного планшета за день до трансфекции. Трансфекцию проводили реагентом FuGene в соответствии с рекомендациями производителя (Roche Diagnostics), и клетки собирали через 48 часов.Люциферазную активность определяли, как описано ранее (49). Для создания стабильной клеточной линии, экспрессирующей репортерный ген люциферазы под контролем LCR BPV1, интегрированного в клеточный геном, клетки RTS3b трансфицировали конструкцией LCR-Luc BPV1. Через 48 ч клетки помещали в среду, содержащую 800 мкг G418 / мл. Еще через 48 ч среду заменяли средой, содержащей 400 мкг G418 / мл; эту среду меняли через день в течение 2–3 недель до тех пор, пока устойчивые колонии не выросли.Затем устойчивые колонии объединяли. Присутствие интактной кассеты LCR-Luc BPV1 в клетках подтверждали с помощью ПЦР (данные не показаны). Через 24 часа после трансфекции этой стабильной клеточной линии плазмидами экспрессии E2 или hNAP-1 среду заменяли средой с добавлением 400 мкг трихостатина A (TSA) / мл; инкубацию продолжали еще 24 часа до определения активности люциферазы, как описано выше. Чтобы обнаружить влияние hNAP-1 на активацию экспрессии p21 с помощью p53, клетки RTS3b временно трансфицировали соответствующими векторами экспрессии (см.рис.6). Всего 90 мкг общего клеточного экстракта наносили на гель додецилсульфат натрия-15% полиакриламид и анализировали на p21 вестерн-блоттингом с антителом, направленным против p21 (Pharmingen).
Коиммунопреципитация и анализы «pull-down». Анализы «pull-down» GST проводили, как описано ранее (49). Чтобы обнаружить прямое межбелковое взаимодействие, His-меченные белки экспрессировали в бактериях, очищали, как описано ранее (25), инкубировали с иммобилизованным слитым белком GST-hNAP-1 или GST, а затем обрабатывали, как описано в другом месте (49).Чтобы обнаружить взаимодействие между различными белками E2 и hNAP-1 in vivo, клетки 293T трансфицировали вектором экспрессии для белка E2 BPV1, меченного FLAG (см. Рис. 2E), или векторами для различных белков E2, меченных FLAG (см. Рис. 2D) и для HA-меченного hNAP-1. Через 48 ч клетки промывали ледяным фосфатно-солевым буфером с последующими четырьмя циклами замораживания-оттаивания в буфере LSDB (50 мМ трис-HCl [pH 7,9], 10% глицерин, 0,5 мМ EDTA, 1 мМ дитиотреитол, 0,2% НП-40), содержащий 100 мМ KCl. Клеточный дебрис удаляли центрифугированием в течение 10 мин при 4 ° C.Супернатанты инкубировали с 20 мкл аффинной смолы против FLAG M2 (Kodak). Образцы инкубировали при 4 ° C в течение 2 часов при осторожном перемешивании, затем четыре раза промывали в буфере LSDB с 200 мМ KCl или 250 мМ KCl и одну промывали в буфере LSDB с 100 мМ KCl. Присутствие hNAP-1 анализировали с помощью вестерн-блоттинга с использованием моноклонального антитела, направленного против эпитопа HA (Roche Diagnostics), или моноклонального антитела, направленного против hNAP-1 (любезно предоставленного Y. Ishimi), для обнаружения взаимодействия между E2 и эндогенным hNAP-1.Белки E2 детектировали с помощью антитела против FLAG M5. Чтобы обнаружить взаимодействие между эндогенным p53 и эндогенным hNAP-1 in vivo, 85 мкг ядерных экстрактов из нормальных неонатальных эпидермальных кератиноцитов человека (приобретенных у Clonetics) и 85 мкг ядерных экстрактов из клеток RTS3b инкубировали с 5 мкл анти-p53- конъюгат агарозы (DO-1; Santa Cruz) при 4 ° C в течение 2 ч при осторожном перемешивании, с последующими четырьмя промываниями в буфере LSDB с 200 мМ KCl и одной промывкой в буфере LSDB с 100 мМ KCl. Наличие эндогенных белков анализировали вестерн-блоттингом с антителами, направленными против hNAP-1 и p53 (DO-1).
Анализ градиента плотности тройного комплекса. HNAP-1, E2 с делецией аминокислот 326-420 (E2Δ326-420) и фрагмент p300 из аминокислот с 1195 по 1761 (p300 от 1195 до 1761) были экспрессированы с метка из шести гистидинов в бактериях и очищена. Всего 800 нг p300 от 1195 до 1761, 600 нг hNAP-1 и 200 нг E2Δ326-420 инкубировали в течение 2 часов в присутствии 25 мМ HEPES-KOH (pH 7,9) -0,1 мМ EDTA-12,5 мМ KCl. -10% глицерин-100 мМ KCl в различных комбинациях (см. Рис.5С). Затем белки загружали в 2 мл градиента от 7,5 до 30% глицерина, приготовленного, как описано Tanese et al. (61), а затем центрифугировали в течение 8 ч при 50 000 об / мин с помощью настольной ультрацентрифуги TL100 (Beckman) и ротора с качающимся ковшом TLS-55. Наконец, 36 фракций по 60 мкл каждая были осторожно удалены, начиная с вершины градиентов. Наличие различных белков анализировали с помощью вестерн-блоттинга.
РЕЗУЛЬТАТЫ
Идентификация клеточных факторов, связывающихся с белками PV E2 дрожжевой двугибридной системой.Чтобы идентифицировать клеточные факторы, которые могут играть роль в E2-опосредованной активации транскрипции, мы создали дрожжевую двугибридную систему. Поскольку мы хотели провести скрининг с использованием белка E2 дикого типа, который является сильным активатором транскрипции в дрожжевых клетках (34), мы модифицировали классическую дрожжевую двугибридную систему. Ранее Ham et al. показали, что в клетках млекопитающих E2 не может эффективно активировать транскрипцию минимального промотора, состоящего только из сайтов связывания E2 и TATA-бокса, но является эффективным активатором более сложных промоторов (24).Напротив, большинство белков-активаторов, специфичных к клеточной последовательности, в том числе GAL4, способны стимулировать такой минимальный промотор (66). Чтобы проверить, не сможет ли E2 активировать транскрипцию с этого минимального промотора в дрожжевых клетках, мы вставили фрагмент, экспрессирующий такой минимальный промотор, в вектор placZi (рис. 1). E2 приводил к трех-четырехкратной активации экспрессии гена lacZ из репортерной конструкции 4E2 23 P 105 placZi, интегрированной в клеточный геном.Слитый белок, состоящий из DBD E2 и AD GAL4, приводит к 14-кратной активации (Рис. 1B), указывая на то, что способ активации, опосредованный двумя AD, также различен в дрожжевых клетках. Кроме того, E2 только слабо активировал промотор HIS min , расположенный за пятью сайтами связывания E2 и контролирующий экспрессию гена HIS в плазмиде 5E2-pHIS min (рис. 1A). Следовательно, плазмиду 5E2-pHIS min использовали для отбора дрожжей в присутствии от 100 до 300 мМ 3-амино-1,2,4-триазола, ингибитора продукта гена HIS .Было необходимо включить этот дополнительный отбор для подавления роста дрожжевых клеток, опосредованного слабой активацией pHIS min с помощью E2.
РИС. 1.Идентификация hNAP-1 в качестве партнера по взаимодействию для белка E2 PV. (A) Схематическое изображение двух репортерных конструкций. 5E2-pHIS min экспрессирует дрожжевой ген HIS3 под контролем его минимального промотора, pHIS min и пяти сайтов связывания E2. 4E2 23 P 105 placZi экспрессирует ген lacZ под контролем синтетического промотора, состоящего из ТАТА-бокса раннего промотора HPV18 (P 105 ) и четырех сайтов связывания E2, расположенных на 23 п.н.Для дрожжевой двугибридной системы обе репортерные конструкции были интегрированы в клеточный геном YM954. (B) Вектор экспрессии дрожжей pGBT9-E2, экспрессирующий белок E2 BPV1 (E2), или pGBT9-E2DBD-GAL4AD (E2DBD-GAL4-AD), экспрессирующий AD GAL4, слитого с DBD BPV1 E2, или вектор один (-) трансформировали в дрожжевой штамм, несущий две репортерные конструкции, показанные на панели A. Определяли активность β-Gal соответствующих штаммов. Активности в присутствии одного вектора (-) было присвоено произвольное значение 1, и была рассчитана кратность активации.Столбцы представляют собой средние значения трех независимых экспериментов, выполненных с двумя разными клонами в каждом случае; показаны планки погрешностей. (C) активности β-Gal родительского штамма дрожжей YM954, трансформированного дрожжевым экспрессирующим вектором для E2 или E2Δ195-282 (белок E2, лишенный внутренней шарнирной области) и плазмидой pACT-c-ΔNAP-1, экспрессирующей слитый hNAP-1 к AD GAL4, были определены. Последний был выделен двугибридным дрожжевым скринингом. Активацию сворачивания по отношению к активации штамма, трансформированного вектором экспрессии, рассчитывали, как описано для панели B.Столбцы представляют собой средние значения для трех независимых клонов; показаны планки погрешностей.
Библиотека кДНК, полученная из линии клеток кератиноцитов HaCaT и обеспечивающая экспрессию кодируемых кДНК белков, слитых с GAL4 AD, была трансформирована в штамм дрожжей, несущий два репортерных гена в дополнение к вектору экспрессии pGBT9-E2. Из 10 7 клонов дрожжей два репортерных гена были значительно активированы в 41 клоне. Белки, кодируемые кДНК, двух независимых клонов были идентичны 300 C-концевым аминокислотам hNAP-1, который в целом содержит 391 аминокислоту.Взаимодействие между hNAP-1 и E2 можно было подтвердить с помощью штаммов дрожжей, которые были повторно трансформированы соответствующими плазмидами (фиг. 1C). Во взаимодействии участвовал N-концевой AD BPV1 E2, поскольку C-концевые 94 аминокислоты BPV1 E2, содержащие DBD, не выявляли повышенной активности β-Gal в присутствии hNAP-1 (данные не показаны). Однако E2Δ195-282, у которого отсутствует внутренняя шарнирная область, сохранил аналогичную кооперативность с hNAP-1 (Fig. 1C). Пониженные абсолютные уровни активации этого делеционного мутанта коррелировали со сниженными уровнями белка (данные не показаны).
белков E2 связываются с hNAP-1 in vitro и in vivo. Чтобы подтвердить взаимодействие между E2 и h-NAP-1 in vitro, фрагмент hNAP-1, выделенный двухгибридной системой дрожжей и кодирующий hNAP-1, лишенный N-концевая 91 аминокислота была вставлена в вектор pGEX-5X-2. Слитый белок GST-hNAP-1 сохранял полноразмерный белок E2 BPV1, полученный трансляцией in vitro с лизатом ретикулоцитов кролика (фиг. 2A). Белки E2 разных типов PV довольно консервативны в N-концевой AD и C-концевой DBD, в отличие от вариабельной шарнирной области, которая, как предполагалось, функционирует как линкер между AD и DBD E2 (20, 42).Транслируемые in vitro белки E2 HPV8 и HPV18 связываются с GST-hNAP-1 (фиг. 2A), что указывает на то, что взаимодействие с hNAP-1 может быть консервативным среди белков E2 различных типов PV.
РИС. 2.белков E2 и hNAP-1 взаимодействуют in vitro. (A) Анализы GST pull-down проводили с очищенным GST, полноразмерным hNAP-1, слитым с GST (GST-hNAP-1), и 35 S-меченых белков E2 BPV1, HPV8 и HPV18, полученных путем трансляции in vitro с лизатом ретикулоцитов кролика. Десять процентов ввода показаны на дорожках 1, 4 и 7.Указаны положения маркерных белков. (B) (Верхняя панель) 35 S-меченный полноразмерный E2 BPV1 (E2), два мутанта с N-концевой делецией с дефектом активации (E2Δ1-161 и E2Δ1-203) и мутант с C-концевой делецией без DBD (E2Δ326-420) инкубировали с GST, GST-hNAP-1 или, в качестве отрицательного контроля, GST-p300-2, экспрессирующим фрагмент p300, который, как было показано ранее, не взаимодействует с E2 (49). Десять процентов белка E2 и его производных, использованных в одном анализе взаимодействия, показаны на дорожках, обозначенных как 10% вход.(Средняя панель) Белки E2 с точечными мутациями в AD, E2 E39A, репликация недостаточна, и E2 I73A, транскрипция которой нарушена (14), были использованы в анализе GST pull-down. (Нижняя панель) Структура белка E2, показывающая положения аминокислот (аа) в различных доменах. (C) His-меченный, бактериально экспрессированный, очищенный белок E2 BPV1 (His-E2) или His-меченный E2Δ326-420 инкубировали с очищенным GST или GST-hNAP-1. Связанные белки E2 детектировали вестерн-блоттингом с антителом, направленным против метки His (Qiagen).Указано положение маркерного белка массой 31 кДа. (D) (Верхняя панель) Клетки 293T трансфицировали экспрессирующим вектором, экспрессирующим FLAG-меченный полноразмерный белок E2 BPV1 (дорожки 1, 3, 5, 7, 10 и 14), мутант E2 BPV1 с тегом FLAG. без N-концевой AD (FLAG-E2Δ1-203; дорожки 12 и 16) или AD с тегом FLAG (FLAG-E2Δ204-420; дорожки 11 и 15) и вектора экспрессии для HA-тегированного hNAP-1 (дорожки 2 , 3, 6, 7 и с 9 по 16). Экстракты клеток инкубировали с аффинным гелем FLAG, и связанный hNAP-1 детектировали с помощью антитела, направленного против эпитопа НА (дорожки 1-4 и дорожки 9-12).В дорожки с 5 по 8 и с 13 по 16 было включено 1/40 введенного клеточного экстракта, и экспрессия HA-меченого hNAP-1 анализировалась с помощью антитела HA. Присутствие белков Е2, меченных FLAG, определяли повторным зондированием блота, показанного на дорожках 1-4 и дорожках 10-12, с антителом FLAG M5. IP, иммунопреципитация; WB, Вестерн-блоттинг. (Нижняя панель) Вестерн-блоттинг, разработанный с использованием антитела, направленного против эпитопа HA, при коиммунопреципитации, аналогичной показанной выше, но с клеточными экстрактами из клеток 293T, которые были трансфецированы вектором экспрессии для FLAG-меченного HPV18 E2 (дорожки 17, 19, 22 и 24) или Е2 HPV8 (дорожки 18, 20, 23 и 25), или вектор экспрессии для HA-tagged hNAP-1 (дорожки 17, 18, 21, 22, 23 и 26).(E) (Верхняя панель) Клеточные экстракты клеток 293T, которые были трансфицированы вектором экспрессии для FLAG-меченого BPV1 E2 (дорожки 3 и 6) или FLAG-меченой бактериальной щелочной фосфатазы (BAP) в качестве отрицательного контроля (дорожки 2 и 5) или с пустым вектором (дорожки 1 и 4) инкубировали с антителом FLAG, связанным с сефарозой (дорожки 4–6). Связанный эндогенный hNAP-1 обнаруживали вестерн-блоттингом с антителом, направленным против hNAP-1. В дорожках 1-3, 1/160 клеточный экстракт, используемый для иммунопреципитации, загружали в качестве входного контроля.(Нижняя панель) Часть блота на верхней панели была повторно зондирована антителом FLAG для обнаружения присутствия белков, меченных FLAG. Клеточный белок, перекрестно реагирующий с обоими антителами, отмечен звездочкой.
В корреляции с данными, полученными на дрожжевых клетках, AD BPV1 E2 требуется для связывания с hNAP-1 in vitro. E2Δ1-161, в котором отсутствуют N-концевые 161 аминокислоты и который представляет собой встречающийся в природе мутант с усеченным N-концом, который подавляет активацию полноразмерным белком E2 (35), все еще взаимодействует с hNAP-1; напротив, E2Δ1-203, в котором отсутствует вся AD, вообще не связывался.Белок E2, у которого отсутствует C-концевой DBD, E2Δ386-420, сохранил связывание, как показано на фиг. 2B. Различные белки E2 в среднем на 30% идентичны аминокислотам в AD. Многие консервативные остатки по всей AD важны как для репликации, так и для транскрипции. Однако две аминокислотные замены четко разделяют эти две емкости. Замена Ile в положении 73 на Ala (I73A) разрушает активацию транскрипции, оставляя функцию репликации интактной, тогда как замена Glu в положении 39 на Ala (E39A) имеет обратный фенотип (14).Ни одна из аминокислотных замен не влияла на связывание BPV1 E2 с GST-hNAP-1, как показано на фиг. 2B.
Чтобы исключить возможность того, что взаимодействие между BPV1 E2 и hNAP-1 опосредуется фактором, присутствующим в лизате ретикулоцитов или в дрожжевых клетках, мы инкубировали GST-hNAP-1 с His-меченным, бактериально экспрессируемым, очищенным полным -длина БПВ1 Е2 или Е2Δ386-420. Как показано на фиг. 2C, как E2, так и E2Δ386-420 специфически удерживаются GST-hNAP-1, указывая на то, что взаимодействие является прямым.В этом эксперименте связывание полноразмерного E2 было более слабым, чем связывание E2Δ386-420. Этот результат мог быть связан с более низкой концентрацией полноразмерного E2. Нам не удалось очистить полноразмерный E2 в тех же количествах, что и E2Δ386-420. Более того, более быстро мигрирующий продукт, который, по-видимому, представляет собой N-концевую протеолитическую форму, поскольку он сохраняет His-метку, продемонстрировал более сильное взаимодействие с hNAP-1, подтверждая представление о том, что взаимодействие опосредуется AD (рис. 2C). ).
E2 и hNAP-1 также взаимодействуют внутри клетки, как показано на рис.2D. Клетки 293T временно трансфицировали векторами экспрессии для FLAG-меченого полноразмерного BPV1 E2, белка E2, в котором отсутствует N-концевой AD (FLAG-E2Δ1-203) или AD BPV1 E2 (FLAG-E2Δ204-420) и вектор экспрессии для HA-меченного полноразмерного hNAP-1. В то время как в присутствии полноразмерного E2 BPV1 или его AD, HA-меченный hNAP-1 мог быть соосажден антителом FLAG, E2Δ1-203 не опосредовал этот эффект, подтверждая, что взаимодействие внутри клетки опосредуется AD BPV1 E2 тоже.Белки E2 HPV8 и HPV18 также были способны совместно преципитировать меченный HA hNAP-1 (фиг. 2D).
Кроме того, E2 связывается с эндогенным hNAP-1. Во всех линиях клеток карциномы шейки матки, инфицированных PV, ДНК HPV интегрирована в клеточный геном, что приводит к нарушению открытой рамки считывания для E2. Следовательно, клеточных линий, экспрессирующих E2 на физиологических уровнях во время инфекции HPV, не существует, и для анализа связывания E2 с эндогенным hNAP-1 мы сверхэкспрессировали E2, меченный FLAG, в клетках 293T.Как показано на фиг. 2E, антитело FLAG было способно осаждать эндогенный hNAP-1 из экстрактов клеток, трансфицированных вектором для FLAG-меченого BPV1 E2, а не из экстрактов клеток, трансфицированных пустым вектором экспрессии или вектором, экспрессирующим контрольный белок, бактериальная щелочная фосфатаза, слитая с эпитопом FLAG.
В совокупности данные, представленные на рис. 1 и 2, демонстрируют, что белки E2 из разных типов PV взаимодействуют с hNAP-1. Для BPV1 E2 мы можем продемонстрировать, что это взаимодействие является прямым и включает AD.
E2 и hNAP-1 взаимодействуют в активации экспрессии гена BPV1. Как уже упоминалось, NAP-1 и NAP-2 действуют как гистоновые шапероны, которым приписываются функции как в транскрипции, так и в репликации ДНК (4, 26-28, 39, 50, 58). Здесь мы сосредоточились на участии взаимодействия E2 с hNAP-1, наблюдаемого in vitro, в активации транскрипции. В BPV1 E2 сильно активирует транскрипцию с нескольких промоторов BPV1, связываясь с 12 сайтами связывания E2, расположенными в регуляторной области, называемой LCR (60).Репортерную конструкцию, содержащую полный LCR BPV1 перед геном люциферазы, котрансфицировали в линию клеток человека RTS3b (57), полученную из кератиноцитов человека, естественных клеток-мишеней PV, вместе с векторами для HA-меченного hNAP-1 и BPV1 E2. . Как показано на фиг. 3A, E2 может активироваться от 100 до 130 раз, в зависимости от количества трансфицированного вектора экспрессии. Экспрессия hNAP-1 на собственной стимулированной промоторной активности в 2 раза; однако совместная экспрессия hNAP-1 и небольших количеств E2 увеличивала активность люциферазы в 500 раз.Эта кооперативность между hNAP-1 и BPV1 E2 снижалась в присутствии большего количества E2. Однако при этих высоких концентрациях E2 кооперативность между E2 и hNAP-1 все еще наблюдалась.
РИС. 3.E2 и hNAP-1 взаимодействуют в активации экспрессии гена. (A) (Верхняя панель) Клетки RTS3b, иммортализованные клетки кератиноцитов кожи (57), трансфицировали люциферазной репортерной конструкцией, содержащей регуляторную область BPV1, называемую LCR, структура которой показана. Указаны различные промоторы, а 12 сайтов связывания E2 изображены черными прямоугольниками.(Средняя панель) 5 или 20 нг вектора экспрессии для BPV1 E2 под контролем промотора SV40 котрансфицировали вместе с 400 нг вектора экспрессии для HA-меченного hNAP-1. На графике показаны результаты одного репрезентативного эксперимента. (Нижняя панель) Эксперименты по временной трансфекции с репортерной плазмидой BPV1 LCR-Luc, вектором экспрессии для HA-меченного hNAP-1 и векторами экспрессии для полноразмерного E2 (E2), E2 E39A (мутант с дефицитом репликации), E2 I73A (мутант с нарушенной транскрипцией), E2Δ1-203 (без AD) и E2Δ195-282 (без внутренней шарнирной области).Активность каждого из белков Е2 в отсутствие коэкспрессированного HA-меченного hNAP-1 произвольно определяли как 1. Рассчитывали кратное увеличение E2-опосредованной активации hNAP-1. На графике представлены средние значения по крайней мере трех независимых экспериментов; показаны планки погрешностей. Активация каждого мутантного белка E2 в отсутствие коэкспрессированного hNAP-1 по сравнению с активацией белка дикого типа, которая была установлена на уровне 100%, представлена под графиком. (B) Клетки RTS3b, содержащие репортерную конструкцию BPV1 LCR-Luc, интегрированную в клеточный геном, котрансфицировали векторами экспрессии для HA-меченного hNAP-1 и для E2.Через 24 часа после трансфекции к среде добавляли TSA еще на 24 часа.
Как показано на фиг. 2B, точечные мутации, отменяющие репликационную или транскрипционную функцию E2, не влияли на связывание с hNAP-1 in vitro. В наших экспериментах мутант I73A E2 с дефицитом активации сохранил 33% активационной способности E2 дикого типа (данные не показаны), что хорошо согласуется с предыдущими результатами (14). Однако даже эта остаточная активация стимулировалась примерно в пять раз после котрансфекции hNAP-1, указывая на то, что этот мутант не влияет на стадию связывания с hNAP-1.То же самое верно и для мутанта E2 E39A с дефицитом репликации (рис. 3A). Как и ожидалось, активация транскрипции с помощью E2 необходима для действия hNAP-1, поскольку мутант, лишенный AD, E2Δ1-203, не стимулировался коэкспрессией hNAP-1, в отличие от мутанта, лишенного внутренней шарнирной области, Е2Δ195-282 (рис. 3А). Активность раннего промотора SV40, который управлял экспрессией BPV1 E2 в предыдущих экспериментах по временной трансфекции, не стимулировалась совместной экспрессией количеств E2 и hNAP-1, используемых здесь (данные не показаны).Этот результат исключает возможность того, что усиливающий эффект обусловлен повышением экспрессии E2 с помощью hNAP-1. Эти результаты демонстрируют, что E2 и hNAP-1 взаимодействуют при активации транскрипции, что коррелирует с прямым взаимодействием двух белков.
Поскольку роль NAP-1 в качестве шаперона гистонов известна, возможно, что hNAP-1, в сотрудничестве с E2, также влияет на состояние хроматина во время активации E2. До сих пор мы использовали E2-чувствительную репортерную конструкцию, которая была временно трансфицирована.Чтобы проверить влияние hNAP-1 на активацию E2 на репортерных конструкциях, организованных в структуру хроматина более высокого порядка, мы установили клеточную линию, несущую репортерную конструкцию BPV1 LCR-Luc, интегрированную в клеточный геном. После временной трансфекции этой стабильной клеточной линии вектором экспрессии для E2 мы не смогли наблюдать никакой активации, независимо от того, котрансфицирован ли вектор hNAP-1 или нет (данные не показаны). Поскольку мы подозревали, что плотная упаковка хроматина может препятствовать доступу E2 к ДНК, мы обработали клетки через 24 часа после трансфекции TSA, ингибитором гистондеацетилаз.Эта обработка позволила E2 слабо стимулировать активность люциферазы; этот эффект был немного усилен коэкспрессией hNAP-1 (рис. 3B). Эти результаты демонстрируют, что с промоторами, организованными в клеточном хроматине, hNAP-1 также стимулирует активацию E2, хотя и в гораздо меньшей степени, чем временно трансфицированные репортерные конструкции.
Полноразмерный hNAP-1 необходим для сотрудничества с E2. Чтобы понять механизм кооперативности между белками hNAP-1 и E2, мы определили домены в hNAP-1, которые связаны с E2, и проанализировали их роли в усилении активации E2.Ряд мутантов hNAP-1 с N- и C-концевой делецией экспрессировали как слитые белки GST. Хотя N-концевой участок от остатков 1 до 162 обладал маргинальной связывающей активностью (GST-hNAP-1Δ4, GST-hNAP-1Δ7 и GST-hNAP-1Δ10), внутренний домен от остатков 162 до 290 (GST-hNAP-1Δ6 ), а С-концевой участок от остатков 291 до 392 (GST-NAP-1Δ5) с большей эффективностью связывается с 35 S-меченым BPV1 E2 (фиг. 4A). Эти результаты предполагают, что E2 связывается по крайней мере с двумя разделяемыми доменами в hNAP-1, одним внутренним от аминокислот с 162 по 290 и одним С-концом от остатков с 291 по 392.
РИС. 4.Связывание E2 с hNAP-1 необходимо, но недостаточно для стимуляции E2-опосредованной активации. (A) Различные области hNAP-1 были слиты с GST. Бактериально экспрессируемые очищенные слитые белки GST-hNAP-1 (показаны в геле додецилсульфата [SDS] на дне, окрашенном кумасси синим) инкубировали с 35 S-меченным BPV1 E2. Связанные белки выявляли авторадиографией. Обобщено связывание E2 с hNAP-1 (-, нет связывания; +/-, слабое связывание; +, связывание; ++, сильное связывание).В некоторых случаях присутствие слитых белков GST-hNAP-1 в геле могло незначительно влиять на миграцию белка E2. аа, аминокислоты; эт, полная длина. (B) (Верхняя левая панель) Клетки RTS3b котрансфицировали репортерной конструкцией BPV1 LCR и вектором экспрессии для HA-меченых hNAP-1 или HA-меченных делеционных мутантов hNAP-1, отдельно или вместе с 5 нг E2. вектор выражения. (Верхняя правая панель) Влияние различных мутантов hNAP-1 на активность промотора BPV1 в отсутствие E2.Активность LCR BPV1 в отсутствие hNAP-1 была произвольно определена как 1. (Нижняя правая панель) Влияние мутантов hNAP-1 на активацию E2. Здесь активность в присутствии E2 без hNAP-1 (полоса -) была установлена на 1, и были рассчитаны эффекты различных мутантов hNAP-1. Оба графика представляют собой средние значения по крайней мере трех независимых экспериментов с двумя разными препаратами плазмидной ДНК; показаны планки погрешностей. (Нижняя левая панель) Уровни экспрессии различных HA-меченых делеционных мутантов hNAP-1, проанализированные с помощью вестерн-блоттинга (WB) с экстрактами клеток, которые были временно трансфицированы соответствующими векторами экспрессии и высокоаффинным антителом против эпитопа HA .Звездочки указывают положения соответствующих мутантов hNAP-1. Дорожки с 1 по 9 — 10% полиакриламидный гель; дорожки с 10 по 12, 15% полиакриламидный гель.
Для анализа вкладов доменов hNAP-1, которые взаимодействуют с E2, в стимуляцию активации E2, делеционные мутанты hNAP-1 (фиг. 4B) были слиты по N-концу с тегом HA и экспрессированы. Результаты, полученные в экспериментах по котрансфекции, показанные на фиг. 4B, демонстрируют, что 91 N-концевой остаток необходимы для hNAP-1 для усиления активации E2.Все мутанты hNAP-1, лишенные N-концевой 91 аминокислоты, не могли взаимодействовать с E2 при активации. Большинство мутантов подавляли активацию E2 от 2-кратной (hNAP-1Δ8) до 10-кратной (hNAP-1Δ1, hNAP-1Δ2, hNAP-1Δ5 и hNAP-1Δ7). Эта репрессия не связана с подавлением из-за нефизиологически высоких концентраций этих мутантов по сравнению с концентрацией полноразмерного hNAP-1 в трансфицированных клетках, поскольку все производные hNAP-1 присутствовали в аналогичных количествах. Единственным исключением является мутант hNAP-1Δ8, который продуцировался на пониженных уровнях, как показано на Вестерн-блоте на фиг.4Б. Кроме того, титрование этих белков выявило постоянное дозозависимое подавление (данные не показаны). Мутантный hNAP-1Δ4, сохраняющий N-концевой домен, но лишенный взаимодействия с E2, не влиял на активацию E2. Мутанты hNAP-1Δ3 и hNAP-1Δ9, содержащие N-концевой домен и один E2-связывающий мотив, соответственно, были способны усиливать активацию E2, хотя и в меньшей степени по сравнению с таковой, полученной с hNAP-1 дикого типа. Эти результаты показывают, что в дополнение к N-концевой 91 аминокислоте, по крайней мере, один E2-связывающий мотив необходим для hNAP-1, чтобы стимулировать активацию E2, хотя кажется, что аминокислоты 162-290 hNAP-1 более важны.
Различные мутанты hNAP-1 также модулировали активность промоторов BPV1 в отсутствие E2. Однако не было четкой корреляции между эффектами каждого мутанта на активацию E2 и на базальную активность промотора. Например, hNAP-1Δ5 и hNAP-1Δ9 оба слегка снижали активность базального промотора, но hNAP-1Δ5 сильно подавлял активацию E2, а hNAP-1Δ9 все еще слабо стимулировал активацию E2. Эти результаты предполагают, что эффекты hNAP-1 могут быть специфичными для E2. Более того, эти данные подразумевают, что hNAP-1 выполняет другую функцию в транскрипции, поскольку некоторые из мутантов, например.g., hNAP-1Δ7 и hNAP-1Δ10, которые не взаимодействуют с E2 in vitro, сильно ингибируют активацию E2; последний результат может быть связан с их ингибирующим действием на активность промотора как таковую.
E2, hNAP-1 и p300 могут образовывать тройной комплекс in vitro. Как уже упоминалось, было показано, что NAP-1 и NAP-2 взаимодействуют с p300. Как и в случае E2, связывание p300 с hNAP-1 может включать, по крайней мере, два домена, один от аминокислот 123 до 230, а второй — с С-конца до аминокислоты 290 (4, 58).Таким образом, области в hNAP-1, необходимые для активации E2, также включают домены взаимодействия p300, и мы не можем исключить возможность того, что связывание hNAP-1 с p300 также может быть необходимым. Поскольку было продемонстрировано, что белки E2 HPV8, HPV18 и BPV1 напрямую и функционально взаимодействуют с p300 (37, 49, 56), как и hNAP-1, нас интересовало влияние hNAP-1 и p300 на E2- опосредованная активация. К сожалению, коэкспрессия p300 с репортерной конструкцией BPV1 LCR-Luc, используемой в экспериментах, описанных до сих пор, привела к ингибированию активности промотора (данные не показаны).Однако Müller et al. ранее показали, что Е2 HPV8 и коэкспрессируемый p300 взаимодействуют в активации экспрессии гена HPV8 (49). Поскольку мы продемонстрировали здесь, что в дополнение к BPV1 E2, HPV8 E2 также связывается с hNAP-1 (рис. 2A и D), а коэкспрессируемый hNAP-1 усиливает активацию E2 HPV8 (данные не показаны; см. Рис. 7), мы использовали Система HPV8 для изучения эффектов hNAP-1 и p300 на E2-опосредованную активацию. В соответствии с предыдущими наблюдениями, E2 и p300 взаимодействуют, поскольку они стимулируют активность промотора 15.В 5 раз по сравнению с 4 раз для каждого из белков по отдельности (фиг. 5A). Как было показано ранее, сверхэкспрессия p300 не влияет на экспрессию белка E2 (49). hNAP-1 увеличивал активацию E2 или p300 до 23 или 17 раз соответственно. Таким образом, hNAP-1 может функционально взаимодействовать с обоими белками в корреляции с прямым связыванием hNAP-1 с E2 (рис. 2) и p300 (4, 58). Сверхэкспрессия всех трех белков вместе приводила к 134-кратной активации. Эта активация в присутствии E2, hNAP-1 и p300 была далеко за пределами суммы эффектов отдельных компонентов, предполагая, что тройной комплекс E2, p300 и hNAP-1, вероятно, очень эффективно способствует активации транскрипции. .
РИС. 5.hNAP-1 и E2 могут образовывать тройной комплекс с p300. (A) (Верхняя панель) Репортерную конструкцию, экспрессирующую ген люциферазы под контролем регуляторной области HPV8, котрансфицировали векторами экспрессии для HPV8 E2, p300 и HA-tagged hNAP-1, либо отдельно, либо в комбинации. Активность люциферазы в присутствии пустого вектора экспрессии была установлена на 1, и была рассчитана кратность активации любого из белков. На графике представлены средние значения пяти независимых экспериментов; показаны планки погрешностей.(Нижняя панель) Структура регуляторной области HPV8, включая два промотора и сайты связывания E2 (черные ящики). (B) GST или GST-p300-4 инкубировали с увеличивающимся количеством His-меченного, бактериально экспрессированного, очищенного hNAP-1 в течение 2 часов при 4 ° C. После нескольких промывок для удаления несвязанного hNAP-1, меченного His, следовала инкубация с 35 S-меченным BPV1 E2 или hNAP-1 в течение еще 2 часов. Связывание радиоактивно меченного E2 или hNAP-1 анализировали с помощью авторадиографии. Радиоактивные сигналы были количественно определены с помощью PhosphorImager; были рассчитаны проценты, связанные с GST-p300-4.Десять процентов ввода 35 S-меченного E2 или 20% ввода 35 S-меченного hNAP-1 были загружены в качестве входного контроля. (Нижняя панель) Вестерн-блоттинг (WB) выявляет связывание hNAP-1 с GST-p300-4 в отсутствие (дорожки 22 и 23) или в присутствии (дорожки 26 и 27) 35 S-меченного BPV1 E2. (C) Седиментационный анализ плотности глицерина, выполненный, как описано ранее (61), с 600 нг His-меченного hNAP-1, 200 нг His-меченного E2Δ326-420 (E2) и 800 нг p300 от 1195 до 1761.Образцы подвергали центрифугированию в градиенте глицерина от 7,5 до 30%, и всего было собрано 36 фракций, начиная с вершины градиента. Поскольку первоначальный анализ показал, что все белки фракционированы во фракции с 10 по 31, только эти фракции показаны здесь после анализа вестерн-блоттингом с антителом против His-метки. Присутствие E2, hNAP-1 и p300 в различных фракциях указано справа.
Для анализа возможного образования тройных комплексов среди трех белков in vitro мы использовали фрагмент p300 из аминокислот 1453–1882, слитый с GST (GST-p300-4).Мюллер и др. ранее наблюдали, что E2 связывается с этим сегментом p300, который совместно с доменом C / h4 (49). В дополнение к домену KIX (4), NAP-2 и NAP-1, как было показано, нуждаются в аминокислотах с 1572 по 1818 для связывания с p300 (58), как и E2. Чтобы подтвердить образование тройного комплекса in vitro, мы провели конкурентный эксперимент. Как показано на фиг. 5B, GST или слитый белок GST-p300-4 инкубировали с возрастающими количествами экспрессируемого бактериями очищенного His-меченного hNAP-1 в течение 2 часов.После нескольких стадий промывки для удаления несвязанного hNAP-1, меченного His, добавляли либо 35 S-меченный E2, либо hNAP-1, каждый из которых был получен трансляцией in vitro; после инкубации в течение еще 2 ч связывание анализировали. Сигналы количественно оценивались с помощью PhosphorImager. В то время как в отсутствие немеченого hNAP-1, меченного His, 59% радиоактивно меченного hNAP-1 осаждали GST-p300-4, предварительная инкубация возрастающих количеств немеченого hNAP-1 снижала связывание меченого hNAP-1 до 19% (рис. 5B, дорожки с 6 по 10).Эта конкуренция согласуется с представлением о том, что сайты связывания hNAP-1 в p300-4 были заняты немеченым белком. Напротив, предварительная инкубация GST-p300-4 с такими же возрастающими количествами His-меченного hNAP-1 не влияла на связывание радиоактивно меченного E2. В общей сложности от 17 до 20% введенных белков E2 осаждали с помощью GST-p300-4, независимо от предварительной инкубации с увеличивающимся количеством His-меченного hNAP-1 (фиг. 5B, дорожки с 1 по 5). Как и ожидалось, очищенный hNAP-1, меченный His, также связывался с GST-p300-4 в присутствии E2, как было выявлено вестерн-блоттингом с моноклональным антителом против hNAP-1 (фиг.5B, дорожки с 21 по 27). Эти результаты предполагают, что E2 все еще может взаимодействовать с p300, когда hNAP-1 связан, и что вероятен тройной комплекс между E2, p300 и hNAP-1.
Для дальнейшего изучения образования тройного комплекса между E2, p300 и hNAP-1 in vitro мы исследовали фракционирование белка E2 в присутствии hNAP-1 и p300 в градиенте плотности. После инкубации His-меченного, бактериально экспрессированного, очищенного E2Δ326-420 с His-меченным hNAP-1 и p300 1195-1761 реакцию связывания подвергали центрифугированию в градиенте глицерина, как описано Tanese (61).Всего было собрано 36 фракций, и присутствие E2Δ326-420, hNAP-1 и p300 1195-1761 контролировалось вестерн-блоттингом. Поскольку все три белка были легко различимы из-за их характерных размеров, мы использовали антитело, направленное против His-метки. Чтобы гарантировать, что большая часть E2 образована комплексом с p300 и hNAP-1, мы добавили E2 в ограниченных количествах для реакции связывания. Первоначальный анализ показал, что все три белка осаждались около середины градиента во фракциях с 10 по 31.Следовательно, только эти фракции использовались для подробного анализа, показанного на фиг. 5C. Большая часть E2 присутствовала во фракциях с 22 по 24. После инкубации с hNAP-1, E2 оседал больше в верхней части градиента, с самыми высокими концентрациями во фракциях с 19 по 22 (рис. 5C, панель I, E2 против E2 +. hNAP-1). Таким образом, взаимодействие между E2 и hNAP-1 может быть продемонстрировано в этом анализе градиента глицерина, на что указывает наблюдение, что hNAP-1 кофракционируется (фиг. 5C, панель II, hNAP-1 по сравнению с hNAP-1 + E2).В отличие от добавления hNAP-1, добавление p300 к E2 привело к тому, что E2 оседал больше в направлении нижней части градиента (фиг. 5C, панель I, E2 + p300). Кроме того, E2 софракционировали с p300, который обнаруживался в более высоких фракциях после инкубации с E2, по сравнению со свободным p300, подтверждая взаимодействие между E2 и p300 (рис. 5C, сравните панель I, E2 + p300, с панелью III, p300 + E2). В присутствии всех трех белков E2 снова смещался к вершине градиента (рис.5C, панель I, E2 + hNAP-1 + p300). Однако паттерн E2 отличался от паттерна, полученного после инкубации только с hNAP-1, поскольку теперь большая часть E2 обнаруживалась во фракциях 13-16, что сопровождалось уменьшением фракций 24 и 25. Значительные количества p300 и hNAP-1 также были обнаружены во фракциях с 13 по 22, когда все три белка были инкубированы (фиг. 5C, панели II и III, все три белка). Таким образом, совместное фракционирование E2, hNAP-1 и p300 и изменение седиментации E2 после одновременной инкубации с hNAP-1 и p300 коррелируют с образованием тройного комплекса между E2, hNAP-1 и p300.
В совокупности эксперименты по временной трансфекции, GST-pull-down и седиментации в градиенте плотности показали, что множественные белок-белковые взаимодействия (E2-p300, E2-hNAP-1 и hNAP-1-p300) могут способствовать образованию стабильного тройной комплекс, эффективный при активации транскрипции.
Прямое взаимодействие между p53 и hNAP-1 также участвует в активации, опосредованной p53. Ранее предполагалось, что NAP-2, который тесно связан с NAP-1, увеличивает транскрипцию с помощью активаторов, таких как p53 и E2F, которые использовать p300 в качестве коактиватора путем прямого взаимодействия с p300 (58).Чтобы проверить, может ли быть вовлечено, помимо связывания p300-hNAP-1, прямое взаимодействие между p53 и hNAP-1 (например, для PV E2), мы инкубировали His-меченный, бактериально экспрессированный, очищенный p53 с GST. -hNAP-1 слитый белок. Как показано на фиг. 6A, р53 был осажден слитым белком GST-hNAP-1, а не GST. Взаимодействие можно подтвердить с помощью GST-p53 и очищенного hNAP-1, меченного His (данные не показаны). Эти результаты демонстрируют, что взаимодействие между hNAP-1 и p53 является прямым, а не просто опосредовано другими факторами, такими как p300, как предполагалось в другом месте (58).Когда синтетическая репортерная конструкция, содержащая один сайт связывания p53 перед промотором Hsp70, была временно трансфицирована в p53-отрицательную клеточную линию RTS3b, hNAP-1 также усиливал активацию p53 (фиг. 6B), подтверждая, что взаимодействие является функциональным, как и показан для НАП-2 (58). На основании этих результатов мы подозревали, что hNAP-1 также способен стимулировать активацию p53 эндогенного p53-чувствительного промотора в отсутствие коэкспрессии p300. Мы проанализировали влияние коэкспрессии p53 и hNAP-1 на уровень p21 в клеточной линии RTS3b.В условиях вестерн-блоттинга, показанных на фиг. 6C, эндогенный p21 не был виден. Котрансфекция вектора экспрессии p53 или небольших количеств вектора экспрессии hNAP-1 не вызывала экспрессию p21 до детектируемых уровней. Однако коэкспрессия p53 и большого количества hNAP-1 привела к появлению белка p21. Кроме того, эндогенный p53 и hNAP-1 могут образовывать стабильный комплекс, как показано на фиг. 6D. Экстракты из клеток RTS3b, которые не экспрессируют детектируемый уровень белка p53, и из первичных неонатальных эпидермальных кератиноцитов человека, которые экспрессируют высокие уровни p53 (рис.6D, нижняя панель), подвергали иммунопреципитации моноклональным антителом против р53. Только в неонатальных эпидермальных кератиноцитах человека может быть соосажден эндогенный hNAP-1. Результаты, показанные на фиг. 6, предполагают, что p53 и hNAP-1 взаимодействуют напрямую друг с другом.
РИС. 6.p53 взаимодействует напрямую с hNAP-1. (A) Слитый белок GST-hNAP-1 или GST инкубировали с меченным His, бактериально экспрессированным, очищенным p53. После промывания шариков буфером, содержащим 200 мМ KCl, связанный р53 выявляли антителом, направленным против His-метки.(B) Р53-отрицательную клеточную линию RTS3b котрансфицировали синтетической р53-чувствительной репортерной конструкцией люциферазы, 5 нг вектора экспрессии для р53 и 400 нг вектора для HA-меченного hNAP-1. Активность люциферазы в присутствии только вектора была установлена на 1. Показаны полосы ошибок. (C) Экстракты (90 мкг) из клеток RTS3b, временно трансфицированных вектором экспрессии для p53 (дорожки 5, 7 и 8) или для HA-меченного hNAP-1 в двух разных количествах (дорожки 6-8), использовали для Вестерн блоттинг (WB) для обнаружения эндогенного p21.Блот повторно зондировали антителом против р53 для определения уровней белка р53. (D) Ядерные экстракты из p53-отрицательных клеток RTS3b (дорожки 2 и 4) и из неонатальных эпидермальных кератиноцитов человека (NHEK) (дорожки 1 и 3), которые экспрессируют p53 дикого типа, подвергали иммунопреципитации (IP) с p53 антитело. Связанный эндогенный hNAP-1 анализировали вестерн-блоттингом с моноклональным антителом против hNAP-1 (дорожки 3 и 4). На дорожках 1 и 2 была загружена шестая часть введенных ядерных экстрактов для определения уровня экспрессии hNAP-1 в обоих типах клеток.Дорожки 3 и 4 были повторно зондированы антителом к р53, чтобы подтвердить характер экспрессии р53. Перекрестно реагирующий клеточный белок обозначен звездочкой.
Чтобы проверить, является ли функциональное взаимодействие с hNAP-1 общей стратегией для активаторов или ограничивается активаторами, взаимодействующими с p300, мы использовали AD TEF-1. Различные исследователи показали, что TEF-1 не связывается с p300 in vitro и не взаимодействует с коэкспрессируемым p300 (49, 59). Как показано на фиг. 7, AD TEF-1 не может взаимодействовать с GST-hNAP-1 in vitro.TEF-1 AD слился с DBD стимулированной GAL4 транскрипции из репортера, содержащего сайты связывания GAL4 примерно в четыре раза; этот эффект не усиливался после коэкспрессии hNAP-1. Однако в качестве контроля активация гибридным белком, состоящим из AD E2 HPV8 и DBD GAL4, дополнительно стимулировалась коэкспрессируемым hNAP-1.
РИС. 7.ТЭФ-1 не взаимодействует с hNAP-1. (Левая панель) Вектор экспрессии для DBD GAL4, слитого либо с AD TEF-1 (pG4-TEF AD) (68), либо с AD HPV8 E2 (pG4-8E2 AD), котрансфицировали отдельно или вместе с вектор экспрессии для HA-меченного hNAP-1 и GAL4-зависимой репортерной конструкции, который схематически показан под графиком.Данные представляют собой средние значения четырех экспериментов, а планки ошибок указывают стандартные отклонения. (Правая панель) 35 S-меченный TEF-1 AD инкубировали с GST или GST-hNAP-1. После промывки образцов буфером, содержащим 100 мМ KCl, реакцию анализировали авторадиографией. Десять процентов ввода показаны на дорожке 1.
ОБСУЖДЕНИЕ
Появляется все больше свидетельств того, что hNAP-1 играет важную роль во время активации транскрипции. Было показано, что NAP-1 и другие шапероны гистонов, такие как ASF1 и фактор сборки хроматина 1, взаимодействуют с АТФ-зависимыми комплексами ремоделирования хроматина (4, 28, 48, 50).Кроме того, NAP-1 и NAP-2 входят в состав коактиваторного комплекса p300 / CBP (4, 27, 58). Таким образом, было высказано предположение, что NAP-1 может служить точкой взаимодействия между коактиваторами транскрипции и хроматином. Мы идентифицировали hNAP-1 как мишень для AD фактора транскрипции Е2 PV с помощью дрожжевого двугибридного скрининга. В отличие от предыдущего скрининга, в котором применялся точечный мутант E2, дефектный по трансактивации (9), мы смогли использовать белок E2 дикого типа в качестве приманки, поскольку наша репортерная конструкция только слабо активировалась E2 по сравнению с GAL4. ОБЪЯВЛЕНИЕ.Мы смогли подтвердить взаимодействие между E2 и hNAP-1 с помощью анализов GST pull-down и экспериментов по коиммунопреципитации. Таким образом, используя три различных метода и белки, синтезированные по-разному, то есть в дрожжевых, бактериальных и эукариотических клетках, мы смогли продемонстрировать взаимодействие между E2 и hNAP-1. Более того, E2 также осаждает эндогенный hNAP-1. Следовательно, маловероятно, что взаимодействие является артефактом в результате неправильного свертывания белков, но действительно специфично.
Помимо белка E2 BPV1, белки E2 двух типов HPV, HPV18 (инфицирование слизистой оболочки половых органов и связанное с раком шейки матки) и HPV8 (заражение кожи), также контактируют с hNAP-1 in vitro и in vivo, это указывает на то, что связывание с hNAP-1 может быть общим для белков E2.Взаимодействие между Е2 BPV1 и hNAP-1 является прямым, поскольку очищенные белки, экспрессируемые бактериями, также соосаждены. Кроме того, мы показываем, что другой активатор транскрипции, p53, также напрямую связывается с hNAP-1. Это открытие показывает, что взаимодействие не обязательно должно опосредоваться связующим фактором. p300 может быть таким фактором, поскольку hNAP-1 взаимодействует с p300, как и PV E2 и p53 (4, 5, 23, 49, 56, 58).
Связывание BPV1 E2 с hNAP-1 опосредуется его БА, как было выявлено с помощью дрожжевой двухгибридной системы, экспериментов с GST pull-down и коиммунопреципитации.Концепция взаимодействия между E2 и hNAP-1 подкрепляется способностью сверхэкспрессии hNAP-1 стимулировать активацию E2. Даже в присутствии насыщающих концентраций E2 hNAP-1 и E2 взаимодействовали, что указывает на то, что связывание с hNAP-1 является этапом, ограничивающим скорость для E2. Наблюдение за тем, что мутант E2 I73A со скомпрометированной активацией (14) стимулировался hNAP-1 в той же степени, что и E2 дикого типа, предполагает, что связывание с hNAP-1 не затронуто у этого мутанта, в корреляции с данными связывания in vitro. изображенный на рис.2Б. Таким образом, связывание с hNAP-1 может быть одним из нескольких этапов, выполняемых E2, ведущих к активации транскрипции. К ним относятся контакты с компонентами PIC, такими как TBP и TFIIB (8, 44). Кроме того, E2 связывается с коактиваторами, такими как AMF-1 / Gps, CBP / p300 и p / CAF (36, 37, 49, 56). Интересно, что Lefebvre et al. ранее показали, что активация транскрипции с помощью E2 коррелирует с изменением структуры хроматина в дрожжевых клетках (38), где взаимодействие с NAP-1 может играть роль.
Хотя E2 сильно активировал транскрипцию из репортерных конструкций, которые были временно трансфицированы, он не мог активировать транскрипцию, когда область энхансера-промотора была интегрирована в клеточный геном. Считается, что временно трансфицированные плазмиды собираются с гистонами в структуру, подобную нуклеосоме (10, 36). Однако возможно, что общая структура не организована как клеточный хроматин (10). Наблюдение за тем, что лечение TSA частично восстанавливает способность E2 активировать и действовать в синергии с hNAP-1, укрепляет представления о том, что компактная структура хроматина может ингибировать доступ E2 к хромосомной ДНК и что hNAP-1 может усиливать E2-опосредованную активация только после привязки E2.Таким образом, hNAP-1 не может стимулировать связывание E2 с сайтами, организованными в хроматине, как было показано для GAL4 in vitro (65).
Как hNAP-1 усиливает активацию E2? С использованием делеционных мутантов мы продемонстрировали, что весь hNAP-1 необходим для стимуляции активности E2. Взаимодействие E2 с hNAP-1 может быть необходимым, поскольку мутанты hNAP-1Δ3, hNAP-1Δ4 и hNAP-1Δ9, которые лишены одного или обоих взаимодействующих с E2 мотивов, не смогли или продемонстрировали сниженную способность стимулировать активацию E2.Помимо С-концевой 301 аминокислоты, которая связана как с E2, так и с p300, решающее значение имеют N-концевая 91 аминокислота hNAP-1. Все протестированные здесь мутанты с укороченным N-концом не только не обладали стимулирующей активностью, но также имели доминантно-отрицательный эффект на активацию E2. Некоторые из этих мутантов сверхэкспрессированного hNAP-1 с укороченным N-концом могут конкурировать с эндогенным hNAP-1 дикого типа за связывание с E2. Таким образом, привлечение N-терминально удаленного hNAP-1 с помощью E2 д. Приводить к нефункциональным комплексам, неспособным поддерживать транскрипцию.Более того, домены hNAP-1, экспрессируемые этими мутантами, могут связываться и секвестрировать другие клеточные факторы, необходимые для активации транскрипции. Это может иметь место в случае мутантов hNAP-1, которые все еще репрессируют активацию E2, но не связываются с E2 in vitro (такими как hNAP-1Δ7 или hNAP-1Δ10). Представление о том, что мишени, отличные от E2 или p300, которые участвуют в транскрипции, связаны с hNAP-1, подтверждается наблюдением, что некоторые из этих мутантов не только подавляли активацию с помощью E2, но также ингибировали базальную активность промоторов.
Какова роль N-концевой части hNAP-1? Согласно литературным данным, он не участвует в связывании с p300 (58). Было высказано предположение, что импорт гистонов h3A и h3B в ядро необходим для функции NAP-1 при активации транскрипции, поскольку мутации дрожжевого NAP-1, инактивирующие сигнал экспорта из ядра с высоким содержанием лейцина (NES) на N-конце, приводили к к снижению транскрипции некоторых генов (47). В нашем исследовании мутации соответствующих консервативных остатков в hNAP-1 не приводили к такому ингибированию.Кроме того, мутации также не повлияли на локализацию hNAP-1, что указывает на то, что человеческий аналог может иметь NES в другом положении (М. Рехтанц и Г. Стегер, неопубликованные результаты). Однако исследования локализации с нашими мутантами hNAP-1 показали более сильное ядерное окрашивание всех мутантов с N-концевой делецией, чем полноразмерных hNAP-1 или мутантов с C-концевым усечением. Точный мутационный анализ N-концевой 91 аминокислоты hNAP-1 должен показать, является ли ядерный импорт h3A и h3B, который требует функционального мотива NES, существенным для hNAP-1 для активации транскрипции.Более того, другие функции, которые были приписаны этому домену, такие как димеризация или мультимеризация NAP (58) или активность сборки нуклеосом (16), могут быть важны.
Привлечение hNAP-1 к промотору посредством слияния с DBD GAL4 не привело к активации транскрипции (M. Rehtanz и G. Steger, неопубликованные результаты), указывая на то, что hNAP-1 должен действовать согласованно с ОБЪЯВЛЕНИЕ. Наблюдение, что усиливающий эффект hNAP-1 на E2-опосредованную активацию был намного сильнее в присутствии субстратных количеств E2, можно объяснить тем фактом, что hNAP-1 влияет на взаимодействие E2 с другим фактором.p300 может быть таким фактором. Было показано, что p300 связывается с двумя разделяемыми доменами в пределах 300 C-концевых аминокислот hNAP-1 (58), как и E2. Наши данные демонстрируют, что in vitro возможно образование тройного комплекса между hNAP-1, p300 и E2. Во-первых, конкурентный эксперимент показал, что hNAP-1 не может вытеснить E2 из p300 (рис. 5B). Во-вторых, результаты седиментации градиента плотности глицерина коррелируют с образованием тройного комплекса. Очищенный hNAP-1 и p300 с 1195 по 1761 влияют на седиментацию E2Δ386-420, результат, который согласуется с прямыми взаимодействиями обоих белков с E2.В присутствии как hNAP-1, так и p300 седиментация E2 снова изменяется, указывая на то, что E2 образует комплекс не только с p300 или hNAP-1, но также с обоими белками одновременно. Более того, E2, hNAP-1 и p300 cofractionate при совместном инкубировании (рис. 5C). Этот тройной комплекс может быть очень эффективным при активации транскрипции, поскольку сверхэкспрессированный E2, hNAP-1 и p300 тесно взаимодействуют при активации экспрессии гена HPV8. Ито и др. продемонстрировали, что ацетилирование гистонов с помощью p300 помогает переносить гистоны h3A и h3B из нуклеосом в NAP-1 (27).In vitro отсутствие h3A и h3B коррелирует с повышенной активностью генов, вероятно, за счет снижения уровня сворачивания хроматина (см. Ссылку 11). Таким образом, E2 может индуцировать рекрутирование hNAP-1 и p300, которые затем участвуют в создании такой среды, свободной от h3A и h3B.
Важность hNAP-1, по-видимому, не ограничивается одним фактором, поскольку мы предоставляем дополнительные доказательства того, что бактериально экспрессируемый очищенный p53 может напрямую связываться с hNAP-1 in vitro и что эндогенный p53 и hNAP-1 также взаимодействуют.Это взаимодействие, по-видимому, является функциональным, вывод, который подтверждается наблюдением, что сверхэкспрессированный hNAP-1 сам по себе может стимулировать р53-опосредованную активацию из синтетической репортерной конструкции и из р53-чувствительного эндогенного промотора р21, что не обязательно требует коэкспрессия p300, как было предложено в другом месте (58). Поскольку мы смогли продемонстрировать, что TEF-1, лишенный связывания с p300, также лишен функционального взаимодействия с hNAP-1, hNAP-1 может функционировать специфически совместно с p300.Однако в будущих экспериментах необходимо будет подтвердить, является ли связывание с hNAP-1 феноменом только для активаторов, связывающихся с p300. Наши данные впервые демонстрируют, что помимо p300 важной мишенью для активации транскрипции является hNAP-1. Результатом могут быть достаточно высокие локальные концентрации hNAP-1 и p300 на активно транскрибируемых промоторах, которые необходимы для эффективной активации.
БЛАГОДАРНОСТИ
Мы благодарим Y. Ishimi за подарок моноклонального антитела против hNAP-1, D.Ливингстон и М. Шеффнер за предоставление плазмидных ДНК и П. Бартель за предоставление штамма дрожжей YM954. Мы также благодарим Герберта Пфистера и Эндрю Баркера за критическое прочтение рукописи и Карин Шнетц за полезную поддержку.
Эта работа была поддержана Deutsche Forschungsgemeinschaft (SFB274-A8, STE604 / 3-1 и STE604 / 3-2) и BMBF (Aktenzeichen 0312708F). Г.С. была получателем стипендии Lise-Meitner Habilitationsship.
СНОСКИ
- Поступило 5 ноября 2003 г.
- Принято 9 декабря 2003 г.
- Авторское право © 2004 Американское общество микробиологии
ССЫЛКИ
- 1.↵
Альтман, Р. и Д. Р. Келлог. 1997. Контроль митотических событий с помощью NAP1 и киназы Gin4. J. Cell Biol. 138 : 119-130.
- 2.
Армстронг, Дж. А., Дж. Дж. Бикер и Б. М. Эмерсон. 1998. Связанный с SWI / SNF комплекс ремоделирования хроматина, E-RC1, необходим для тканеспецифической регуляции транскрипции с помощью EKLF in vitro.Клетка 95 : 93-104.
- 3.↵
Асахара, Х., Б. Сантосо, Э. Гусман, К. Ду, П. А. Коул, И. Дэвидсон и М. Монмини. 2001. Хроматин-зависимая кооперативность между конститутивными и индуцибельными доменами активации в CREB. Мол. Клетка. Биол. 21 год : 7892-7900.
- 4.↵
Асахара, Х., С. Тартар-Декерт, Т. Накагава, Т. Икехара, Ф. Хиросе, Т. Хантер, Т. Ито и М. Монмини. 2002. Двойная роль p300 в сборке хроматина и активации транскрипции в сотрудничестве с белком сборки нуклеосом 1 in vitro.Мол. Клетка. Биол. 22 : 2974-2982.
- 5.↵
Авантаджати, М. Л., В. В. Огрызко, К. Гарднер, А. Джордано, С. А. Левин и К. Келли. 1997. Рекрутирование p300 / CBP в зависимых от p53 сигнальных путях. Клетка 89 : 1175-1184.
- 6.↵
Becker, P. B. 2002. Скольжение нуклеосом: факты и вымысел. EMBO J. 21 год : 4749-4753.
- 7.↵
Бенсон, Дж.Д. и П. М. Хоули. 1995. Аминоконцевые домены белков E1 и E2 вируса папилломы крупного рогатого скота 1 типа участвуют в комплексообразовании. J. Virol. 69 : 4364-4372.
- 8.↵
Бенсон, Дж. Д., Р. Лаванде и П. М. Хоули. 1997. Консервативное взаимодействие белков-активаторов транскрипции папилломавируса E2 с человеческими и дрожжевыми белками TFIIB. J. Virol. 71 : 8041-8047.
- 9.↵
Брейдинг, Д.E., F. Sverdrup, M. J. Grossel, N. Moscufo, W. Boonchai и E. J. Androphy. 1997. Функциональное взаимодействие нового клеточного белка с доменом трансактивации папилломавируса E2. Мол. Клетка. Биол. 17 : 7208-7219.
- 10.↵
Cervoni, N., and M. Szyf. 2001. Активность деметилазы направляется ацетилированием гистонов. J. Biol. Chem. 276 : 40778-40787.
- 11.↵
Чан, Х. М. и Н.Б. Ла Танге. 2001. p300 / CBP белки: HAT для транскрипционных мостов и каркасов. J. Cell Sci. 114 : 2363-2373.
- 12.↵
Донг, X.-P., Ф. Штубенраух, Э. Бейер-Финклер и Х. Пфистер. 1994. Распространенность делеций YY1-связывающих сайтов в эписомальной ДНК ВПЧ 16 от рака шейки матки. Int. J. Рак 58 : 803-808.
- 13.↵
Экнер, Р., М. Э. Юэн, Д. Ньюсом, М. Гердес, Дж.А. ДеКаприо, Дж. Б. Лоуренс и Д. М. Ливингстон. 1994. Молекулярное клонирование и функциональный анализ аденовирусного E1A-ассоциированного белка 300 кДа (p300) выявил белок со свойствами транскрипционного адаптера. Genes Dev. 8 : 869-884.
- 14.↵
Ferguson, M. K., and M. R. Botchan. 1996. Генетический анализ домена активации белка E2 вируса папилломы крупного рогатого скота: его роль в транскрипции и репликации. J. Virol. 70 : 4193-4199.
- 15.↵
Фрайер, К. Дж. И Т. К. Арчер. 1998. Ремоделирование хроматина глюкокортикоидным рецептором требует комплекса BRG1. Природа 393 : 89-91.
- 16.↵
Fujii-Nakata, T., Y. Ishimi, A. Okuda, and A. Kikuchi. 1992. Функциональный анализ белка сборки нуклеосом, NAP-1. J. Biol. Chem. 267 : 20980-20986.
- 17.↵
Funk, W. D., Д. Т. Пак, Р. Х. Карас, В. Э. Райт и Дж. У. Шэй. 1992. Транскрипционно активный сайт связывания ДНК для белковых комплексов р53 человека. Мол. Клетка. Биол. 12 : 2866-2871.
- 18.↵
Гиллитцер, Э., Г. Чен и А. Стенлунд. 2000. Отдельные домены в белках E1 и E2 выполняют архитектурные и продуктивные роли для кооперативного связывания ДНК. EMBO J. 19 : 3069-3079.
- 19.↵
Джордано, А., и М. Л. Авантаджати. 1999. p300 и CBP: партнеры на жизнь и смерть. J. Cell. Physiol. 181 : 218-230.
- 20.↵
Гири И., Янив М. 1988. Структурный и мутационный анализ трансактивирующих белков E2 папилломавирусов выявил три различных функциональных домена. EMBO J. 7 : 2823-2829.
- 21.↵
Гудман, Р. Х. и С. Смолик. 2000. CBP / p300 в росте, трансформации и развитии клеток.Genes Dev. 14 : 1553-1577.
- 22.↵
Grunstein, M. 1997. Ацетилирование гистонов в структуре и транскрипции хроматина. Природа 389 : 349-352.
- 23.↵
Gu, W., X.-L. Ши и Р. Г. Рёдер. 1997. Синергетическая активация транскрипции CBP и p53. Природа 387 : 819-823.
- 24.↵
Хэм, Дж., Г. Стегер и М. Янив. 1994 г.Кооперативность in vivo между трансактиватором E2 и связывающим белком ТАТА-бокса зависит от структуры корового промотора. EMBO J. 13 : 147-157.
- 25.↵
Hoffmann, A., and R.G. Roeder. 1991. Очистка белков, меченных гистами, в неденатурирующих условиях предлагает удобный метод исследования взаимодействия белков. Nucleic Acids Res. 19 : 6337-6338.
- 26.↵
Ито, Т., М. Балджер, Р.Кобаяси и Дж. Т. Кадонага. 1996. Drosophila NAP-1 является коровым гистоновым шапероном, который функционирует в АТФ-облегченной сборке регулярно расположенных массивов нуклеосом. Мол. Клетка. Биол. 16 : 3112-3124.
- 27.↵
Ито Т., Т. Икехара, Т. Накагава, В. Л. Краус и М. Мурамацу. 2000. p300-опосредованное ацетилирование облегчает перенос димеров гистона h3A-h3B из нуклеосом в шаперон гистона. Genes Dev. 14 : 1899–1907.
- 28.↵
Цзян В., С. К. Нордин и Дж. Т. Кадонага. 2000. Транскрипционный анализ хроматина, собранного с очищенным ACF и dNAP-1, показывает, что ацетил-КоА необходим для сборки преинициативного комплекса. J. Biol. Chem. 275 : 39819-39822.
- 29.↵
Kellogg, D. R., A. Kikuchi, T. Fujii-Nakata, C. W. Turck и A. W. Murray. 1995. Члены семейства белков NAP / SET специфически взаимодействуют с циклинами B-типа.J. Cell Biol. 130 : 661-673.
- 30.↵
Kingston, R. E., and G. J. Narlikar. 1999. АТФ-зависимое ремоделирование и ацетилирование как регуляторы текучести хроматина. Genes Dev. 13 : 2339-2352.
- 31.↵
Kowenz-Leutz, E., and A. Leutz. 1999. Изоформа C / EBP β рекрутирует комплекс SWI / SNF для активации миелоидных генов. Мол. Клетка 4 : 735-743.
- 32.№
Kraus, W. L., and J. T. Kadonaga. 1998. p300 и рецептор эстрогена совместно активируют транскрипцию посредством дифференциального усиления инициации и повторной инициации. Genes Dev. 12 : 331-342.
- 33.↵
Kraus, W. L., E. T. Manning, and J. T. Kadonaga. 1999. Биохимический анализ различных функций активации в p300, которые усиливают инициацию транскрипции с помощью хроматиновых матриц. Мол. Клетка. Биол. 19 : 8123-8135.
- 34.↵
Ламберт, П. Ф., Н. Достатни, А. А. Макбрайд, М. Янив, П. М. Хоули и Б. Аркангиоли. 1989. Функциональный анализ трансактиватора E2 вируса папилломы в Saccharomyces cerevisiae. Genes Dev. 3 : 38-48.
- 35.↵
Ламберт, П. Ф., Б. А. Спальхольц и П. М. Хоули. 1987. Репрессор транскрипции, кодируемый BPV1, имеет общий карбоксиконцевой домен с трансактиватором E2.Клетка 50 : 69-78.
- 36.↵
Ли, Д., С. Г. Хван, Дж. Ким и Дж. Чоу. 2002. Функциональное взаимодействие между p / CAF и белком E2 вируса папилломы человека. J. Biol. Chem. 277 : 6483-6489.
- 37.↵
Ли Д., Б. Ли, Дж. Ким, Д. В. Ким и Дж. Чоу. 2000. белок, связывающий элемент ответа цАМФ, связывается с белком E2 вируса папилломы человека и активирует E2-зависимую транскрипцию.J. Biol. Chem. 275 : 7045-7051.
- 38.↵
Лефевр, О., Г. Стегер и М. Янив. 1997. Синергетическая активация транскрипции белком E2 папилломавируса происходит после связывания ДНК и коррелирует с изменением структуры хроматина. J. Mol. Биол. 266 : 465-478.
- 39.↵
LeRoy, G., A. Loyola, W. S. Lane и D. Reinberg. 2000. Очищение и характеристика человеческого фактора, который собирает и модифицирует хроматин.J. Biol. Chem. 275 : 14787-14790.
- 40.↵
Ли, К., А. Имхоф, Т. Н. Коллингвуд, Ф. Д. Урнов и А. П. Вольф. 1999. p300 стимулирует транскрипцию, инициированную лиганд-связанным рецептором тироидного гормона на стадии, следующей за разрушением хроматина. EMBO J. 18 : 5634-5652.
- 41.↵
Martinez-Balbas, M. A., A. J. Bannister, K. Martin, P. Haus-Seuffert, M. Meisterernst, and T. Kouzarides. 1998. Ацетилтрансферазная активность CBP стимулирует транскрипцию. EMBO J. 17 : 2886-2893.
- 42.↵
McBride, A. A., J. C. Byrne, P. M. Howley. 1989. Полипептиды E2, кодируемые вирусом папилломы крупного рогатого скота типа 1, образуют димеры через общий карбоксиконцевой домен: трансактивация опосредуется консервативным аминоконцевым доменом. Proc. Natl. Акад. Sci. Соединенные Штаты Америки 86 : 510-514.
- 43.↵
Мейерс, К., М. Г. Фраттини и Л. А. Лайминьш. 1994. Методы культивирования тканей для изучения вирусов папилломы человека в стратифицированном эпителии. Academic Press, Inc., Орландо, Флорида,
- 44.↵
Miller Rank, N., and P.F. Lambert. 1995. Регуляторы транскрипции E2 вируса папилломы крупного рогатого скота типа 1 напрямую связывают два клеточных фактора транскрипции, TFIID и TFIIB. J. Virol. 69 : 6323-6334.
- 45.↵
Минк, С., Б. Хениг, К.-Х. Клемпнауэр. 1997. Взаимодействие и функциональное сотрудничество p300 и C / EBPβ. Мол. Клетка. Биол. 17 : 6609-6617.
- 46.
Мор, И. Дж., Р. Кларк, С. Сан, Э. Дж. Андрофи, П. Макферсон и М. Р. Ботчан. 1990. Нацеливание белка репликации E1 на ориджин репликации вируса папилломы путем образования комплекса с трансактиватором E2. Наука 250 : 1694-1699.
- 47.↵
Мосаммапараст, Н., К. С. Эварт и Л. Ф. Пембертон. 2002. Роль белка сборки нуклеосом 1 в ядерном транспорте гистонов h3A и h3B. EMBO J. 21 год : 6527-6538.
- 48.↵
Мошкин, Ю. М., Дж. А. Армстронг, Р. К. Маеда, Дж. В. Тамкун, К. П. Верриджзер, Дж. А. Кеннисон и Ф. Карч. 2002. Гистоновый шаперон ASF1 взаимодействует с механизмом ремоделирования хроматина Брахмы. Genes Dev. 16 : 2621-2626.
- 49.↵
Мюллер А., А. Рицковски и Г. Штегер. 2002. Совместная активация экспрессии гена вируса папилломы человека типа 8 белком E2 и клеточным коактиватором p300. J. Virol. 76 : 11042-11053.
- 50.↵
Накагава Т., М. Балджер, М. Мурамацу и Т. Ито. 2001. Многоступенчатая сборка хроматина на суперспиральной плазмидной ДНК с помощью белка-1 сборки нуклеосом и АТФ-использующего хроматина и фактора ремоделирования.J. Biol. Chem. 276 : 27384-27391.
- 51.↵
Накаджиама Т., К. Учида, С. Ф. Андерсон, Дж. Д. Парвин и М. Монмини. 1997. Анализ c-AMP-чувствительного активатора выявляет двухкомпонентный механизм индукции транскрипции через зависимые от сигнала факторы. Genes Dev. 11 : 738-743.
- 52.↵
Нили, К. Э., А. Х. Хассан, К. Э. Браун, Х. Линн и Дж. Л. Уоркман. 2002. Взаимодействие активатора транскрипции с множеством субъединиц SWI / SNF.Мол. Клетка. Биол. 22 : 1615-1625.
- 53.↵
Не, З., Я. Сюэ, Д. Ян, С. Чжоу, Б. Дж. Деру, Т. К. Арчер и В. Ван. 2000. Специфичность и нацеливающая субъединица человеческого комплекса ремоделирования хроматина, связанного с семейством SWI / SNF. Мол. Клетка. Биол. 20 : 8879-8888.
- 54.↵
Нигро, Дж. М., С. Дж. Бейкер, А. К. Прайзингер, Дж. М. Джессап, Р. Хостеттер, К. Клири, С. Х. Бигнер, Н. Дэвисдсон, С.Бейлин, П. Девили, Т. Гловер, Ф. С. Коллинз, А. Вестон, Р. Модали, К. К. Харрис и Б. Фогельштейн. 1989. Мутации в гене p53 встречаются в различных типах опухолей человека. Природа 342 : 705-708.
- 55.↵
Огрызко, В. В., Р. Л. Шильц, В. Руссанова, Б. Х. Ховард, Ю. Накатани. 1996. Коактиваторы транскрипции p300 и CBP представляют собой гистоновые ацетилтрансферазы. Клетка 87 : 953-959.
- 56.↵
Peng, Y.-C., Д. Э. Брейдинг, Ф. Свердруп, Дж. Ричард и Э. Дж. Андрофи. 2000. AMF-1 / Gps2 связывает p300 и усиливает его взаимодействие с белками папилломавируса E2. J. Virol. 74 : 5872-5879.
- 57. №
Purdie, K. J., C. J. Sexton, C. M. Proby, M. T. Glover, A. T. Williams, J. N. Stables, and I. M. Leigh. 1993. Злокачественная трансформация кожных поражений у пациентов с почечным аллотрансплантатом: роль вируса папилломы человека. Cancer Res. 53 : 5328-5333.
- 58.↵
Шикама, Н., Х. М. Чан, М. Крстич-Демонакос, Л. М. Смит, К.-В. Ли, У. Кэрнс и Н. Б. Ла Танге. 2000. Функциональное взаимодействие между белками сборки нуклеосом и коактиваторами семейства p300 / CREB-связывающих белков. Мол. Клетка. Биол. 20 : 8933-8943.
- 59.↵
Слепак, Т. И., К. А. Вебстер, Дж. Занг, Х. Прентис, А. О’Дауд, М. Н. Хикс и Н. Х. Бишоприк. 2001. Контроль кардиоспецифической транскрипции с помощью p300 посредством фактора-2D усилителя миоцитов.J. Biol. Chem. 276 : 7575-7585.
- 60.↵
Спальхольц, Б. А., П. Ф. Ламберт, К. Л. Ли и П. М. Хоули. 1987. Регуляция транскрипции вируса папилломы крупного рогатого скота: локализация E2-чувствительных элементов длинной контрольной области. J. Virol. 61 : 2128-2137.
- 61.↵
Tanese, N. 1997. Седиментация в мелкомасштабном градиенте плотности для разделения и анализа мультибелковых комплексов. Методы 12 : 224-234.
- 62.↵
Тайлер, Дж. 2002. Сотрудничество между гистоновыми шаперонами и АТФ-зависимыми машинами ремоделирования нуклеосом. Евро. J. Biochem. 269 : 2268-2274.
- 63.↵
Виньяли М., А. Хасан, К. Э. Нили и Дж. Л. Уоркман. 2000. АТФ-зависимые комплексы ремоделирования хроматина. Мол. Клетка. Биол. 20 : 1899-1910 гг.
- 64.↵
Валлберг, А. Э., К.Э. Нили, А. Хасан, Ж.-А. Густафссон, Дж. Л. Воркман и А. П. Райт. 2000. Рекрутирование комплекса ремоделирования хроматина SWI-SNF как механизм активации генов доменом активации рецептора глюкокортикоидов τ1. Мол. Клетка. Биол. 20 : 2004-2013 гг.
- 65.↵
Уолтер П. П., Т. А. Оуэн-Хьюз, Дж. Кот и Дж. Л. Воркман. 1995. Стимуляция связывания факторов транскрипции и замещения гистонов белком сборки нуклеосомы 1 и нуклеоплазмином требует разрушения октамера гистонов.Мол. Клетка. Биол. 15 : 6178-6187.
- 66.
Вебстер Н., Дж. Р. Джин, С. Грин, М. Холлис и П. Шамбон. 1988. UASG дрожжей представляет собой усилитель транскрипции в клетках HeLa человека в присутствии трансактиватора GAL 4. Клетка 52 : 169-178.
- 67. №
Уилсон, Р. Б., А. А. Бреннер, Т. Б. Уайт, М. Дж. Энглер, Дж. П. Гогран и К. Татчелл. 1991. Ген Saccharomyces cerevisiae SRK1 , супрессор bcy1 и ins1 , может быть вовлечен в функцию протеинфосфатазы.Мол. Клетка. Биол. 11 : 3369-3373.
- 68.↵
Сяо, Дж. Х., И. Дэвидсон, Х. Маттес, Ж.-М. Гарнье, П. Шамбон. 1991. Клонирование, экспрессия и транскрипционные свойства человеческого энхансерного фактора TEF-1. Клетка 65 : 551-568.
- 69.↵
Янг, Ю.-К., Х. Окаяма и П. М. Хоули. 1985. Вирус папилломы крупного рогатого скота содержит несколько трансформирующих генов. Proc. Natl. Акад. Sci. Соединенные Штаты Америки 82 : 1030-1034.
- 70.↵
Yao, J.-M., D. E. Breiding, and E. J. Androphy. 1998. Функциональное взаимодействие домена трансактивации Е2 вируса папилломы крупного рогатого скота с TFIIB. J. Virol. 72 : 1013-1019.
определение прямого взаимодействия | Словарь английских определений
прямой
vb в основном tr
1 для регулирования, ведения или контроля дел
2 также введите , чтобы отдавать приказы или приказы с властью (человеку или группе)
Он приказал им уйти
3 рассказать или показать (кому-то) путь к месту
4 прицеливаться, указывать или заставлять двигаться к цели
5 на адрес (письмо, посылка и др.)
6 в адрес (замечания, слова и т. Д.)
направлять комментарии кому-либо
7 также введите для руководства (актеров, операторов и т. Д.) При репетиции спектакля или съемке фильма
a Дирижировать (музыкальное произведение или музыканты), обычно во время выступления
b другое слово (особенно.США) для →
проводить →
9
прил.
9 без промедления и уклонения; прямой
прямой доступ
10 без сворачивания; непрерывный; самый короткий; прямой
прямой
11 без вмешательства лиц или агентств; сразу
прямая ссылка
12 честный; откровенный; откровенный
прямой ответ
13 обычно предварительный точный; точное
прямое предложение
14 диаметральная
прямая напротив
15 по непрерывной линии происхождения, как от отца к сыну в последующих поколениях
прямой потомок
16 (правительства, постановлений и т. Д.) избирателями или от них, а не через представителей
17 (Логика, математика) (доказательства), продвигаясь от посылок к заключению, вместо того чтобы исключить возможность ложности вывода
Сравнить →
косвенное доказательство
18 (Астрономия) движется с запада на восток по небесной сфере
Сравнить →
ретроградный →
4a
а или относящиеся к постоянному току
b (вторичного индуцированного тока), имеющего то же направление, что и первичный ток
a (движения) в том же направлении
См. →
движение →
9
b (интервала или хорды) в корневом положении; не инвертировано
adv
он пошел прямо в офис
(C14: от латинского directus; от dirigere, чтобы направлять, от разъединения + regere, чтобы управлять)
♦ непосредственность n
прямой доступ
n метод чтения данных из компьютерного файла без чтения файла с самого начала, как на диске или барабане, (также называется) произвольный доступ Сравнить →
последовательный доступ
прямого действия
n действий, таких как забастовки или гражданское неповиновение, нанятых организованными рабочими или другими группами для получения требований от работодателя, правительства и т. Д.
прямая связь
n (Электроника) проводящая связь между электронными цепями, в отличие от индуктивной или емкостной связи
См. Также →
муфта →
4
♦ прямая муфта прил.
постоянный ток
n непрерывный электрический ток, который течет только в одном направлении, без существенного изменения величины (аббревиатура.) DC Сравнить →
переменный ток
прямой дебет
n приказ, отданный банку или строительной кооперации владельцем счета с указанием выплатить указанному лицу или организации любую сумму, требуемую этим лицом или организацией
Сравнить →
постоянный платеж
прямой телефонный номер
n U.S. и канадский эквивалент →
абонентская линия набора
прямой краситель
n любой из ряда красителей, которые можно наносить без использования протравы. Обычно это азокрасители, наносимые на хлопок или вискозу из жидкой ванны, содержащей электролит, такой как сульфат натрия
прямые доказательства
n (Правовые) доказательства, обычно показания свидетеля, непосредственно относящиеся к обсуждаемому факту
Сравнить →
косвенные доказательства
школа с прямыми грантами
n (ранее в Великобритании) школа, финансируемая за счет пожертвований, платы за обучение и государственной субсидии при условии зачисления определенного процента не платящих учеников, назначенных местным органом образования
прямой впрыск
n см. →
твердый впрыск
прямой ввод
n устройство, такое как клавиатура, используемое для ввода данных непосредственно в компьютеризированную систему
прямой труд
n (торговля)
1 работа, которая является неотъемлемой частью производственного процесса или оказания услуги
Сравнить →
непрямой труд
2 (брит.) Рабочие, которые являются частью собственной рабочей силы работодателя, а не наняты через подрядчика, например, строительные рабочие, нанятые местными властями
прямое освещение
n (Электротехника) система освещения, в которой большая часть (не менее 90 процентов) света направлена вниз
прямая почтовая рассылка
n (маркетинг) рассылка незапрошенной коммерческой литературы на дома или рабочие адреса потенциальных клиентов
прямой маркетинг
n продажа товаров напрямую потребителям, а не через розничных торговцев, обычно по почте, прямой почтовой рассылке, газетной рекламе, продажам от двери до двери или по телефону (также называется) прямые продажи
прямой доступ к памяти
n процесс, в котором данные могут быть перемещены непосредственно в или из основной памяти компьютерной системы с помощью операций, не находящихся под контролем центрального процессора (аббревиатура.) DMA
прямой метод
n метод обучения иностранному языку с минимальным использованием родного языка ученика и формальной грамматики
прямой объект
n (Грамматика) существительное, местоимение или именная фраза, референт которой получает прямое действие глагола. Например, книга является прямым объектом в предложении Они купили Анне книгу
Сравнить →
косвенный объект
прямой первичный
n (U.S. правительство) первичные выборы, на которых избиратели напрямую выбирают кандидатов, которые будут баллотироваться на должность
прямой вопрос
n вопрос, задаваемый в прямой речи, например, Почему вы пришли?
Сравнить →
косвенный вопрос
прямое считывание
adj (прибора) откалибровано таким образом, чтобы заданная величина, подлежащая измерению, могла быть считана непосредственно со шкалы без необходимости использования постоянной умножения
прямые продажи
n другое название для →
прямой маркетинг
прямая речь , (особенно.США) прямой дискурс
n сообщение о том, что кто-то сказал или написал, цитируя его точные слова
прямой налог
n налог, уплачиваемый лицом или организацией, с которых он взимается
Сравнить →
косвенный налог
♦ прямое налогообложение n
представителей службы поддержки клиентов утверждают, что лица, напрямую взаимодействующие с ними, неправильно классифицировали свои рабочие места, чтобы не платить сверхурочные
Сообщается, что представители службы поддержки клиентов (CSR), работающие на дому в Direct Interactions, Inc.были вынуждены работать в качестве независимых подрядчиков и им отказывали в оплате сверхурочных в нарушение федерального закона о труде.
Согласно утверждениям в недавно поданном иске, Direct Interactions требует от своих CSR работать не менее 30 часов в неделю, но не может компенсировать им работу «вне рабочего времени»:
- Под угрозой штрафа компания не позволяет своим служащим вести отсчет времени более чем за три минуты до начала смены, независимо от того, сколько времени может потребоваться служащему для загрузки их компьютеров и запуска программ, необходимых для их работы.В среднем процесс запуска и входа в систему занимает от 5 до 10 минут в день, а иногда и до 15 минут.
- Компания требует, чтобы CSR проверяли электронную почту, связанную с работой, перед тем, как приступить к работе.
- Во время смены CSR часто лишены обязательных перерывов на обед и отдых, если объем звонков велик. Перерыв на обед обычно составляет 30 минут.
- CSR не оплачиваются, если по техническим причинам они отключаются от серверов компьютеров компании, что происходит 2-3 дня в неделю.
- Один CSR сообщил, что от него требовалось попросить разрешения на перерыв в туалете, в чем руководитель иногда отказывался. Если CSR не вернется к работе в течение 5 минут после перерыва в туалете, системы компании автоматически отключатся, и это время, а также время, потраченное на повторный вход в систему, не будут оплачены.
Каждое из этих злоупотреблений является нарушением федерального закона о справедливых трудовых стандартах, и из-за неоплачиваемого рабочего времени количество рабочих часов в неделю превышает 40-часовой порог сверхурочной работы.Тем не менее, явно пытаясь обойти FLSA, Direct Interactions вынуждает CSR подписывать «Соглашение с независимым подрядчиком», которое характеризует их как «поставщиков», освобожденных от сверхурочной компенсации.
Как мы уже писали в предыдущих публикациях, злоупотребления в сфере заработной платы в call-центрах слишком распространены и являются предметом расследования Отдела заработной платы и часов Министерства труда США, который выпустил информационный бюллетень, чтобы предупредить звонящих на дому операторов и сотрудники колл-центра о том, как работодатели могут попытаться удержать компенсацию.
Адвокаты Группы по трудовым спорам Соммерса Шварца инициировали коллективный иск от имени представителей службы поддержки Direct Interactions по всей стране, утверждая, что их права были нарушены и что они имеют право на взыскание невыплаченной заработной платы. Если вы работали в Direct Interactions, Inc. в качестве представителя службы поддержки клиентов и подозреваете, что стали жертвой кражи заработной платы, свяжитесь с нами сегодня!
[contact-form-7 id = ”2392 ″ title =” Контактная форма для коллективных действий ”]
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Есть много причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или уточнить у системного администратора.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует от сайта создания нового сеанса для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файлах cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
ATLAS наблюдает прямое взаимодействие бозона Хиггса с топ-кварком
Это позволяет физикам проверить критические параметры механизма Хиггса в Стандартной модели физики элементарных частиц.
В результате используется полный набор данных, доставленных в ATLAS Большим адронным коллайдером (LHC), и устанавливается сигнал со статистической значимостью 6,3 стандартного отклонения. Это согласуется с недавним наблюдением CMS Collaboration со значимостью 5,2 стандартных отклонения при использовании меньшего набора данных. «Это измерение представляет собой знаменательное достижение в исследовании механизма Хиггса и взаимодействия частиц Стандартной модели с бозоном Хиггса», — говорит Карл Якобс, пресс-секретарь ATLAS.«Это является прямым доказательством того, что самая тяжелая из известных частиц, топ-кварк, взаимодействует с предсказанной большой силой с бозоном Хиггса». Сигнал для этого производства медленно нарастал за годы накопления данных, и первые свидетельства были опубликованы ATLAS в декабре прошлого года.
«Это измерение представляет собой знаменательное достижение в исследовании механизма Хиггса и взаимодействия частиц Стандартной модели с бозоном Хиггса». — Карл Якобс, пресс-секретарь ATLAS.
Поскольку только 1% всех бозонов Хиггса рождается в ассоциации с топ-кварками, наблюдение было чрезвычайно сложным. Чтобы прийти к такому результату, физики ATLAS изучили данные о столкновениях за пять лет. «Это был один из самых сложных поисков, когда-либо выполнявшихся ATLAS Collaboration, и требовал согласованных усилий нескольких аналитических групп», — говорит Фабио Черутти, руководитель рабочей группы ATLAS Higgs. «Поскольку это такой редкий процесс, нам пришлось изучить множество различных каналов распада бозона Хиггса.Некоторые из них были ограничены экспериментальными неопределенностями, а другие — количеством собранных данных. Только объединив все эти различные анализы, мы смогли добиться этого наблюдения ».
Новый результат может также пролить свет на один из самых загадочных аспектов Стандартной модели: широкий диапазон масс фермионов, класс частиц, составляющих материю и включающих кварки и лептоны. Понимание природы массы топ-кварка во многом поможет решить этот открытый вопрос.«Измерение убедительно свидетельствует о том, что бозон Хиггса играет ключевую роль в большом значении массы верхнего кварка», — говорит Черутти. «Хотя это, безусловно, ключевая особенность Стандартной модели, это первый раз, когда она была подтверждена экспериментально с огромной значимостью».
Успех этого результата намекает на новые аналитические возможности эксперимента ATLAS. Благодаря большому количеству данных, полученных с помощью LHC, и отличным характеристикам эксперимента ATLAS, физики впервые смогут изучать бозон Хиггса в дальнейших редких и экспериментально сложных взаимодействиях.Такие исследования будут и дальше ставить под сомнение ограничения Стандартной модели и могут открыть новые возможности для открытий.