Тест «акцентуация личности» версия н. Козлова инструкция
Николай Козлов при грандиозном, просто оглушительном внешнем лоске всегда вызывал у меня недоверие. Но разве может ли быть так, что человек имеющий, наверное все возможные регалии и признание коллег — всего лишь огромный мыльный пузырь? Николай Козлов имеет от самых разных организаций. Деятельность Николая Козлова одобряют коллеги: Перечислять заслуги можно было бы и продолжить, но это не принесет никакой пользы. Итак понятно, что Николай Козлов сияет ярче сверхновой. А можно ли найти хоть маленькое пятнышко в этом ослепительном блеске? И где его искать? Как ни открою, как ни начну читать — так восхищаюсь и стилем, и содержанием.
Козлов Николай Иванович
Они ведут дело к расколу, начиная работать внутри Церкви, на ее окраинах. В наш Центр уже обращаются люди, близкие которых попали в общины лжестарцев, группировки, занятые борьбой с ИНН, озабоченные канонизацией царя Иоанна Грозного и т. Все эти маргинальные, но очень громкие группировки часто создают лицо православию.
В нашем городе есть и такие, как например»Опричное братство во имя благоверного царя Иоанна Грозного» возглавляемое своеобразным гуру Сабуровым.
Прот. Максим Козлов не советует отказываться от службы из страха перед гриппом, если только мы сами не понесем вирус в храм.
А для меня ЛП — это совсем другое! Света Стали мы с Антоном жить, все вроде бы нормально, но скоро я на него начала наезжать: Что ты на самом деле хочешь? Что-то мне не нравится Что мне не нравится? Почему ты посуду не помыл?? Для меня это — одно, для него это — другое. Для меня любовь, это когда обо мне заботятся, все делают, и я, раз Антона люблю, для него все делаю. А он — не делает! Значит, думаю я, не любит, и начинаю возмущаться.
Псевдоправославная секта»Опричное братство»
Это нужно для того чтобы, во-первых, быть в курсе всех своих слабых мест, а во-вторых, чтобы знать, на чем натренировать свой навык борьбы со страхом, на чем, так сказать, набить руку. Теперь временно оставим практические занятия и обратимся к механизмам страха. страхи, к сожалению, почти неотъемлемая часть нашей жизни. Они пронизывают ее снизу доверху и от края до края. Я привел примеры, и, наверное, некоторые из них похожи на забавные психотерапевтические байки.
страхи, к сожалению, почти неотъемлемая часть нашей жизни. Они пронизывают ее снизу доверху и от края до края. Я привел примеры, и, наверное, некоторые из них похожи на забавные психотерапевтические байки.
романтической поэзии, об разно названной Н. В. Гоголем «поэтической ло неизвестно в нашей литературе. Г. Козлов первый знакомит русскую публику с сим . но жить и помнить божий страх, // От нужд искать убежища в.
Вам скажут, что лицо у вас потеплело. А теперь подойдите к другу еще ближе и тепло его обнимите. Замрите и отследите еще раз выражение своего совсем теплого и, скорее всего, очень довольного лица. А теперь разнимите объятия, сделайте шаг назад и тресните этого друга по физиономии. А теперь отследите СВОЕ выражение лица. Простите, мы забыли сделать уточнение: Если вы это прожили, теперь вы знаете один из самых эффективных законов практической психологии: Меняя внутренние жесты, вы можете менять свое отношение к людям.
Если вы, разговаривая с человеком на любой дистанции, хоть по телефону! Если, подходя к человеку, вы представляете, что ваши руки немного приоткрываются в дружеские предобъятия, вы уже заранее проживаете ваши теплые объятия с этим человеком — выражение вашего лица становится открытым, теплым и внимательным. Это один из самых простых рецептов, как сделать так, чтобы люди вас не боялись, и состоит он из двух пунктов:
Это один из самых простых рецептов, как сделать так, чтобы люди вас не боялись, и состоит он из двух пунктов:
Часть 2. Мир душевной жизни
В ближайшее время страница будет существенно пополняться. Планируется не только выложить здесь наиболее ценные философские труды как уже сделано сейчас — пока мы просто в алфавитном порядке выстроили фамилии авторов , но и систему изучения философского наследия. То есть это будет определенная последовательность работ, перемежающаяся современными комментариями и пояснениями, а также методическая часть. На этой же страничке мы будем публиковать в электронном виде номера журнала русской интеллигенции»Философская культура», главным редактором которого является санкт-петербургский философ Николай Петрович Ильин.
Персональный сайт Николая Козлова, психолога и писателя. Мои книги по психологии прочитали уже 10 миллионов читателей. Пишу дальше! Как вам.
Если бы он всегда оставался таким и вне Синтона! А проблема в следующем… У многих синтоновцев на уровне условного рефлекса закрепилось: А ты кто такой?! Синтон — Синтоном, а жизнь — жизнью. Где-то это разделение исключительно понятно и глубоко оправданно: Но грош цена была бы Синтону, если бы он учил жизни только в Синтоне. А самое главное, народу самому хочется и интересно: С утра — до вечера?
Где-то это разделение исключительно понятно и глубоко оправданно: Но грош цена была бы Синтону, если бы он учил жизни только в Синтоне. А самое главное, народу самому хочется и интересно: С утра — до вечера?
В окружении проблем не игровых, а самых что ни на есть реальных? Чего мы действительно стоим? Так родилась идея сделать свой, летний синтоновский монастырь. Всегда были люди, которые были убеждены:
Козлов, Николай Иванович
Сегодня публикую письмо от одной из читательниц и свой на него ответ. Думаю, все будет ясно из контекста. Здравствуйте, дорогой Николай Иванович. Меня мучает один очень актуальный для меня вопрос.
По сути ум-эго – это один большой страх жить на земле и чувствовать, страхи – это стражи ума, не позволяющие ему выйти за свои пределы и.
Истинная правда, или учебник для психолога по жизни. Написано для книжного обозрения Независимой газеты Работы Николая Козлова нельзя не признать одним из феноменов современного книжного рынка. Его произведения издаются и переиздаются астрономическими для нашего времени тиражами. Появилось уже несколько изданий, сделанных» под Козлова» — с похожей обложкой и оформлением. Его имя известно продавцам и на лотках, и в элитарных книжных магазинах — и там, и там постоянны вопросы покупателей: И все это — на фоне огромного количества аналогичных по тематике американских или американизированных изданий типа» Как добиться успеха» ,» Как бороться со стрессом» или» Как читать человека словно книгу» — учебников жизни, где по пунктам расписаны ее правила и даны рецепты, прямой дорогой ведущие к успеху семейному, деловому, всеобщему, недостающее вписать, ненужное вычеркнуть.
Его произведения издаются и переиздаются астрономическими для нашего времени тиражами. Появилось уже несколько изданий, сделанных» под Козлова» — с похожей обложкой и оформлением. Его имя известно продавцам и на лотках, и в элитарных книжных магазинах — и там, и там постоянны вопросы покупателей: И все это — на фоне огромного количества аналогичных по тематике американских или американизированных изданий типа» Как добиться успеха» ,» Как бороться со стрессом» или» Как читать человека словно книгу» — учебников жизни, где по пунктам расписаны ее правила и даны рецепты, прямой дорогой ведущие к успеху семейному, деловому, всеобщему, недостающее вписать, ненужное вычеркнуть.
Первыми для нашего читателя в ряду подобных книг, еще в дорыночные времена, стали руководства Дейла Карнеги, а позднее такого рода издания хлынули неиссякающим потоком. На этом фоне вряд ли возможно объяснить» феномен Козлова» одной лишь умелой раскруткой или дурным вкусом массового читателя. Скорее возникнет мысль о том,что эти самые читатели находят в книгах Козлова нечто, отсутствующее в руководствах американского типа, но близкое и необходимое нашему родному менталитету.
Книга Николая Козлова, без ложной скромности названная» Истинная правда, или учебник для психолога по жизни» , будет, без сомнения, с восторгом встречена его постоянными читателями и почитателями. Тот, кто впервые столкнется с Козловым, взяв в руки именно эту книгу, получит прежде всего увлекательное чтение, от которого трудно оторваться.
Отношение же к содержанию прочитанного может оказаться разным — интересно, полезно, неожиданно, спорно, возмутительно. Вполне возможно — все это одновременно. В книге можно найти серию рецептов Карнеги на тему» Как надо знакомиться» под дождем, в магазине, в очереди, у киноафиши, а также знакомство методом взлома и советов для особо стеснительных больше общайтесь, следите за своим лицом.
Страх неизвестного
Практическая психология Это книга о вкусной и здоровой жизни. Как живой и пристрастный собеседник, она поможет обычному человеку открыть в себе психолога — психолога-практика, психолога по жизни, а тем, кто уже психолог, — не потерять в себе Человека. Поскольку истинная правда — всегда объемна, то книга вам поможет увидеть, кроме правды своей, правду другого человека, совсем разную правду мужчин и женщин и правду общечеловеческую — правду детей, которыми, похоже, является каждый из нас.
Поскольку истинная правда — всегда объемна, то книга вам поможет увидеть, кроме правды своей, правду другого человека, совсем разную правду мужчин и женщин и правду общечеловеческую — правду детей, которыми, похоже, является каждый из нас.
Книга всерьез отвечает на много веселых как.
Все стихи русского поэта Ивана Козлова на одной странице. И грусть свою , и тайный страх В молчаньи скрыв тяжелом, С слезами часто на глазах.
Это книги, которые просто помогают жить хорошо. В философском смысле это книги человека, который так ни в чем и не разобрался. Чувство превосходства над другими людьми в данном случае презрение к женщинам — вот он показатель глубоких невычищенных комплексов и заморочек. Особенно, где речь идет об отношениях внутри семьи, почему-то все, что относится к терпению, прощению и вообще, грубо говоря, к принятию всех косяков мужа-сожителя Васи касается именно женщины, а о мужчинах речь как-то идет слабовато Подразумевается, что это мы, женщины, должны начитаться Козлова и быть такими писями-одуванчиками, а мужики сядут нам, умным и всепонимающим, на шею и будут со спокойной душой по-сволочному к нам относится нет, конечно не все, есть и умные мужчины.
А я говорю все же о тех, кто не то что Козлова с его семейными правилами не читал, но и вообще книг не читает, а любит в гараже ковыряться и пиво пить с такими же грубыми животными, как он сам. А мне мой еще и попреки делает, когда я его пытаюсь учить психологическим тонкостям:
Козлов Н. Подборка текстов
Разместите, пожалуйста, ссылку на эту страницу на своём веб-сайте: Код для вставки на сайт или в блог: Код для вставки в форум : Если это не так — свяжитесь с администрацией сайта.
Санкт-Петербургский — театр «Мастерская» п/р Григория Козлова | Санкт- Петербург. Меню . — «Любовь сильнее страха» · — С Новым годом! — Гастроли в Москве на сцене ТЦ «На Страстном ».
Понедельник, 29 Апреля г. В таком подходе считается, что бороться с нежелательными эмоциями непродуктивно, часто транслируется тезис»Плохих эмоций не бывает» и предлагается любую эмоцию в первую очередь принять, как живое существо, заботящееся о клиенте. Вместо силового подхода к нежелательным эмоциям здесь предлагается продемонстрировать к ним подход дружественный и уважительный, в расчете на ответную взаимность.
Работа с эмоциями преполагает, что любая эмоция — это всегда нечто серьезное и значительное, от чего нельзя просто отмахнуться и проигнорировать; это то, чем заниматься нужно всерьез и скорее всего долго, поскольку убрать эмоцию нельзя, а неладить с нею отношение — непросто. Сотрудника можно уволить, а детей приходится воспитывать. Это, конечно, более трудная задача. Всегда ли к нежелательным эмоциям необходимо относиться именно так? К примеру, серьезное уважительное отношение к страхам ребенка оправдано не всегда.
Иногда испугавшемуся ребенка лучше всего помогает веселый взгляд веселый взгляд и бодрый голос взрослого:
Наука о душе — 26. А. Гордон
Тест акцентуации Н. Козлова — Записки мечтателя — ЖЖ
Я и не сомневалась!Шизоид
С детских лет поражает ребёнок, который любит играть один, не тянется к сверстникам, избегает шумных забав, предпочитает держаться среди взрослых, иногда подолгу молча слушает их беседы. К этому иногда добавляется какая-то холодность и недетская сдержанность. Малоподвижное, как будто замороженное лицо… Это — шизоид.
К этому иногда добавляется какая-то холодность и недетская сдержанность. Малоподвижное, как будто замороженное лицо… Это — шизоид.
Шизоиду природа выморозила чувства и вставила один интеллект, причём, как правило, без ориентиров. Смутный и плотный мир переживаний, тень предчувствий и пламя страстей, радость и стыд чувственности, всё то, что так близко поэтам и женщинам, — всё это не его мир. Он может им интересоваться, но усваивает его только в качестве иностранного языка. Его мир — мир понятий, схем и вычислений.
И что ему с этим делать среди людей, то есть того, что не вычисляется?
Он может разложить и проанализировать. А обольстить и соблазнить — для него так же невозможно, как пройти сквозь стену вслед за домашним привидением: ему нечем это сделать, у него для этого просто нет органов. У него отсутствует интуиция, и в компании ему трудно, как и компании с ним, где он всегда «белая ворона» и почти всегда «некстати». Он не может ловить «тонкие вещи» во взаимоотношениях: когда что сказать или промолчать, когда уйти или остаться.
Поэтому, как правило, он уходит.
В результате это человек, как будто всегда отгороженный от окружающих какой-то невидимой, как бы стеклянной, стеной. Он не может, да и редко хочет, разомкнуть своё одиночество и выйти к людям, к себе же во внутренний мир пускает крайне неохотно. Шизоид — это человек-отшельник, человек «не от мира сего». Чаще всего он неразговорчив («не знаю, о чём ещё говорить»), однако с пол-оборота «грузит» подходящего собеседника своими теориями, например о строении мира. Шизоиды — это великие теоретики. Не умея поддерживать самый элементарный светский трёп, они глубоко просистематизируют и обобщат процесс общения; в жизни их элементарно обведут вокруг пальца, зато они напишут работу по психологии и философии человеческих взаимоотношений. В жизни они беспомощны, но в теории — всесильны.
А научные работы их называются примерно так: «Самая общая теория Всего».
Если шизоида не трогать, он совершенно безобиден, но при этом не приручается и остаётся «вещью в себе». Близким людям рядом с ним спокойно, но неуютно: шизоид не даст им ни тепла, ни любви и благодарности, а некоторые его поступки могут выглядеть жестокими. На самом деле он не жесток, а просто аналитичен, а его холодность — скорее неспособность почувствовать душу другого, отсутствие эмоционального резонанса. Тем не менее он всё-таки человек, и, если его «скорлупа» даёт трещину, он обнаруживает неожиданную для окружающих ранимость.
Близким людям рядом с ним спокойно, но неуютно: шизоид не даст им ни тепла, ни любви и благодарности, а некоторые его поступки могут выглядеть жестокими. На самом деле он не жесток, а просто аналитичен, а его холодность — скорее неспособность почувствовать душу другого, отсутствие эмоционального резонанса. Тем не менее он всё-таки человек, и, если его «скорлупа» даёт трещину, он обнаруживает неожиданную для окружающих ранимость.
ТЕСТ АКТИВАЦИИ БАЗОФИЛОВ ДЛЯ ДИАГНОСТИКИ МИКОГЕННОЙ СЕНСИБИЛИЗАЦИИ У БОЛЬНЫХ МУКОВИСЦИДОЗОМ | Козлова
1. Козлова Я.И., Учеваткина А.Е., Бычкова Н.С., Филиппова Л.В., Аак О.В., Пятакова А.В., Фролова Е.В., Давыдова Н.И., Климко Н.Н. Тест активации базофилов в диагностике аллергического бронхолегочного аспергиллеза // Проблемы медицинской микологии, 2016. Т. 18, № 3. С. 7-11.
2. Красовский С. А., Самойленко В.А., Амелина Е.Л. Муковисцидоз: диагностика, клиника, основные принципы терапии // Пульмонология и аллергология, 2013. № 1. С. 42-46.
А., Самойленко В.А., Амелина Е.Л. Муковисцидоз: диагностика, клиника, основные принципы терапии // Пульмонология и аллергология, 2013. № 1. С. 42-46.
3. Agarwal R., Chakrabarti A., Shah A., Gupta D., Meis J.F., Guleria R., Moss R., Denning D.W. For the ABPA complicating asthma ISHAM working group 2013. Allergic bronchopulmonary aspergillosis: Review of literature and proposal of new diagnostic and classification criteria. Clin. Exp. Allergy, 2013, Vol. 43, no. 8, pp. 850-873.
4. Baxter C.G., Dunn G., Jones A.M., Webb K., Gore R., Richardson M.D., Denning D.W. Novel immunologic classification of aspergillosis in adult cystic fibrosis. J. Allergy Clin. Immunol., 2013, Vol. 132, no. 3, pp. 560-566.
5. Baxter C.G., Moore C.B., Jones A.M., Webb A.K., Denning D. W. IgE-mediated immune responses and airway detection of Aspergillus and Candida in adult cystic fibrosis. Chest, 2013, Vol. 143, no. 5, pp. 1351-1357.
W. IgE-mediated immune responses and airway detection of Aspergillus and Candida in adult cystic fibrosis. Chest, 2013, Vol. 143, no. 5, pp. 1351-1357.
6. Boumiza R., Debard A.L., Monneret G. The basophil activation test by flow cytometry: Recent development in clinical studies, standardization and emerging perspectives. Clin. Mol. Allergy, 2005, no. 3, pp. 9-13.
7. Chirumbolo S., Vella A., Ortolani R., de Gironcoli M., Solero P., Tridente G., Bellavite P. Differential response of human basophil activation markers: a multiparameter flow cytometry approach. Clin. Mol. Allergy, 2008, no. 6, pp. 12-16.
9. Chirumbolo S.H. Basophil activation test in allergy: time for an update? Int. Arch. Allergy Immunol., 2012, no. 158, pp. 99-114.
Chirumbolo S.H. Basophil activation test in allergy: time for an update? Int. Arch. Allergy Immunol., 2012, no. 158, pp. 99-114.
10. Chotirmall S.H., Branagan P., Gunaratnam C., McElvaney N.G. Aspergillus/allergic bronchopulmonary aspergillosis in an Irish cystic fibrosis population: A diagnostically challenging entity. Respir. Care, 2012, Vol. 53, no. 8, pp. 1035-1041.
11. Delhaes L., Monchy S., Fréalle E., Hubans C., Salleron J., Leroy S., Prevotat A., Wallet F., Wallaert B., Dei-Cas E., Sime-Ngando T., Chabé M., Viscogliosi E. The airway microbiota in cystic fibrosis: a complex fungal and bacterial community – implications for therapeutic management. PLoS ONE, 2012, Vol. 7, no. 4, e36313. doi: 10.1371/journal.pone.0036313.
12. Denning D.W., Pashley C.
13. Farrell Ph.M., White T.B., Ren Cl.L., Hempstead S.E., Accurso F., Derichs N., Howenstine M., McColley S.A., Rock M., Rosenfeld M., Sermet-Gaudelus I., Southern K.W., Marshall B.C., Sosnay P.R. Diagnosis of Cystic Fibrosis: Consensus Guidelines from the Cystic Fibrosis Foundation. J. Pediatr., 2017, Vol. 181, pp. 4-15.
15. Janahi I.A., Rehman A., Al-Naimi A.R. Allergic bronchopulmonary aspergillosis in patients with cystic fibrosis. Ann. Thorac. Med., 2017, Vol. 12, no. 2, pp. 74-82.
Janahi I.A., Rehman A., Al-Naimi A.R. Allergic bronchopulmonary aspergillosis in patients with cystic fibrosis. Ann. Thorac. Med., 2017, Vol. 12, no. 2, pp. 74-82.
16. Karasuyama H., Tsujimura Y., Оbata K., Mukai K. Role for basophils in systemic anaphylaxis. Anaphylaxis. Chem. Immunol. Allergy, 2010, Vol. 95, pp. 85-97.
17. Kang M.G., Song W.J., Park H.K., Lim K.H., Kim S.J., Lee S.Y., Kim S.H., Cho S.H., Min K.U., Chang Y.S. Basophil activation test with food additives in chronic urticaria patients. Clin. Nutr. Res., 2014, Vol. 3, no. 1, pp. 9-16.
18. Knutsen A.P., Kariuki B., Consolino J.D., Warrier M.R. IL-4 alpha chain receptor (IL-4R alpha) polymorphisms in allergic bronchopulmonary aspergillosis. Clin. Mol. Allergy, 2006, no. 4, p. 3.
19. Maturu V.N., Agarwal R. Prevalence of Aspergillus sensitization and allergic bronchopulmonary aspergillosis in cystic fibrosis: Systematic review and meta-analysis. Clin. Exp. Allergy, 2015, Vol. 45, no. 12, pp. 1765-1778.
Maturu V.N., Agarwal R. Prevalence of Aspergillus sensitization and allergic bronchopulmonary aspergillosis in cystic fibrosis: Systematic review and meta-analysis. Clin. Exp. Allergy, 2015, Vol. 45, no. 12, pp. 1765-1778.
20. Mircovic B., Lavelle G.M., Abdul Azim A., Helma K., Gargoum F.S., Molloy K., Gernez Y., Dunne K., Renwick J., Murphy P., Moss R.B., Greene C.M., Gunaratnam C., Chotirmall S.H., McElvaney N.G. The Basophil surface marker CD203с identifies Aspergillus species sensitization in patients with cystic fibrosis. J. Allergy Clin. Immunol., 2016, Vol. 137, no. 2, pp. 436-443.
21. McMahon M.A., Chotirmall S.H., McCullagh B., Branagan P., McElvaney N.G., Logan P.M. Radiological abnormalities associated with Aspergillus colonization in a cystic fibrosis population. Eur. J. Radiol., 2012, Vol. 81, no. 3, pp. e197-202.
 Oppenheimer J., Nelson H.S. Skin testing. Ann. Allergy Asthma Immunol., 2006, Vol. 96, no. 2, pp. 6-12.
Oppenheimer J., Nelson H.S. Skin testing. Ann. Allergy Asthma Immunol., 2006, Vol. 96, no. 2, pp. 6-12.23. Peetermans M., Goeminne P., de Boeck C., Dupont L.J. IgE sensitization to Aspergillus fumigatus is not a bystander phenomenon in cystic fibrosis lung disease. Chest, 2014, Vol. 146, pp. 99-100.
24. Sabino R., Ferreira J.A.G., Moss R.B., Valente J., Veríssimo C., Carolino E., Clemons K.V., Everson C., Banaei N., Penner J., Stevens D.A. Molecular epidemiology of Aspergillus collected from cystic fibrosis patients. J. Cyst. Fibros., 2015, Vol. 14, pp. 474-481.
25. Stevens D. A., Moss R.B., Kurup V.P., Knutsen A.P., Greenberger P., Judson M.A., Denning D.W., Crameri R., Brody A.S., Light M., Skov M., Maish W., Mastella G. Participants in the Cystic Fibrosis Foundation Consensus Conference. Allergic bronchopulmonary aspergillosis in cystic fibrosis – state of the art: Cystic Fibrosis Foundation Consensus Conference. Clin. Infect. Dis., 2003, Vol. 37, Suppl. 3, pp. 225-264.
Allergic bronchopulmonary aspergillosis in cystic fibrosis – state of the art: Cystic Fibrosis Foundation Consensus Conference. Clin. Infect. Dis., 2003, Vol. 37, Suppl. 3, pp. 225-264.
26. Williams C., Ranjendran, R., Ramage G. Pathogenesis of fungal infections in cystic fibrosis. Curr. Fungal Infect. Rep., 2016, Vol. 10, pp. 163-169.
Психологос — Я солнышко Автор теста – Н.И. Козлов Милые…
КАКИЕ ЖЕНЩИНЫ ПОЛУЧАЮТ СЕКСУАЛЬНОЕ УДОВОЛЬСТВИЕ?
• Насколько верен совет матери Вероники Франко (в прошлом куртизанки), которые она дает дочери, обучая ее искусству соблазнения («Чтобы дарить наслаждение, его надо ощущать»?
• Как раскрыть женскую сексуальность?
• Почему сексуальные способности женщины действуют только, когда они наложены на ее интеллектуальные способности?
• Почему практики, дающиеся на многих тренингах и курсах по раскрытию сексуальности, не работают?
• Как соотносится женский интеллект с ее сексуальностью?
• Как развить свои кинестетические способности?
• Почему японские женщины воспринимаются как сексуальные?
****
Чтобы получить ответы на эти вопросы, смотрите отрывок из моего двухчасового интервью-киноразбора с профессионалом высокого класса, психологом, экспертом в сфере сложных отношений, преподавателем Университета Практической Психологии, ЭМИЛИЕЙ ВАСИЛЬЕВНОЙ ГОНЧАРОВОЙ:
СЕКРЕТЫ женской мудрости и влюбления в фильме «ЧЕСТНАЯ КУРТИЗАНКА». Разбор и комментарии. (Э. Гончарова и А. Литовкина)
Разбор и комментарии. (Э. Гончарова и А. Литовкина)
***
Это интервью только для девочек! Мужчин не подпускайте к нему ни на шаг!
Это просто ВОСТОРГ, КРУТО, СНОГСШИБАТЕЛЬНО!
Забейте на работу и на учебу!
Берите отгул или академку!
Забейте на семью!
Скорее смотрите!
Жалко, у нас в Будапеште все заведения закрыты, и комендантский час с 20.00-5.00! И по городу нужно в маске ходить! А то бы я все техники соблазнения, которыми со мной поделилась Эмилия Васильевна во время нашего двухчасового интервью-киноразбора, сразу бы пустила в ход! 😂
Только тем, кто занимается спортом, можно ходить без маски! Так что остается только кататься на велосипеде без маски и с велосипеда соблазнять проходящих по велосипедной дорожке в масках мужчин!
*********************************
Из этого интервью вы получите подсказки на следующие вопросы, например:
• Как правильно заключать брак, по расчету или по любви? Какие еще существуют альтернативы?
• Как сказать своему любимому мужчине, что вы хотите за него замуж?
• Насколько целесообразно в наши дни пользоваться следующими советами матери Вероники Франко (в прошлом куртизанка), которые она дает дочери, обучая ее искусству соблазнения («Главный секрет куртизанок. Чтобы дарить наслаждение, его надо ощущать.»; «Заставь его поверить, что он единственный мужчина во вселенной»; «Люби любовь, а не люби мужчин, а то иначе окажешься в их власти»)?
Чтобы дарить наслаждение, его надо ощущать.»; «Заставь его поверить, что он единственный мужчина во вселенной»; «Люби любовь, а не люби мужчин, а то иначе окажешься в их власти»)?
• Как женщине научиться не привязываться к мужчине?
• Какие секреты соблазнения использовала Вероника, чтобы разжечь в Марко неугасающий интерес?
• Если бы Вероника не пошла в куртизанки, и вышла бы замуж за Марко, была бы она счастлива с ним в браке?
• Чему могут поучиться современные девушки у Вероники в контексте поиска наставника и выстраивания с ним (с ней) отношений?
• Какими приемами соблазнения пользовалась честная куртизанка, чтобы заинтересовать самых знатных жителей Венеции (и даже самого французского короля) и преуспеть на поприще куртизанки?
• Что нам, женщинам живущим в 21 веке, показывают Вероника как личность и ее жизненный путь?
• Как современные женщины могли бы использовать в наше время техники Вероники?
• Как женщине научиться привносить свою личность в отношения и с ее мужчиной, и с социумом?
(…и многие другие вопросы).
👉🏻 https://youtu.be/TZCTSYR5R5g
***
Эмилия Васильевна запускает ЖЕНСКУЮ СТАТУСНУЮ ДИСТАНЦИЮ! Мы про это тоже говорим в интервью.
Девочки, присоединяйтесь к нам!
Девиз женской ДИСТАНЦИИ:
«ХОРОШИЕ ДЕВОЧКИ БЕРУТ, ЧТО ДАЮТ, А СМЕЛЫЕ ДЕВОЧКИ ПОЛУЧАЮТ ТО, ЧТО ХОТЯТ».
Старт Дистанции: 17 февраля.
ДИСТАНЦИЯ будет проходить в среду, с 19 – 21 00.
https://www.univer-pp.ru/distantsiya#status
Автор: Anna Litovkina
Козлова Наталья Владимировна
Родилась в Ростовской области.
В настоящее время проживаю в г. Ростове-на-Дону.
В 2001 году окончила Барило-Крепинскую среднюю школу.
В 2007г. окончила Южный федеральный университет.
С декабря 2016 года работаю в ДГТУ в отделе довузовской подготовки заведующей сектором. – Организация и обеспечение работы отдела в соответствии с целями и задачами.
Образование
2005
Ростовский государственный педагогический университет, бакалавр социально-экономического образования по направлению «Социально-экономическое образование», профиль подготовки «История»
2007
Южный федеральный университет, магистр социально-экономического образования по направлению подготовки «Социально-экономическое образование», магистерская программа «Историческое образование»
2011
диссертационный совет при Южном федеральном университете, присуждена ученая степень кандидата исторических наук
Тесты для средней школы | Votum
Русский язык
- Звуки языка.
 Гласные и согласные звуки, учебник — Бунеев Р.Н., Бунеева Е.В. — скачать тест (Составитель: учитель русского языка и литературы Старикова Е. Ю., г. Волгоград).
Гласные и согласные звуки, учебник — Бунеев Р.Н., Бунеева Е.В. — скачать тест (Составитель: учитель русского языка и литературы Старикова Е. Ю., г. Волгоград). - Орфограмма. Опознавательный признак орфограммы. Условия выбора орфограммы, учебник — Бунеев Р.Н., Бунеева Е.В. — скачать тест (Составитель: учитель русского языка и литературы Старикова Е. Ю., г. Волгоград).
- Буквы о, ё после шипящих в корне слова, учебник — Бунеев Р.Н., Бунеева Е.В. — скачать тест (Составитель: учитель русского языка и литературы Старикова Е. Ю., г. Волгоград).
- Правописание слов с чередующимися гласными а-о в корне, учебник — Бунеев Р.Н., Бунеева Е.В. — скачать тест (Составитель: учитель русского языка и литературы Старикова Е. Ю., г. Волгоград).
- Синтаксис и пунктуация. Словосочетание. скачать (Научный руководитель Бобровская Л.Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).
- Синтаксис и пунктуация. Второстепенные члены предложения. Дополнение. скачать тест (Научный руководитель Бобровская Л.Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).
- Синтаксис и пунктуация. Предложение. скачать тест (Научный руководитель Бобровская Л.Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).
- Синтаксис и пунктуация. Главные члены предложения. Подлежащее. скачать тест (Научный руководитель Бобровская Л.Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).
- Синтаксис и пунктуация. Тире между подлежащим и сказуемым. скачать тест (Научный руководитель Бобровская Л.Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).
- Синтаксис и пунктуация. Определение. скачать тест (Научный руководитель Бобровская Л. Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).
- Синтаксис и пунктуация. Обстоятельство. скачать тест (Научный руководитель Бобровская Л.Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).
 Гласные и согласные звуки, учебник — Бунеев Р.Н., Бунеева Е.В. — скачать тест (Составитель: учитель русского языка и литературы Старикова Е. Ю., г. Волгоград).
Гласные и согласные звуки, учебник — Бунеев Р.Н., Бунеева Е.В. — скачать тест (Составитель: учитель русского языка и литературы Старикова Е. Ю., г. Волгоград). Второстепенные члены предложения. Дополнение. скачать тест (Научный руководитель Бобровская Л.Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).
Второстепенные члены предложения. Дополнение. скачать тест (Научный руководитель Бобровская Л.Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).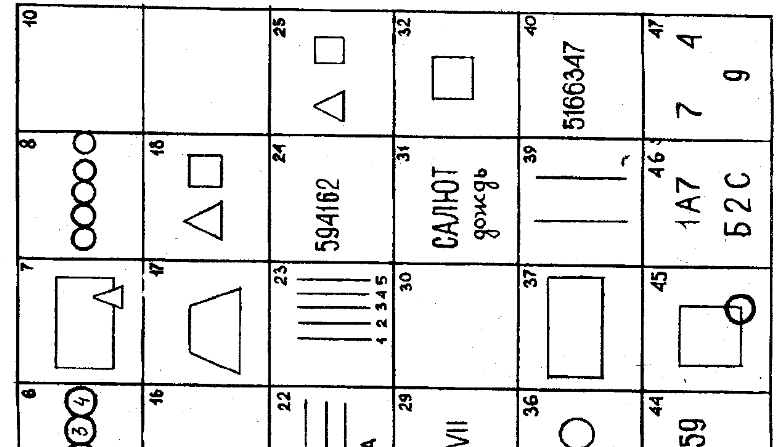 Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).
Н., ГБОУ ДПО ВГАПКиПРО. Составители:учителя русского языка и литературы Акентьева Е.И. и Ковалев С.А., г. Волгоград).Математика
- Десятичная система счисления (часть 1), учебник — И.И Зубарева, А. Г. Мордкович — скачать тест (Составитель: учитель математики Ялунина А. С., г. Волгоград).
- Десятичная система счисления (часть 2), учебник — И.И Зубарева, А. Г. Мордкович — скачать тест (Составитель: учитель математики Ялунина А. С., г. Волгоград).
- Ряд натуральных чисел, учебник — Дорофеев Г.В. — скачать тест (Составитель: учитель математики Медведева С.А, г. Волгоград).
- Обыкновенные дроби, математический диктант, скачать тест (Составитель: учитель математики Лобанова О.Е., г. Саранск).
Тесты по учебнику Виленкин Н.Я., Жохов В.И., Составители: учитель математики Лопатина Т.А., Шишкина Л.Е., г. Волгоград:
- Буквенная запись свойств сложения и вычитания. Упрощение выражений. Выбор ответа. Скачать тест
- Определение разрядов и классов заданных чисел. Ввод ответа. Скачать тест
- Определение разрядов и классов заданных чисел. Выбор ответа. Скачать тест
- Сложение натуральных чисел с помощью координатного луча. Ввод ответа. Скачать тест
- Сложение натуральных чисел в различных величинах. Ввод ответа. Скачать тест
- Сложение натуральных чисел и его свойства. Выбор ответа. Скачать тест
- Сложение натуральных чисел и его свойства. Соревнование. Скачать тест
- Сложение двузначных чисел. Устный счет. Вариант 1 Скачать тест
- Сложение двузначных чисел. Устный счет. Вариант 2 Скачать тест
- Вычисление суммы чисел наиболее рациональным способом. Соревнование. Вариант 1. Скачать тест
- Вычисление суммы чисел наиболее рациональным способом. Соревнование. Вариант 2. Скачать тест
- Сложение натуральных чисел. Решение задач на сложение. Ввод ответа. Скачать тест
- Вычитание натуральных чисел. Ввод ответа. Скачать тест
- Вычитание натуральных чисел. Задачи. Ввод ответа. Скачать тест
- Натуральные числа. Сравнение. Ввод ответа. Скачать тест
- Натуральные числа. Выбор наименьшего числа. Ввод ответа. Скачать тест
- Натуральные числа. Выбор наибольшего числа. Ввод ответа. Скачать тест
- Натуральные числа. Упорядочивание ряда чисел. Ввод ответа. Скачать тест
- Вычитание натуральных чисел. Соревнование. Скачать тест
- Текстовые задачи на нахождение длин частей отрезка. Ввод ответа. Скачать тест
- Текстовые задачи на нахождение периметра многоугольника. Ввод ответа. Скачать тест
- Числовые и буквенные выражения. Вычисление выражений. Ввод ответа. Скачать тест
- Числовые и буквенные выражения. Составление выражений для решения задачи. Выбор ответа. Скачать тест
- Числовые и буквенные выражения. Решение задач Скачать тест
- Буквенная запись свойств сложения и вычитания. Упрощение выражений. Выбор ответа. Вариант 1. Скачать тест
- Буквенная запись свойств сложения и вычитания. Упрощение выражений. Выбор ответа. Вариант 2. Скачать тест
- Решение уравнений с выбором варианта. Скачать тест
- Уравнения. Решение уравнений. Ввод ответа. Скачать тест
- Решение уравнений с введением ответа. Скачать тест
- Решение уравнений с использованием свойств сложения и вычитания. Скачать тест
- Уравнения. Нахождение уравнения по его корню. Скачать тест
- Уравнения. Решение задач с помощью уравнений. Ввод ответа. Скачать тест
- Уравнения. Составление уравнений по условию задачи. Выбор ответа. Скачать тест
 Соревнование. Вариант 1. Скачать тест
Соревнование. Вариант 1. Скачать тест Вычисление выражений. Ввод ответа. Скачать тест
Вычисление выражений. Ввод ответа. Скачать тест Скачать тест
Скачать тест
Английский язык
- Present Continuos скачать тест (Составитель: Мальцева Л.С. — учитель английского языка, г. Волгоград).
- Present Simple скачать тест (Составитель: Мальцева Л.С. — учитель английского языка, г. Волгоград).
- Present Simple скачать тест (Составитель: Пасимишина В. — учитель английского языка, г. Волгоград).
- Present Simple, часть 2, скачать тест (Составитель: Пасимишина В. — учитель английского языка, г. Волгоград).
- Present Simple, часть 3, выбор между do и does, скачать тест (Составитель: Пасимишина В. — учитель английского языка, г. Волгоград).
- Present Simple, часть 4, скачать тест (Составитель: Пасимишина В. — учитель английского языка, г. Волгоград).
- Глагол To be, скачать тест (Составитель: Пасимишина В. — учитель английского языка, г. Волгоград).
- Lexical lesson house and furniture, скачать тест (Составитель: Мальцева Л. С. — учитель английского языка, г. Волгоград).
 С. — учитель английского языка, г. Волгоград).
С. — учитель английского языка, г. Волгоград).Информатика
- Проверочная работа «Параметры шрифта и абзаца», учебник — Л.Л.Босова, скачать тест (составитель: учитель информатики Аршинова О.Н.).
- Устройство компьютера, учебник Босова Л.Л., скачать тест (учитель физики и информатики Галкин С.И., Нижегородская область).
- Информация вокруг нас, учебник Босова Л.Л., скачать тест (учитель физики и информатики Галкин С.И., Нижегородская область).
- «Устройство компьютера» учебник Босова Л.Л., скачать тест (Составитель: Рассохина Г.В., учитель информатики, г. Саранск).
История
- Установление господства Рима во всем Средиземноморье, скачать тест (Составитель: Гриневич И.Б., учитель истории и обществознания, г. Фролово, Волгоградская обл.).
Психологические тесты
- Диагностика структуры учебной мотивации школьника, методика М. В. Матюхиной, скачать тест
- Тест для родителей «Уровень агрессивности ребёнка», Лаврентьева Г.П., Титаренко Т.М, скачать тест
- Диагностика принятия других, В. Фей, скачать тест
- Методика изучения уровня внимания у школьников (Гальперин П.Я., Кабылицкая С.Л.), скачать тест
- Тест «Избирательность внимания», 4 класс (Тест Г.Мюнстерберга), скачать тест
- Тест «Настойчивость» (Методика Е.П.Ильина, Е.К.Фещенко), скачать тест
- Тест «Упорство» (Методика Е.П.Ильина, Е.К.Фещенко), скачать тест
- Методика для отслеживания некоторых аспектов нравственного развития младших и средних школьников, З. Гильбух, Опросник «Мой класс», скачать тест
- Методика «Родителей оценивают дети», И.А.Фурманов и А.А.Аладьин, скачать тест
- Уровень и тип чувства юмора Домбровская И.С., скачать тест
- «Анализ семейных взаимоотношений» Э.Г. Эйдемиллер, В.В. Юстицкий (АСВ), скачать тест
- Измерение родительских установок и реакций (РАКУ; Т. Архиреева), для родителей, скачать тест
- Опросник для родителей «Типы воспитания детей», для родителей, скачать тест (Шапарь В.Б. Практическая психология. Психодиагностика отношений между родителями и детьми).
 В. Матюхиной, скачать тест
В. Матюхиной, скачать тест Архиреева), для родителей, скачать тест
Архиреева), для родителей, скачать тестТест Розенцвейга, интерпретация Н.Козлова
Дано двенадцать картинок теста и вам нужно обычным образом дать реплики для озадаченного персонажа. Ответы лучше давать не размышляя над тем, что вы говорите, (если именно так вы среди людей и живете), но можете одновременно включать и голову, если вы к этому приучены и вас это не затрудняет.
Для самых продвинутых и недовольных любыми ограничениями разрешается давать даже несколько ответов — в конце концов, у вас же бывают разные настроения!
| 1) – Как обидно, что моя машина сломалась, и вы из-за этого опоздали на поезд! | |
| 2) – Вам же не видно экрана! | |
| 3) – Простите пожалуйста, значит нас неправильно соединили! | |
| 4) – Она должна была быть здесь уже десять минут назад! | |
| 5) – Вот ваша газета, мне очень жаль, что ребенок ее разорвал! | |
6) – Я понимаю, что подвожу тебя, но приехать я не смогу. | |
| 7) – Это тетя Наташа, она просит нас немного подождать, пока она не придет и еще раз не пожелает нам доброго пути! | |
| 8) – Вы не ушиблись? | |
| 9) – Как нарочно, теперь ты потерял ключи. | |
| 10) – Вот уже третий раз я прихожу к вам с этими часами. Я их купила всего неделю назад, но как только прихожу домой, они останавливаются. | |
| 11) – Мне очень жаль, но мы как раз продали последний экземпляр! | |
| 12) – Твоя подруга пригласила меня сегодня вечером на танцы. Она сказала, что ты не пойдешь. |
Ответы на тест Розенцвейга
Связи с усилием контроля ребенка, личностью и проблемным поведением — Новосибирский государственный университет
TY — JOUR
T1 — Тест на размер животного, похожий на Stroop
T2 — Связи с усилием контроля ребенка, личностью и проблемным поведением
AU — Баирова, Надежда Б.
AU — Бочаров Андрей Викторович
AU — Савостьянов Александр Николаевич
AU — Петренко Евгения Н.
AU — Козлова Елена Александровна
AU — Сапригин Александр Е.
AU — Слободская, Елена Р.
PY — 2020/4/2
Y1 — 2020/4/2
N2 — В этом исследовании изучались результаты детей в тесте размера животного Струпа и его связь с родителями. сообщили о контроле над темпераментом, личности и общих эмоциональных и поведенческих проблемах в российской выборке детей 5–12 лет (N = 202). Тест на размер животных продемонстрировал эффект Струпа в отношении точности и времени реакции (RT) для обоих полов и всех возрастов.Успеваемость детей в тесте на размер животных с возрастом значительно улучшалась: дети старшего возраста выполняли задания более точно, быстрее и их ответы были менее вариабельными, чем ответы детей младшего возраста. Полученные данные показали, что RT отрицательно связана с чертой темперамента, требующей усилий, контролем и ее ключевыми компонентами, фокусировкой внимания и тормозящим контролем, в то время как вариабельность RT отрицательно связана как с характеристиками регуляции, контролем с усилием и сознательностью, так и с их компонентами. Успеваемость детей на AST была значимо связана с гиперактивностью-невниманием, экстернализирующим поведением и общим уровнем детской психопатологии. Эти результаты подтверждают полезность теста на размер животных в нейропсихологической оценке детей дошкольного и школьного возраста.
Успеваемость детей на AST была значимо связана с гиперактивностью-невниманием, экстернализирующим поведением и общим уровнем детской психопатологии. Эти результаты подтверждают полезность теста на размер животных в нейропсихологической оценке детей дошкольного и школьного возраста.
AB — В этом исследовании изучались результаты детей в тесте на размер животного, подобного Струпу, и его связь с родительским контролем над темпераментом, личностью и общими эмоциональными и поведенческими проблемами в российской выборке детей в возрасте 5-12 лет (N = 202).Тест на размер животных продемонстрировал эффект Струпа в отношении точности и времени реакции (RT) для обоих полов и всех возрастов. Успеваемость детей в тесте на размер животных с возрастом значительно улучшалась: дети старшего возраста выполняли задания более точно, быстрее и их ответы были менее вариабельными, чем ответы детей младшего возраста. Полученные данные показали, что RT отрицательно связана с чертой темперамента, требующей усилий, контролем и ее ключевыми компонентами, фокусировкой внимания и тормозящим контролем, в то время как вариабельность RT отрицательно связана как с характеристиками регуляции, контролем с усилием и сознательностью, так и с их компонентами. Успеваемость детей на AST была значимо связана с гиперактивностью-невниманием, экстернализирующим поведением и общим уровнем детской психопатологии. Эти результаты подтверждают полезность теста на размер животных в нейропсихологической оценке детей дошкольного и школьного возраста.
Успеваемость детей на AST была значимо связана с гиперактивностью-невниманием, экстернализирующим поведением и общим уровнем детской психопатологии. Эти результаты подтверждают полезность теста на размер животных в нейропсихологической оценке детей дошкольного и школьного возраста.
кВт — управление усилиями
кВт — индивидуальность
кВт — проблемное поведение
кВт — самоконтроль
кВт — тест размера животного
кВт — ЭФФЕКТИВНОСТЬ
кВт —
ТЕНДЕНЦИЙ, СВЯЗАННЫХ С ВОЗРАСТОМ 9000 КВТ — САМОРЕГУЛИРОВАНИЕКВТ — НАРУШЕНИЯ АУТИЗМНОГО СПЕКТРА
КВТ — ВРЕМЕННАЯ РЕАКЦИЯ ВАРИАБЕЛЬНОСТЬ
КВТ — КОНТРОЛЬ ПОМЕХ
КВ — ИСПОЛНИТЕЛЬНАЯ ФУНКЦИЯ
КВТ — ИНДИВИДУАЛЬНОЕ УПРАВЛЕНИЕ
КВТ
КВТ — ИНДИВИДУАЛЬНОЕ УПРАВЛЕНИЕ
КВТ — ИНДИВИДУАЛЬНОЕ УПРАВЛЕНИЕ
КВТ — ИНДИВИДУАЛЬНОЕ УПРАВЛЕНИЕ
КВТ ДЕТИ
UR — http: // www.scopus.com/inward/record.url?scp=85073813762&partnerID=8YFLogxK
U2 — 10. 1080 / 09297049.2019.1665173
1080 / 09297049.2019.1665173
DO — 10.1080 / 09297049.2019.1665173
M3 — артикул
0003M3
VL — 26
SP — 409
EP — 432
JO — Детская нейропсихология
JF — Детская нейропсихология
SN — 0929-7049
IS — 3
ER Пустые мезопористые частицы
замедляет прогрессирование заболевания и увеличивает выживаемость на мышиной модели ALSСинтез мезопористого диоксида кремния
Pluronic 123 (трехблочный сополимер, EO20PO70EO20, Sigma-Aldrich) (4 г) в качестве агента-шаблона и триметилбензол (ТМБ) (мезитилен, Sigma-Aldrich) (3.3 г) в качестве агента набухания растворяли в 127 мл дистиллированной H 2 O и 20 мл соляной кислоты (HCl, 37%, Sigma-Aldrich) при перемешивании при комнатной температуре (RT) в течение 3 дней. Раствор предварительно нагревали до 40 ° C перед добавлением 9,14 мл TEOS (тетраэтилортосиликат, Sigma-Aldrich). Смесь перемешивали еще 10 мин со скоростью 500 об / мин и выдерживали при 40 ° C в течение 24 часов, затем подвергали гидротермической обработке в печи при 100 ° C еще 24 часа. Наконец, смесь фильтровали, промывали и сушили при комнатной температуре.Продукт прокаливали для удаления шаблона поверхностно-активного вещества и агента набухания. Прокаливание проводили путем нагревания до 600 ° C со скоростью нагрева 1,5 ° C / мин и выдерживали при 600 ° C в течение 6 часов с последующим охлаждением до условий окружающей среды. Полученный продукт представлял собой белый порошок, состоящий из нанопористых частиц диоксида кремния.
Наконец, смесь фильтровали, промывали и сушили при комнатной температуре.Продукт прокаливали для удаления шаблона поверхностно-активного вещества и агента набухания. Прокаливание проводили путем нагревания до 600 ° C со скоростью нагрева 1,5 ° C / мин и выдерживали при 600 ° C в течение 6 часов с последующим охлаждением до условий окружающей среды. Полученный продукт представлял собой белый порошок, состоящий из нанопористых частиц диоксида кремния.
Физико-химические характеристики мезопористого диоксида кремния
Мезопористые частицы диоксида кремния были охарактеризованы ранее 60 . Загрузочное количество пептида определяли термогравиметрическим анализом (PerkinElmer, Waltham, MA, USA).Температурное изменение было выполнено от 20 ° C до 900 ° C при скорости нагрева 20 ° C / мин. Атмосфера подключаемого газа представляла собой сухой воздух (расход 20 мл / мин). Масса навески варьировала от 5 до 10 мг.
Изотерма адсорбции-десорбции азота была проведена для характеристики пористости и определения площади поверхности порошков MSP (прибор Micromeritics Tristar II 3020, Норкросс, Джорджия, США). Прокаленные MSP дегазировали при 300 ° C в течение 6 ч в токе газообразного азота. Образцы, нагруженные трофическим фактором, дегазировали при 60 ° C в течение 48 ч перед анализом.Распределение пор по размерам рассчитывали с использованием модели Density Functional Theory (DFT), предполагая цилиндрическую геометрию пор, с использованием пакета программного обеспечения, входящего в комплект Micromeritics Instrument (рис. 1) 34 .
Прокаленные MSP дегазировали при 300 ° C в течение 6 ч в токе газообразного азота. Образцы, нагруженные трофическим фактором, дегазировали при 60 ° C в течение 48 ч перед анализом.Распределение пор по размерам рассчитывали с использованием модели Density Functional Theory (DFT), предполагая цилиндрическую геометрию пор, с использованием пакета программного обеспечения, входящего в комплект Micromeritics Instrument (рис. 1) 34 .
Изображения, полученные с помощью сканирующей электронной микроскопии (СЭМ), были записаны с помощью СЭМ LEO 1550, оборудованного автоэмиссионной пушкой Шоттки, работающей при ускоряющем напряжении 1–3 кВ и увеличении 20 000–50 000 × на образцах без золотого покрытия. Просвечивающая электронная микроскопия (ПЭМ) кальцинированных образцов была проведена на микроскопе JEOL-3010 (JEOL), работающем при 300 кВ (Cs, 0.6 мм; Разрешение 1,7 Å). Изображения были записаны с помощью камеры с зарядовой связью (CCD) (Keen View, SIS Analysis Specialized Imaging, Olympus Soft Imaging Solutions, Мюнстер, Германия; 1024 × 1024 пикселей; поле зрения 23,5 × 23,5 мкм) при увеличении 30 000–100 000 × с использованием условий низкой дозы на синтезированных и прокаленных образцах 34 .
Функционализация и маркировка мезопористых наночастиц диоксида кремния
Чтобы привить изотиоцианат родамина B на поверхность частиц диоксида кремния, частицы диоксида кремния сначала функционализировали пропиламиновыми группами.Кальцинированный MSP (1 г) добавляли к толуолу (50 мл) и обрабатывали ультразвуком в течение 30 минут. (3-аминопропил) триэтоксисилан (APES, Sigma-Aldrich) (650 мкл) растворяли в толуоле (5 мл) при комнатной температуре и добавляли к суспензии диоксида кремния. Смесь перемешивали в течение ночи, фильтровали, промывали этанолом и сушили на воздухе. Полученные в результате функционализированные аминопропилом MSP были названы нанопористый диоксид кремния-NH 2 . Термогравиметрический анализ (ТГА) использовали для определения количества аминопропильных групп, привитых на поверхность диоксида кремния.Потеря веса функционализированных частиц диоксида кремния в диапазоне температур 100–900 ° C, связанная с органическими пропиламиновыми группами, составила около 11%.
Изотиоцианат родамина B (Rh, Sigma-Aldrich) использовали для мечения частиц диоксида кремния. Rh (1 мг) растворяли в метаноле (100 мл) и добавляли к 1 г нанопористого диоксида кремния-NH 2 . Смесь перемешивали при 40 ° C в течение 2 ч, фильтровали, промывали этанолом и сушили на воздухе. Окончательные образцы были названы «Нанопористый кремнезем-Rh.
Площадь поверхности и пористость нанопористого кремнезема-Rh измеряли методом адсорбции-десорбции N 2 (рис. 1). Площадь поверхности функционализированного кремнезема уменьшилась с 722 до 402 м 2 2 / г по сравнению с нефункционализированным кремнеземом, в то время как размер пор незначительно уменьшился с 23 до 21,5 нм, что указывает на то, что пористость частиц остается доступной для загрузки пептидных миметиков. . Флуоресцентные свойства меченых частиц визуализировали с помощью флуоресцентной микроскопии и выявляли очень сильный флуоресцентный сигнал (данные не показаны).
Пептидные миметики нейротропного фактора
Пептиды Глиафин (ETMYDKILKNLSRSR) 41 , Цинтрофин (DGGLFEKKLWGLKV) 42 и Vefin1 (AKFMDVYQRSYSHA, состоящие из четырех моноаффинов, синтезированных на 90-концевых концах) 44 Твердофазный пептидный синтез Fmoc (Schafer-N, Копенгаген, Дания). Чистота пептида составляла ≥ 95%, как было определено с помощью высокоэффективной жидкостной хроматографии.
Чистота пептида составляла ≥ 95%, как было определено с помощью высокоэффективной жидкостной хроматографии.
Загрузка мезопористых частиц диоксида кремния пептидными миметиками
Цинтрофин, Глиафин и Вефин1 загружали в мезопоры путем пропитки водой, как описано ранее 34 .Вкратце, MSP добавляли к водным растворам каждого пептида и перемешивали при 4 ° C в течение 16 часов с последующим выпариванием воды в атмосферных условиях. Количество загружаемых пептидов определяли термогравиметрическим анализом (PerkinElmer, Waltham, MA, USA) путем сканирования образцов от 20 до 900 ° C при скорости нагревания 20 ° C / мин. Заполняемая газовая атмосфера представляла собой сухой воздух со скоростью потока 20 мл / мин. Загружаемые количества Цинтрофина, Глиафина и Вефина1 составляли 8,3 мас.%, 11,8 мас.% И 8 мас.% Соответственно. Соответственно, данные изотермы адсорбции показали значительное уменьшение объема пор и размера пор MSP после включения миметиков.Порошковые рентгеновские дифрактограммы загруженных образцов показали, что пептиды были загружены в аморфном состоянии, поскольку дифракционные пики не наблюдались. Кинетика высвобождения мезопористых частиц диоксида кремния, нагруженных пептидами, была продемонстрирована ранее 34 .
Кинетика высвобождения мезопористых частиц диоксида кремния, нагруженных пептидами, была продемонстрирована ранее 34 .
Культура мотонейронов, полученных из эмбриональных стволовых клеток мыши (mESC), для тестирования нейротрофической активности Vefin1 in vitro
Поскольку нейротрофическая эффективность пептидного миметика Vefin1 на МН ранее не была задокументирована, мы создали ту же систему in vitro, что и ранее использовался для Цинтрофина и Глиафина 21 .Линия ESC мыши, несущая GFP-кодируемые гены под контролем специфического для MN фактора транскрипции HB9 (HB9: GFP) (подарок д-ра Кевина Эггана, Гарвардский институт стволовых клеток), была напрямую дифференцирована in vitro, как описано ранее 61 . Было проведено три независимых эксперимента. Вкратце, mESC размножали поверх эмбриональных фибробластов мыши в среде для культивирования клеток ES (Embryonic Stem), содержащей 82% KoDMEM с высоким содержанием глюкозы, 1% GlutaMAX, 1% пенициллин-стрептомицин, 1% заменимых аминокислот (все от Life Technologies , Карлсбад, США), 14% заменитель сыворотки KnockOut (KoSR, Invitrogen, Eugene, USA), 100 мкМ бета-меркаптоэтанол (ME) и 1% фактор ингибирования лейкемии ESGRO (LIF, Millipore, Billerica, США). Чтобы инициировать дифференцировку нейронов, диссоциированные mESC повторно суспендировали в среде ADFNB, содержащей Advanced DMEM / F12: Neurobasal (1: 1 об. / Об.), 0,5% N2, 1% B27 (все от Life Technologies), 1% GlutaMAX, 1% Пенициллин – стрептомицин, 1% KoSR и 100 мкМ бета-ME. Два дня спустя, когда клетки сформировали однородные эмбриоидные тельца (EB), среду заменили на свежий ADFNB с добавлением 0,1 мкМ ретиноевой кислоты (Sigma, Сент-Луис, США) и 0,2 мкМ агониста Shh Ag1.3 (Phoenix Pharmaceuticals; Burlingame, США). ), и клетки культивировали в течение пяти дней подряд, меняя среду через день.Для культур MN EB диссоциировали и высевали с плотностью 7,5 × 10 4 клеток / покровное стекло на 0,01% поли-1-орнитина и 10% ламинина (Sigma), предварительно покрытых покровными стеклами в среде ADFNB. Сразу после посева клетки стимулировали серийно разведенным пептидом Vefin1 в течение 24 часов. Обработанные и необработанные культуры затем фиксировали 4% формальдегидом и окрашивали 1: 10 000 Hoechst 33342 (Invitrogen, Eugene, USA). Прорастание нейритов из MN, экспрессирующих GFP, анализировали, как описано ранее 44 .
Чтобы инициировать дифференцировку нейронов, диссоциированные mESC повторно суспендировали в среде ADFNB, содержащей Advanced DMEM / F12: Neurobasal (1: 1 об. / Об.), 0,5% N2, 1% B27 (все от Life Technologies), 1% GlutaMAX, 1% Пенициллин – стрептомицин, 1% KoSR и 100 мкМ бета-ME. Два дня спустя, когда клетки сформировали однородные эмбриоидные тельца (EB), среду заменили на свежий ADFNB с добавлением 0,1 мкМ ретиноевой кислоты (Sigma, Сент-Луис, США) и 0,2 мкМ агониста Shh Ag1.3 (Phoenix Pharmaceuticals; Burlingame, США). ), и клетки культивировали в течение пяти дней подряд, меняя среду через день.Для культур MN EB диссоциировали и высевали с плотностью 7,5 × 10 4 клеток / покровное стекло на 0,01% поли-1-орнитина и 10% ламинина (Sigma), предварительно покрытых покровными стеклами в среде ADFNB. Сразу после посева клетки стимулировали серийно разведенным пептидом Vefin1 в течение 24 часов. Обработанные и необработанные культуры затем фиксировали 4% формальдегидом и окрашивали 1: 10 000 Hoechst 33342 (Invitrogen, Eugene, USA). Прорастание нейритов из MN, экспрессирующих GFP, анализировали, как описано ранее 44 .
Культура двигательных нейронов
Первичные культуры двигательных нейронов получали, как описано ранее 62 . Моторные нейроны выделяли и помещали на восьмилуночные предметные стекла LabTek Permanox (Nunc, Roskilde, Дания), предварительно покрытые поли-L-орнитином и ламинином. Клетки культивировали в течение 7–10 дней в нейробазальной среде с добавлением 2% B27, 2% (об. / Об.) Лошадиной сыворотки, глютамина 100 Ед / мл пенициллина и 100 мкг / мл стрептомицина. Чтобы вызвать гибель клеток, клетки обрабатывали H 2 O 2 (60 мкМ; Sigma-Aldrich) отдельно или вместе с 10 мкл пустых MSP в течение 48 часов.Для каждого индивидуального эксперимента лунку с нестимулированными клетками использовали в качестве положительного контроля. Супернатант собирали, центрифугировали при 12000 × g и уровень глутамата оценивали с помощью набора для анализа глутамата (Sigma). Клетки фиксировали 4% формальдегидом и морфологию ядра визуализировали с помощью Hoechst 33258 (1: 1000, Invitrogen). Случайные изображения регистрировали с помощью микроскопа Zeiss Axiovert 100, установленного с камерой AxioCam MRm (Zeiss), и выживаемость клеток оценивалась, как описано ранее 44 .
Клетки фиксировали 4% формальдегидом и морфологию ядра визуализировали с помощью Hoechst 33258 (1: 1000, Invitrogen). Случайные изображения регистрировали с помощью микроскопа Zeiss Axiovert 100, установленного с камерой AxioCam MRm (Zeiss), и выживаемость клеток оценивалась, как описано ранее 44 .
Культура стволовых клеток пограничного капсюля нервного гребня (bNCSCs)
bNCSC были получены от трансгенных мышей, несущих красный флуоресцентный белок (RFP) под универсальным промотором актина 63 согласно ранее опубликованным протоколам 64,65 . Вкратце, DRG вместе с граничными колпачками были механически отделены от изолированного спинного мозга и механо-ферментативно диссоциированы с использованием коллагеназы / диспазы (1 мг / мл) и ДНКазы (0,5 мг / мл) в течение 30 минут при комнатной температуре.Клетки высевали при 0,5–1 × 10 5 клеток / см 2 в среду N2, содержащую B27 (Gibco) с добавлением EGF и bFGF (каждый 20 нг / мл; R&D Systems (Абингдон, Великобритания). Через 12 лет h, неприлипающие клетки удаляли вместе с половиной среды, которую заменяли свежей средой.Среду меняли через день (50% среды заменяли свежей средой) до образования нейросфер (примерно две недели культивирования В последующих экспериментах использовали непассированные нейросферы в культуре от двух до трех недель.
Через 12 лет h, неприлипающие клетки удаляли вместе с половиной среды, которую заменяли свежей средой.Среду меняли через день (50% среды заменяли свежей средой) до образования нейросфер (примерно две недели культивирования В последующих экспериментах использовали непассированные нейросферы в культуре от двух до трех недель.
Животные
Всего 40 фоновых мышей B6 / SJL (B6SJL-TgN-SOD1-G93A-1Gur; SOD1 G93A ) были получены из лаборатории Джексона (штат Мэн, США), а 6 мышей NMRI были получены из Möllegaard и Bomholgard Breeding и Исследовательский центр (M&B A / S, Bomholt, Дания) использовался для экспериментов. Все эксперименты проводились в соответствии с Директивами Европейского Союза и Руководством Общества неврологии для исследований на животных. Были предприняты все усилия, чтобы минимизировать страдания животных.Для животных B6SJL-TgN-SOD1-G93A-1Gur протокол лечения был одобрен Комитетом по уходу и использованию животных Университета Бен-Гуриона в Негеве в соответствии с требованиями законодательства Израиля, а для животных NMRI одобрение было дано Упсальский комитет по этике исследований на животных.
Хирургия
Мышей перед операцией анестезировали 3% изофлураном в начале процедуры и поддерживали с течением времени 0,8% при скорости потока изофлурана 500–480 мл / мин. Кожа на дорсальной части шеи была надрезана, а левые позвоночные пластинки C3–4 обнажены и удалены, чтобы обнажить левую дорсальную часть соответствующих сегментов спинного мозга (дополнительный рис.S3A, B). Три инъекции (по 2 мкл каждая) были сделаны в вентральную часть на левой стороне шейного отдела спинного мозга (C3 – C4) с помощью шприца Hamilton объемом 10 мкл, присоединенного к микрошприцевому контроллеру насоса (Micro 4, World Precision Instruments, Сарасота, Флорида) (дополнительный рисунок S3C). Мышей SOD1 G93A оперировали в возрасте 79 дней (~ 10 недель) и разделили на три экспериментальные группы: (1) контрольная группа (n = 10; незагруженные (пустые) MSP), (2) группа пептидных миметиков (n = 10). = 10; MSP, нагруженные Cintrofin, Gliafin и Vefin1) и (3) группа bNCSC (n = 10; ~ 26000 клеток на инъекцию). Мышей NMRI (n = 6) оперировали в том же возрасте и таким же образом с инъекцией 2 мкл меченных родамином мезопористых наночастиц кремнезема (Rh-MSP). Мыши выздоровели без осложнений после операции (дополнительный рис. S3D).
Мышей NMRI (n = 6) оперировали в том же возрасте и таким же образом с инъекцией 2 мкл меченных родамином мезопористых наночастиц кремнезема (Rh-MSP). Мыши выздоровели без осложнений после операции (дополнительный рис. S3D).
Иммуногистохимия
Мышей NMRI повторно анестезировали через одну неделю (n = 3) и один месяц (n = 3) после операции и перфузировали теплым физиологическим раствором (~ 38 ° C) с последующим введением фиксирующего раствора, содержащего ледяной холод. 4% формальдегида в фосфатно-солевом буфере (PBS; ~ 4 ° C, pH 7.35–7.45). Сегменты спинного мозга C3–5 удаляли, затем фиксировали при 4 ° C в течение 4 часов и подвергали криозащите в течение ночи в PBS, содержащем 15% сахарозы. Серийные коронковые срезы (14 мкм) вырезали через спинной мозг на криостате и помещали на предметные стекла SuperFrost Plus (Menzel-Gläser, Брауншвейг, Германия). Срезы предварительно инкубировали с блокирующим раствором (1% бычьего сывороточного альбумина, 0,3% Triton X-100 и 0,1% NaN3 в PBS) в течение 1 ч при комнатной температуре, а затем инкубировали в течение ночи при 4 ° C с первичными антителами к глиальному фибриллярному кислому белку ( GFAP; кроличьи поликлональные, 1: 500, Dako) или антитела к IBA1 (микроглиальный маркер; козьи поликлональные. , 1: 500, Абкам). Иммуногистохимия выполнялась, как описано ранее 23,33 . После промывки PBS соответствующие вторичные антитела наносили на 4 часа при комнатной температуре, и ядра клеток метили Hoechst. Срезы исследовали с помощью конфокального микроскопа LSM700 (Zeiss).
, 1: 500, Абкам). Иммуногистохимия выполнялась, как описано ранее 23,33 . После промывки PBS соответствующие вторичные антитела наносили на 4 часа при комнатной температуре, и ядра клеток метили Hoechst. Срезы исследовали с помощью конфокального микроскопа LSM700 (Zeiss).
Оценка поведения
После операции мышей взвешивали ежедневно в течение всего периода наблюдения и проверяли на силу захвата два раза в неделю, как описано ранее. 66 , начиная с двух недель до операции.Время начала заболевания ретроспективно определяли как время, когда мыши достигли максимальной массы тела. Раннее заболевание определялось в то время, когда атрофия мышц, вызванная денервацией, приводила к потере 10% максимального веса. Конечная стадия была определена параличом настолько серьезным, что животное не могло выпрямиться в течение 20 секунд, когда помещалось на бок, конечная точка, часто используемая для мутантных мышей SOD1, и та, которая соответствует требованиям Комитета по уходу за животными и использованию животных Бена. -Университет Гуриона в Негеве.Проверка силы захвата проводилась с помощью прибора для измерения силы Chatillon (Ametek). Измерение каждого интегранта в группах состояло из двух измерений (натяжение передних конечностей и полное натяжение конечностей).
-Университет Гуриона в Негеве.Проверка силы захвата проводилась с помощью прибора для измерения силы Chatillon (Ametek). Измерение каждого интегранта в группах состояло из двух измерений (натяжение передних конечностей и полное натяжение конечностей).
Статистический анализ
Данные собирались на протяжении всего исследования и анализировались с помощью RStudio Team (2020) версии 1.3.959. (RStudio: интегрированная разработка для R. RStudio, PBC, Бостон, Массачусетс, URL http://www.rstudio.com/). Кривые Каплана-Мейера были построены с помощью пакета выживания v3.2-3 и построено с помощью Survminer package v0.4.7. Гистограммы и линейные графики были созданы с помощью пакета ggplot2 v3.3.2. Статистический анализ был дополнительно выполнен с использованием stats v3.6.3 (R Core Team) и других пакетов, загруженных в среду RStudio, dplyr v1.0.0, tidyverse v1.3.0. И multicomp v1.4–13 . Множественное сравнение средних значений было выполнено с использованием однофакторного дисперсионного анализа с posthoc-анализом Тьюки, значение p <0. 05 статистически значимо (**** <0,001, *** <0,001; ** <0,01; * <0,05) загружено в multicomp RStudio.
05 статистически значимо (**** <0,001, *** <0,001; ** <0,01; * <0,05) загружено в multicomp RStudio.
акозлова (Анна Козлова) · GitHub
акозлова (Анна Козлова) · GitHubОсновные моменты
- Участник Arctic Code Vault
Популярные репозитории
Форк от gluonhq / scenebuilder
Scene Builder — это визуальный инструмент компоновки с возможностью перетаскивания для разработки пользовательских интерфейсов приложений JavaFX.
Ява 2
Разветвляется от yopox / test_problem
Инспекция IntelliJ отлавливает веселье класса # в коде Java и Kotlin
Котлин
1001 публикация в прошлом году
ФевМарАпрМайИюньИюлАвгСентОктНоябДекЯнвснПн ВтСрЧтПтСбВклад деятельности
Февраль 2021 г.
 Создано 38
совершает в
2
репозитории
Отзыв 1
запрос на включение в
1
хранилище
JetBrains / intellij-покрытие
1 запрос на включение
Вы не можете выполнить это действие в настоящее время.Вы вошли в систему с другой вкладкой или окном. Перезагрузите, чтобы обновить сеанс.
Вы вышли из системы на другой вкладке или в другом окне. Перезагрузите, чтобы обновить сеанс.
Создано 38
совершает в
2
репозитории
Отзыв 1
запрос на включение в
1
хранилище
JetBrains / intellij-покрытие
1 запрос на включение
Вы не можете выполнить это действие в настоящее время.Вы вошли в систему с другой вкладкой или окном. Перезагрузите, чтобы обновить сеанс.
Вы вышли из системы на другой вкладке или в другом окне. Перезагрузите, чтобы обновить сеанс.Новая стратегия объединенного анализа молока для диагностики Dictyocaulus viviparus на уровне стада
Основные моменты
- •
Нетели в первый период лактации имеют более высокие уровни антител к молоку, чем у старых животных.
- •
Оптимальный тест производится при объединении 10 образцов телок первого периода лактации.
- •
Объединенный тест превосходит общий тест при диагностике легочного червя в стаде.
Реферат
Современные методы диагностики для обнаружения присутствия или отсутствия Dictyocaulus viviparus в молочных стадах нечувствительны, если они основаны на определении уровней антител в молоке бестарной емкости (BTM). Здесь мы представляем новый метод подтверждения присутствия паразита, основанный на образце объединенного молока от 10 случайно выбранных первотелок (FLH).Это исследование состояло из двух частей. Во-первых, было проведено продольное исследование для изучения динамики заражения в образцах молока в течение пастбищного сезона с использованием прототипа ELISA, разработанного Svanova (Boehringer-Ingelheim, Uppsala). Мы определили, что средние значения ODR в образцах молока из FLH были значительно выше, чем у более старых коров (0,13 против 0,07 соответственно, p <0,001), что позволяет предположить, что образцы из когорты FLH должны быть объединены для проведения теста. Во-вторых, объединенный тест молока оценивался на поперечном обследовании молочных стад в Великобритании (n = 25 стад и n = 25 стад без выпаса) для оценки эффективности теста в полевых условиях.Пороговое значение коэффициента оптической плотности (ODR) для нашего теста на объединенное молоко с использованием 10 образцов молока FLH было оптимальным при значении 0,16. Объединение 10 образцов FLH создало чувствительность и специфичность 66,7% и 95,5% соответственно. Для сравнения, образцы BTM всего стада имели максимальную чувствительность 37,5% и специфичность 63,6% при пороговом значении ODR 0,18. Площадь под кривой согласно анализу эксплуатационных характеристик получателя (ROC) была высокой для теста из 10 телок (0,87), но плохой для теста BTM всего стада (0.45). Это исследование обеспечивает более чувствительную стратегию диагностических тестов для скрининга D. viviparus в молочных стадах. Тестирование стада в конце пастбищного сезона облегчило бы планирование эффективных мер борьбы, таких как вакцинация от легочных червей или стратегическая дегельминтизация, на следующий пастбищный сезон. Это может оказаться полезной стратегией тестирования для диагностики различных паразитарных болезней домашнего скота.
Во-вторых, объединенный тест молока оценивался на поперечном обследовании молочных стад в Великобритании (n = 25 стад и n = 25 стад без выпаса) для оценки эффективности теста в полевых условиях.Пороговое значение коэффициента оптической плотности (ODR) для нашего теста на объединенное молоко с использованием 10 образцов молока FLH было оптимальным при значении 0,16. Объединение 10 образцов FLH создало чувствительность и специфичность 66,7% и 95,5% соответственно. Для сравнения, образцы BTM всего стада имели максимальную чувствительность 37,5% и специфичность 63,6% при пороговом значении ODR 0,18. Площадь под кривой согласно анализу эксплуатационных характеристик получателя (ROC) была высокой для теста из 10 телок (0,87), но плохой для теста BTM всего стада (0.45). Это исследование обеспечивает более чувствительную стратегию диагностических тестов для скрининга D. viviparus в молочных стадах. Тестирование стада в конце пастбищного сезона облегчило бы планирование эффективных мер борьбы, таких как вакцинация от легочных червей или стратегическая дегельминтизация, на следующий пастбищный сезон. Это может оказаться полезной стратегией тестирования для диагностики различных паразитарных болезней домашнего скота.
Ключевые слова
Dictyocaulus viviparus
Диктиокаулез
Диагностика
ELISA
Молочная корова
Молоко
ROC-анализ
0 (Рекомендуемые статьи)
0 Цитирующие статьиОпубликовано Elsevier B.V.
Рекомендуемые статьи
Цитирование статей
| |
Эта статья цитируется в научной статье 1 (всего в статье 1 ) Численный алгоритм самосогласованного стационарного уровня для многомерных нестационарных временных рядов Кислицын А.А. , Козлова А.Б. , Машеров Е.Л. , Ю.Орлов Н.А. Аннотация: В данной статье мы рассматриваем самосогласованный стационарный уровень временных рядов электроэнцефалограммы. Практическая цель этой статистики — построить индикатор беспорядка. В отличие от классической задачи стационарного тестирования двух выборок, в нашем случае необходимо построить индикатор для предсказания смены нестационарного режима. Например, мы рассматриваем специальный предиктор приступа эпилепсии. Ключевые слова: нестационарный индекс, индикатор расстройства, электроэнцефалограмма, приступ эпилепсии. DOI: https://doi.org/10.20948/prepr-2017-124-e Полный текст: PDF-файл (1124 kB) Язык: Образец цитирования: А.А. Кислицын, А.Б. Козлова, Е.Л. Машеров, Ю. Н. Орлов, “Численный алгоритм самосогласованного стационарного уровня для многомерных нестационарных временных рядов”, Препринты ИПМ им. М.В. Келдыша, 2017, 124, 14 с. Цитирование в формате AMSBIB Варианты соединения: Цитирующие статьи в Google Scholar: Русские цитаты,
Цитаты на английском языке Эта публикация цитируется в следующих статьях:
|
| |
 Келдыша, 2017, 124, 14 с. (Mi ipmp2340)
Келдыша, 2017, 124, 14 с. (Mi ipmp2340)
 org / 10.20948 / prepr-2017-124-e}
org / 10.20948 / prepr-2017-124-e} Совместное культивирование стволовых клеток нервного гребня (NCSC) и клеток, продуцирующих бета-TC6 инсулина, приводит к соединениям кадгерина и защите от индуцированной цитокинами гибели бета-клеток
Аннотация
Назначение
Трансплантации островков поджелудочной железы пациентам с диабетом 1 типа препятствуют воспалительные реакции в месте трансплантации, ведущие к дисфункции и гибели бета-клеток, продуцирующих инсулин. Недавно мы показали, что совместная трансплантация стволовых клеток нервного гребня (NCSC) вместе с островковыми клетками улучшает результат трансплантации. Целью настоящего исследования было описание взаимодействий in vitro, между NCSC и инсулин-продуцирующими бета-TC6 клетками, которые могут опосредовать защиту от индуцированной цитокинами гибели бета-клеток.
Недавно мы показали, что совместная трансплантация стволовых клеток нервного гребня (NCSC) вместе с островковыми клетками улучшает результат трансплантации. Целью настоящего исследования было описание взаимодействий in vitro, между NCSC и инсулин-продуцирующими бета-TC6 клетками, которые могут опосредовать защиту от индуцированной цитокинами гибели бета-клеток.
Процедуры
КлеткиBeta-TC6 и NCSC культивировали либо по отдельности, либо вместе, либо со вставками для культур клеток, либо без них.Затем культуры подвергали действию провоспалительных цитокинов IL-1β и IFN-γ в течение 48 часов с последующим анализом скорости клеточной гибели (проточная цитометрия), продукции нитритов (реагент Грисса), локализации белка (иммунофлуоресценция) и фосфорилирования белка (проточная цитометрия). цитометрия).
Результаты
Мы наблюдали, что клетки бета-TC6, совместно культивируемые с NCSC, были защищены от цитокин-индуцированной гибели клеток, но не при разделении вставками из клеточных культур. Это происходило параллельно с (i) повышенным продуцированием нитрита из клеток бета-ТС6, что указывает на то, что повышенная выживаемость клеток позволяет устойчиво производить оксид азота; (ii) продуцирование ламинина на основе NCSC; (iii) снижение окрашивания фосфо-FAK в очаговых адгезиях клеток бета-TC6 и (iv) снижение клетками бета-TC6 фосфорилирования ERK (T202 / Y204), FAK (Y397) и FAK (Y576).Более того, совместное культивирование также приводило к накоплению кадгерина и бета-катенина на стыках клеток NCSC / бета-TC6. Наконец, ингибитор щелевых соединений карбеноксолон не влиял на вызванную цитокинами гибель бета-клеток во время совместного культивирования с NCSC.
Это происходило параллельно с (i) повышенным продуцированием нитрита из клеток бета-ТС6, что указывает на то, что повышенная выживаемость клеток позволяет устойчиво производить оксид азота; (ii) продуцирование ламинина на основе NCSC; (iii) снижение окрашивания фосфо-FAK в очаговых адгезиях клеток бета-TC6 и (iv) снижение клетками бета-TC6 фосфорилирования ERK (T202 / Y204), FAK (Y397) и FAK (Y576).Более того, совместное культивирование также приводило к накоплению кадгерина и бета-катенина на стыках клеток NCSC / бета-TC6. Наконец, ингибитор щелевых соединений карбеноксолон не влиял на вызванную цитокинами гибель бета-клеток во время совместного культивирования с NCSC.
Заключение
Таким образом, прямые контакты, но не растворимые факторы, способствуют повышению жизнеспособности бета-TC6 при совместном культивировании с NCSC. Мы предполагаем, что кадгериновые соединения между NCSC и бета-TC6 клетками обеспечивают мощные сигналы, которые поддерживают выживание бета-клеток, даже несмотря на то, что передача сигналов ERK и FAK подавлена. Возможно, что будущие стратегии по улучшению результатов трансплантации островков могут выиграть от попыток увеличить количество соединений кадгерина бета-клеток с соседними клетками.
Возможно, что будущие стратегии по улучшению результатов трансплантации островков могут выиграть от попыток увеличить количество соединений кадгерина бета-клеток с соседними клетками.
Цитирование: Ngamjariyawat A, Turpaev K, Vasylovska S, Kozlova EN, Welsh N (2013) Совместное культивирование стволовых клеток нервного гребня (NCSC) и инсулиновых клеток, продуцирующих бета-TC6, приводит к соединениям кадгерина и защите от индуцированных цитокинами Бета-клеточная смерть. PLoS ONE 8 (4): e61828. https: // doi.org / 10.1371 / journal.pone.0061828
Редактор: Либинг Сун, Онкологический центр Университета Сунь Ятсена, Китай
Поступила: 18 октября 2012 г .; Одобрена: 13 марта 2013 г .; Опубликовано: 17 апреля 2013 г.
Авторские права: © 2013 Ngamjariyawat et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа была частично поддержана Шведским исследовательским советом (2010-11564-15-3, (NW) и K2011-62X-20716-04-6 (EK)), Шведской диабетической ассоциацией ( NW), семья Ernfors Foundation (NW), Barndiabetesfonden (NW), Novo-Nordisk Foundation (NW), Stiftelsen Olle Engkvist Byggare (EK), Signhild Engkvist Stiftelse (EK) и программа Шведского института Visby 00613/2011 (EK ). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Диабет 1 типа — это аутоиммунное заболевание, которое приводит к разрушению бета-клеток, продуцирующих инсулин. Цитокины, такие как IL-1β, TNF-α и IFN-γ, вызывают гибель бета-клеток in vitro ; и предполагается, что местное высвобождение тех же цитокинов участвует в разрушении бета-клеток поджелудочной железы in vivo [1]. Действительно, уровни провоспалительных цитокинов коррелировали с инсулитом и деструкцией бета-клеток как у мышей NOD [2], так и в биоптатах поджелудочной железы человека от пациентов с недавно начавшимся диабетом 1 типа [3].После активации рецептора передача сигналов, индуцированная цитокинами, включает активацию митоген-активируемых протеинкиназ (MAPK), c-Jun Nh3-терминальной киназы (JNK), киназы, регулируемой внеклеточным сигналом (ERK), и p38 MAPK [4], [5 ]. Помимо митоген-активируемых протеинкиназ, индуцированная IL-1β и TNF-α передача сигналов приводит к активации провоспалительного фактора транскрипции NF-κB [6]. Было высказано предположение, что на островках грызунов индуцированная цитокинами гибель клеток вызывается повышенным образованием оксида азота, которое является результатом активации транскрипции гена индуцибельной синтазы оксида азота, опосредованной NF-κB [6], [7].
Действительно, уровни провоспалительных цитокинов коррелировали с инсулитом и деструкцией бета-клеток как у мышей NOD [2], так и в биоптатах поджелудочной железы человека от пациентов с недавно начавшимся диабетом 1 типа [3].После активации рецептора передача сигналов, индуцированная цитокинами, включает активацию митоген-активируемых протеинкиназ (MAPK), c-Jun Nh3-терминальной киназы (JNK), киназы, регулируемой внеклеточным сигналом (ERK), и p38 MAPK [4], [5 ]. Помимо митоген-активируемых протеинкиназ, индуцированная IL-1β и TNF-α передача сигналов приводит к активации провоспалительного фактора транскрипции NF-κB [6]. Было высказано предположение, что на островках грызунов индуцированная цитокинами гибель клеток вызывается повышенным образованием оксида азота, которое является результатом активации транскрипции гена индуцибельной синтазы оксида азота, опосредованной NF-κB [6], [7].
Трансплантация изолированных островков стала вариантом лечения отдельных пациентов с диабетом 1 типа. Однако адекватная функция трансплантата наблюдается менее чем у 10% пациентов через 5 лет [8]. Немедленная посттрансплантационная гибель клеток и отказ трансплантата, вероятно, связаны с гипоксией в сочетании с воспалительными явлениями [9]. Таким образом, увеличение массы бета-клеток и ослабление воспаления после трансплантации являются важными целями для успешной трансплантации островков. Возможно, что совместная трансплантация островковых клеток со стволовыми клетками нервного гребня (NCSC) будет способствовать улучшенному результату трансплантации островков.Действительно, недавние исследования показали, что NCSCs играют важную роль в дифференцировке бета-клеток, регулируя массу бета-клеток во время развития [10]. Также было замечено, что ганглии зародышевых дорзальных корешков влияют на секрецию инсулина в совместно культивируемых островках [11], что совместно культивируемые островки и NCSCs оказывают взаимовыгодное действие in vitro [12], и что совместная трансплантация NCSC-производных нейросферы с островками индуцируют пролиферацию и стимулируют функцию трансплантированных бета-клеток [13].
Однако адекватная функция трансплантата наблюдается менее чем у 10% пациентов через 5 лет [8]. Немедленная посттрансплантационная гибель клеток и отказ трансплантата, вероятно, связаны с гипоксией в сочетании с воспалительными явлениями [9]. Таким образом, увеличение массы бета-клеток и ослабление воспаления после трансплантации являются важными целями для успешной трансплантации островков. Возможно, что совместная трансплантация островковых клеток со стволовыми клетками нервного гребня (NCSC) будет способствовать улучшенному результату трансплантации островков.Действительно, недавние исследования показали, что NCSCs играют важную роль в дифференцировке бета-клеток, регулируя массу бета-клеток во время развития [10]. Также было замечено, что ганглии зародышевых дорзальных корешков влияют на секрецию инсулина в совместно культивируемых островках [11], что совместно культивируемые островки и NCSCs оказывают взаимовыгодное действие in vitro [12], и что совместная трансплантация NCSC-производных нейросферы с островками индуцируют пролиферацию и стимулируют функцию трансплантированных бета-клеток [13]. Совсем недавно совместное культивирование инсулин-продуцирующих клеток и NCSC привело как к защите от цитокин-индуцированной гибели бета-клеток [14], так и к увеличению роста мышиной массы бета-клеток [15]. Однако механизмы, с помощью которых NCSCs опосредуют положительный эффект на бета-клетки, еще не выяснены. Таким образом, цель настоящего исследования состояла в более подробном изучении взаимодействий между NCSC и инсулино-продуцирующими клетками in vitro и в сопоставлении таких взаимодействий с улучшением выживаемости в ответ на воздействие цитокинов.Понимание таких событий имеет решающее значение для разработки новых протоколов для улучшения результатов трансплантации островков у пациентов с диабетом.
Совсем недавно совместное культивирование инсулин-продуцирующих клеток и NCSC привело как к защите от цитокин-индуцированной гибели бета-клеток [14], так и к увеличению роста мышиной массы бета-клеток [15]. Однако механизмы, с помощью которых NCSCs опосредуют положительный эффект на бета-клетки, еще не выяснены. Таким образом, цель настоящего исследования состояла в более подробном изучении взаимодействий между NCSC и инсулино-продуцирующими клетками in vitro и в сопоставлении таких взаимодействий с улучшением выживаемости в ответ на воздействие цитокинов.Понимание таких событий имеет решающее значение для разработки новых протоколов для улучшения результатов трансплантации островков у пациентов с диабетом.
Материалы и методы
Мыши
Трансгенных гетерозиготных мышей с зеленым флуоресцентным белком (EGFP), усиленного C57BL / 6-β-актином (Jackson Laboratories, Bar Harbor, ME, http://www.jax.org), использовали для создания NCSC.
Заявление об этике
Все процедуры были одобрены Региональным комитетом по этике исследований на животных (Региональный комитет по этике исследований на животных округа Уппсала).
Ячейки Beta-TC6
Клетки бета-TC6 мыши, приобретенные в ATCC, поддерживали в RPMI-1640 (Gibco, Гранд-Айленд, Нью-Йорк, США) с добавлением 10% фетальной телячьей сыворотки (Sigma Chemicals St Louis, Миссури, США), 2 мМ L-глутамина. , стрептомицин (0,1 мг / мл) и бензилпенициллин (100 Ед / мл). Все клетки выращивали при 37 ° C в инкубаторе с увлажненным воздухом с 5% CO 2 .
Подготовка культуры NCSC
Ганглии задних корешков из эмбрионов мышей EGFP в возрасте E11,5 дня были изолированы и использованы для создания нейросфер NCSC (линия клеток NL38) из так называемой пограничной крышки [16], [17].Вкратце, мышей анестезировали путем внутрибрюшинной инъекции ксилазина (RompunVR vet .; http://www.bayer.com) и кетамина (KetaminolVR vet .; http://www.intervet.com) (10 и 100 нг на грамм тела). вес, соответственно), и матка была удалена из анестезированной беременной мыши и помещена в холодный PBS. Эмбрионы были отделены и промыты в PBS, помещены в среду DMEM / F12 (Invitrogen, Carlsbad, CA, USA), дополненную N-2 (Invitrogen, Carlsbad, CA, USA) (среда N-2), после чего спинной корешок ганглии были удалены и собраны. Собранным ганглиям задних корешков давали осесть перед удалением фракций супернатанта и добавлением коллагеназы / диспазы (1 мг / мл) (Roche Diagnostics Scandinavia, Bromma, Швеция) и раствора ДНКазы (0,5 мг / мл) (Sigma-Aldrich) в N -2 и инкубировать от 20 до 30 мин на водяной бане при 37 ° C. После этого проводили промывание в среде N-2 с добавлением B-27 (1-50) (Invitrogen, Карлсбад, Калифорния, США), титрование и высев 1-2 × 10 5 клеток / лунку в 24-луночный планшет. после разобщения.Клетки помещали непосредственно в 500 мкл среды N-2, содержащей B-27, эпидермальный фактор роста (20 нг / мл) (R&D Systems, Миннеаполис, Миннесота, США) и основной фактор роста фибробластов (20 нг / мл) (R&D Systems). Через 12 часов неприлипающие клетки удаляли вместе с половиной среды перед добавлением 250 мкл свежей среды. Затем среду меняли через день (50% среды заменяли свежей средой) до начала формирования нейросфер. Нейросферы из пассажей с 4 по 5 трипсинизировали в течение 5 минут для получения свободных суспензий отдельных клеток для последующего совместного культивирования с клетками бета-TC6.
Собранным ганглиям задних корешков давали осесть перед удалением фракций супернатанта и добавлением коллагеназы / диспазы (1 мг / мл) (Roche Diagnostics Scandinavia, Bromma, Швеция) и раствора ДНКазы (0,5 мг / мл) (Sigma-Aldrich) в N -2 и инкубировать от 20 до 30 мин на водяной бане при 37 ° C. После этого проводили промывание в среде N-2 с добавлением B-27 (1-50) (Invitrogen, Карлсбад, Калифорния, США), титрование и высев 1-2 × 10 5 клеток / лунку в 24-луночный планшет. после разобщения.Клетки помещали непосредственно в 500 мкл среды N-2, содержащей B-27, эпидермальный фактор роста (20 нг / мл) (R&D Systems, Миннеаполис, Миннесота, США) и основной фактор роста фибробластов (20 нг / мл) (R&D Systems). Через 12 часов неприлипающие клетки удаляли вместе с половиной среды перед добавлением 250 мкл свежей среды. Затем среду меняли через день (50% среды заменяли свежей средой) до начала формирования нейросфер. Нейросферы из пассажей с 4 по 5 трипсинизировали в течение 5 минут для получения свободных суспензий отдельных клеток для последующего совместного культивирования с клетками бета-TC6.
10 5 диспергированных NCSC помещали в 24-луночные планшеты или вставки из ПЭТ-трековых мембран с размером пор 0,4 мм (Falcon) и позволяли им покрыть большую часть поверхности в течение трех дней культивирования в данной культуральной среде N-2. над. Все лунки / вставки были предварительно покрыты ламинином (10 мкг / мл), чтобы способствовать эффективному распространению NCSC. Через три дня 10 4 бета-TC6 клеток высевали отдельно или вместе с клетками NCSC.На этом этапе культуральную среду заменили на среду RPMI-1640, содержащую такие же добавки, как указано выше. Для совместного культивирования со вставками клетки бета-TC6 высевали так, чтобы клетки располагались на дне лунки, а клетки NCSC находились выше во вставках. После двух дней совместного культивирования клетки либо оставляли необработанными, либо обрабатывали смесью цитокинов (20 нг / мл IL-1β + 20 нг / мл IFN-γ; Peprotech) в течение дополнительных 48 часов. После периода воздействия цитокинов образцы культуральной среды анализировали на содержание нитритов с помощью реактива Грисса [4].
После периода воздействия цитокинов образцы культуральной среды анализировали на содержание нитритов с помощью реактива Грисса [4].
Проточно-цитометрический анализ жизнеспособности клеток
In vitro культур клеток бета-TC6, NCSC или бета-TC6 + NCSC метили в течение 10 мин при 37 ° C с помощью 10 мкг / мл йодида пропидия (Sigma-Aldrich). В некоторых экспериментах клетки обрабатывали ингибитором щелевых контактов карбеноксолоном (50 мкМ; Sigma Aldrich) в течение 48-часового периода воздействия цитокинов. Клетки один раз промывали PBS, а затем трипсинизировали в течение 5 минут при 37 ° C. Суспензии клеток анализировали в проточном цитометре Becton Dickinson FACSCalibur на флуоресценцию FL1 (GFP) и FL3 (иодид пропидия).Частоты гибели клеток определяли количественно для GFP-положительных и GFP-отрицательных клеток отдельно и выражали как процент от общего количества GFP-положительных и отрицательных клеток, соответственно.
Иммуноокрашивание
Клетки фиксировали в 4% забуференном параформальдегиде при комнатной температуре в течение 5 минут, затем промывали PBS перед проницаемостью и блокированием с использованием PBS с 0,1% тритоном® X-100 (Sigma), 1% BSA (Sigma) и 3% эмбрионального теленка.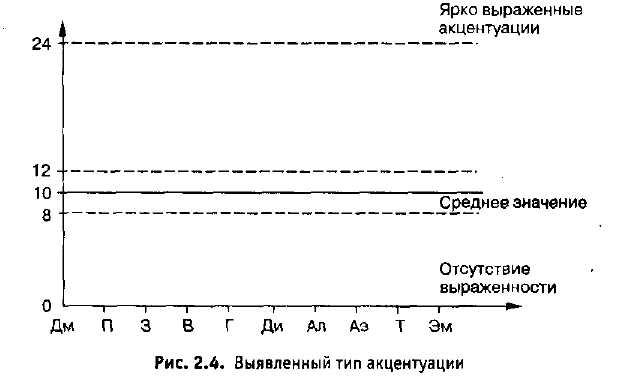 сыворотка. Клетки инкубировали с первичными антителами в PBS с 1% BSA и 1% эмбриональной телячьей сывороткой в течение 30 минут при 37 ° C перед двукратной промывкой PBS.Затем культуры инкубировали со вторичными антителами в течение 30 минут при 37 ° C и трижды промывали PBS в течение 15 минут, вторая промывка включала Hoechst 33242 (11 нг / мл, Invitrogen). Покровные стекла помещали на предметные стекла с флуоресцентным монтажным раствором Dako Cytomation. Первичные антитела были следующими: анти-NOS2 (моноклональная мышь, 1-100, Санта-Крус), анти-бета-катенин (поликлональный кролик, 1-100, Abcam), анти-пан кадгерин (моноклональная мышь, 1-100, Abcam). , PE-конъюгированный альфа-6-интегрин (1100, Abcam), анти-ламинин (поликлональный кролик, 1∶200, Sigma) и фосфо-FAK (pY397) (поликлональный кролик, 1∶100, Invitrogen).Вторичными антителами были Cy3 (ослиные антимышиные, 1-500, лаборатории Джексона) и Alexa мука 555 (козьи антикроличьи, 1-600, Invitrogen).
сыворотка. Клетки инкубировали с первичными антителами в PBS с 1% BSA и 1% эмбриональной телячьей сывороткой в течение 30 минут при 37 ° C перед двукратной промывкой PBS.Затем культуры инкубировали со вторичными антителами в течение 30 минут при 37 ° C и трижды промывали PBS в течение 15 минут, вторая промывка включала Hoechst 33242 (11 нг / мл, Invitrogen). Покровные стекла помещали на предметные стекла с флуоресцентным монтажным раствором Dako Cytomation. Первичные антитела были следующими: анти-NOS2 (моноклональная мышь, 1-100, Санта-Крус), анти-бета-катенин (поликлональный кролик, 1-100, Abcam), анти-пан кадгерин (моноклональная мышь, 1-100, Abcam). , PE-конъюгированный альфа-6-интегрин (1100, Abcam), анти-ламинин (поликлональный кролик, 1∶200, Sigma) и фосфо-FAK (pY397) (поликлональный кролик, 1∶100, Invitrogen).Вторичными антителами были Cy3 (ослиные антимышиные, 1-500, лаборатории Джексона) и Alexa мука 555 (козьи антикроличьи, 1-600, Invitrogen).
Иммуноблоттинг
Клетки лизировали в буфере для образцов SDS, кипятили в течение 5 мин и разделяли с помощью SDS-PAGE. Белки электрофоретически переносили на мембрану Hybond-P (GE Healthcare, Упсала, Швеция). Мембраны инкубировали со следующими первичными антителами: мышиным анти-NOS2 (C-11, Santa Cruz) и мышиным антителом к тубулину (Santa Cruz).Иммунодетекция выполнялась, как описано для системы определения иммуноблоттинга ECL (GE healthcare) и с использованием станции Kodak Image 4000 MM.
Белки электрофоретически переносили на мембрану Hybond-P (GE Healthcare, Упсала, Швеция). Мембраны инкубировали со следующими первичными антителами: мышиным анти-NOS2 (C-11, Santa Cruz) и мышиным антителом к тубулину (Santa Cruz).Иммунодетекция выполнялась, как описано для системы определения иммуноблоттинга ECL (GE healthcare) и с использованием станции Kodak Image 4000 MM.
Микроскопический анализ
Иммуномеченных слайдов анализировали на флуоресцентном микроскопе Nikon Eclipse E800.
Проточно-цитометрический анализ фосфорилирования ERK, Akt и FAK
Использовали следующие первичные антитела: PerCP-конъюгированный Phospho-ERK1 / 2 (T202 / Y204) (Becton Dickinson), PE-конъюгированный Phospho-Akt (T473) (Becton Dickinson), кроличий Phospho-FAK (Y397) (Invitrogen) и кроличий фосфо-FAK (Y576 / 577) (Cell Signaling).Вторичным антителом к фосфо-FAK-антителам был DyLight 650 (козий антикроличий, Abcam). Вкратце, клетки быстро фиксировали в соответствии с протоколом набора BD Phosflow (Becton-Dickinson). Затем клетки пермабилизировали, инкубировали с антителами и промывали для анализа флуоресцентных сигналов с помощью проточной цитометрии (FACSCalibur).
Затем клетки пермабилизировали, инкубировали с антителами и промывали для анализа флуоресцентных сигналов с помощью проточной цитометрии (FACSCalibur).
Статистический анализ
Данные были проанализированы с использованием парного t-критерия Стьюдента. Достоверность принималась при p <0,05.
Результаты
Влияние совместного культивирования NCSC-beta-TC6 на вызванную цитокинами гибель клеток, продукцию нитритов и экспрессию iNOS
В предыдущем исследовании мы наблюдали, что NCSC частично защищают бета-клетки от гибели, вызванной цитокинами, при совместном культивировании [14].Однако мы не смогли определить, была ли защита получена за счет высвобождения растворимых факторов или требовался прямой межклеточный контакт. Таким образом, в настоящее время мы культивировали NCSC совместно с продуцирующими инсулин мышиными бета-TC6 клетками со вставками для культивирования клеток размером 0,4 микрометра или без них, которые обеспечивают прохождение растворимых факторов, но не прямой контакт между клетками. Мы наблюдали, что индуцированной цитокинами гибели клеток бета-TC6 противодействовали совместное культивирование с NCSC (рис. 1). Когда клетки были разделены вставками, позволяющими проникать растворимым факторам, не было обнаружено защиты от цитокинов (рис. 1).Это предполагает, что для защиты необходим прямой контакт между клетками бета-TC6 и NCSC. NCSC не реагировали на воздействие цитокинов ни при культивировании отдельно, ни в сочетании с клетками бета-TC6 (рис. 1).
Мы наблюдали, что индуцированной цитокинами гибели клеток бета-TC6 противодействовали совместное культивирование с NCSC (рис. 1). Когда клетки были разделены вставками, позволяющими проникать растворимым факторам, не было обнаружено защиты от цитокинов (рис. 1).Это предполагает, что для защиты необходим прямой контакт между клетками бета-TC6 и NCSC. NCSC не реагировали на воздействие цитокинов ни при культивировании отдельно, ни в сочетании с клетками бета-TC6 (рис. 1).
Рисунок 1. Совместное культивирование клеток бета-ТС6 с NCSC в прямом контакте приводит к защите от индуцированной цитокинами гибели бета-клеток.
NCSC помещали на планшеты или вставки, покрытые ламинином, и через три дня добавляли клетки бета-TC6. Еще через 48 часов добавляли IL-1β (20 нг / мл) и IFN-β (20 нг / мл).Спустя 48 часов клетки метили йодидом пропидия, трипсинизировали и анализировали на гибель клеток проточной цитометрией. GFP-положительные клетки (NCSC) и GFP-отрицательные клетки (бета-TC6) регистрировали и анализировали отдельно. Результаты представляют собой средние значения ± SEM для 5 независимых экспериментов. ** обозначает p <0,01 с использованием парного t-критерия Стьюдента.
Результаты представляют собой средние значения ± SEM для 5 независимых экспериментов. ** обозначает p <0,01 с использованием парного t-критерия Стьюдента.
https://doi.org/10.1371/journal.pone.0061828.g001
Цитокины способствуют гибели бета-клеток грызунов in vitro в основном за счет активации фермента индуцибельной синтазы оксида азота (iNOS) [1].Чтобы выяснить, влияет ли совместное культивирование клеток бета-TC6 с NCSC на индукцию iNOS, мы затем проанализировали нитрит, который является конечным продуктом оксида азота, производного от iNOS. Уровни нитритов в клетках, не подвергшихся действию цитокинов IL-1β и IFN-γ, не были обнаружены (результаты не показаны). В клетках, подвергнутых воздействию цитокинов, продукция нитритов была примерно в 10 раз выше в клетках бета-ТС6, чем в NCSC при культивировании отдельно (рис. 2). Совместное культивирование клеток бета-TC6 и NCSC, разделенных вставкой, не влияло на продукцию нитрита (рис. 2).Однако совместное культивирование без вставок привело к усилению продукции нитрита по сравнению с одними клетками бета-ТС6 (рис. 2).
2).
Рисунок 2. Совместное культивирование клеток бета-TC6 с NCSC, находящимися в прямом контакте, приводит к увеличению продукции нитритов в ответ на воздействие цитокинов.
NCSC помещали на планшеты или вставки, покрытые ламинином, и через три дня добавляли клетки бета-TC6. После 48 часов совместного культивирования IL-1β (20 нг / мл) и IFN-γ (20 нг / мл) добавляли в течение еще 48 часов периода культивирования.В конце этого периода культивирования образцы питательной среды анализировали на нитриты с использованием реактива Грисса. Результаты представляют собой средние значения ± SEM для 4 независимых экспериментов. * обозначает p <0,05 с использованием парного t-критерия Стьюдента.
https://doi.org/10.1371/journal.pone.0061828.g002
Чтобы оценить происхождение цитокин-индуцированной продукции оксида азота, мы окрашивали клетки, культивированные без вставок, на иммунореактивность iNOS. Клетки бета-TC6 наблюдались как не-GFP-положительные клетки, расположенные в виде клеточных островков, окруженных GFP-положительным монослоем NCSC (рис. 3А).Окрашивание iNOS не наблюдалось в клетках, не обработанных цитокинами. Обработка цитокинов привела к явной потере клеток бета-TC6 при культивировании отдельно (рис. 3А). Среди оставшихся клеток бета-TC6 некоторые проявляли четкую иммунореактивность iNOS. Также при совместном культивировании с NCSC некоторые клетки бета-TC6 были iNOS-положительными (рис. 3A). Ни один из NCSC не показал иммунореактивности iNOS. Чтобы дополнительно подтвердить, что iNOS экспрессируется только в клетках бета-TC6, обработанных цитокинами, а не в NCSC, мы провели иммуноблот-анализ двух типов клеток.Мы наблюдали иммунореактивность iNOS в клетках бета-TC6, обработанных цитокинами, но не в NCSC (рис. 3В). Таким образом, оксид азота в основном продуцируется клетками бета-TC6, и более высокая выживаемость, индуцированная прямым совместным культивированием с NCSC, позволяет дополнительно увеличивать и / или пролонгировать продукцию оксида азота.
3А).Окрашивание iNOS не наблюдалось в клетках, не обработанных цитокинами. Обработка цитокинов привела к явной потере клеток бета-TC6 при культивировании отдельно (рис. 3А). Среди оставшихся клеток бета-TC6 некоторые проявляли четкую иммунореактивность iNOS. Также при совместном культивировании с NCSC некоторые клетки бета-TC6 были iNOS-положительными (рис. 3A). Ни один из NCSC не показал иммунореактивности iNOS. Чтобы дополнительно подтвердить, что iNOS экспрессируется только в клетках бета-TC6, обработанных цитокинами, а не в NCSC, мы провели иммуноблот-анализ двух типов клеток.Мы наблюдали иммунореактивность iNOS в клетках бета-TC6, обработанных цитокинами, но не в NCSC (рис. 3В). Таким образом, оксид азота в основном продуцируется клетками бета-TC6, и более высокая выживаемость, индуцированная прямым совместным культивированием с NCSC, позволяет дополнительно увеличивать и / или пролонгировать продукцию оксида азота.
Рисунок 3. Экспрессия iNOS может быть обнаружена в клетках бета-TC6.
(A) Не-GFP-положительные бета-TC6 клетки культивировали либо отдельно (левые панели), либо вместе с GFP-положительными клетками NCSC (правые панели, зеленые клетки) на покровных стеклах, покрытых ламинином.Клетки обрабатывали 20 нг / мл IL-1β и 20 нг / мл IFN-γ в течение 48 часов. Затем клетки фиксировали, пермеабилизировали и окрашивали на iNOS (красный). Ядра были визуализированы Hoechst. Затем флуоресценцию фотографировали во флуоресцентном микроскопе с объективом 40x. Белая шкала показывает расстояние 100 мкм. Результаты являются репрезентативными для трех независимых экспериментов. (B) Иммуноблот-анализ NCSC (левая панель) и бета-TC6 клеток (правая панель). Оба типа клеток культивировали по отдельности. Клетки стимулировали в течение 12 часов IL-1β и IFN-γ, а затем анализировали на экспрессию iNOS с помощью анализа иммуноблоттинга.В качестве контроля нагрузки использовали тубулин. Результаты являются репрезентативными для трех наблюдений.
https://doi. org/10.1371/journal.pone.0061828.g003
org/10.1371/journal.pone.0061828.g003
Влияние совместного культивирования бета-TC6 и NCSC клеток на иммунофлуоресценцию ламинина, альфа-интегрина и фосфо-FAK
Чтобы изучить механизмы, с помощью которых прямые контакты между NCSC и бета-TC6 клетками способствуют повышению выживаемости бета-клеток, мы затем провели иммуноокрашивание на путь клеточной адгезии ламинин-интегрин-киназа очаговой адгезии (FAK).Иммунореактивность ламинина не наблюдалась в клетках бета-TC6, культивируемых отдельно (фиг. 4). Окрашивание совместно культивированных клеток показало, что NCSC, но не клетки бета-TC6, продуцируют ламинин (рис. 4). Иммунореактивность NCSC по ламинину часто находится в тесном контакте с ламинин-отрицательными бета-клетками (рис. 4, кружки). Это повышает вероятность того, что клетки бета-TC6 через интегрины присоединяются к ламинину внеклеточного матрикса, продуцируемому NCSC. Поэтому мы проанализировали иммуноокрашивание альфа6-интегрина, рецептора внеклеточного матрикса, связывающего ламинин [18]. Клетки Beta-TC6, культивированные отдельно, демонстрировали слабый цитоплазматический сигнал альфа6-интегрина (фиг. 4). Однако при совместном культивировании с NCSC этот сигнал не увеличивался. Вместо этого NCSCs показали четкие сигналы альфа6-интегрина, которые часто были локализованы в областях плазматической мембраны NCSCs (Рисунок 4). Это указывает на то, что взаимодействия ламинина / альфа6-интегрина происходят преимущественно в NCSC, но также могут присутствовать в клетках бета-TC6. Чтобы проверить это дальше, мы затем изучили иммуноокрашивание фосфо-FAK (Y397). Активация рецептора интегрина влияет на функцию, выживаемость и пролиферацию бета-клеток [19] и часто стимулирует фосфорилирование и накопление FAK в очаговых участках адгезии.Клетки Beta-TC6, культивированные в отсутствие NCSC, содержали многочисленные очаговые адгезии, окрашенные фосфо-FAK, локализованные на плазматической мембране (рис. 4). Эти фосфо-FAK-позитивные сайты встречались реже в NCSC. Интересно, что клетки бета-TC6 при совместном культивировании с NCSC и окружении с ними обнаруживают явное снижение количества фокальных адгезий фосфо-FAK (рис.
Клетки Beta-TC6, культивированные отдельно, демонстрировали слабый цитоплазматический сигнал альфа6-интегрина (фиг. 4). Однако при совместном культивировании с NCSC этот сигнал не увеличивался. Вместо этого NCSCs показали четкие сигналы альфа6-интегрина, которые часто были локализованы в областях плазматической мембраны NCSCs (Рисунок 4). Это указывает на то, что взаимодействия ламинина / альфа6-интегрина происходят преимущественно в NCSC, но также могут присутствовать в клетках бета-TC6. Чтобы проверить это дальше, мы затем изучили иммуноокрашивание фосфо-FAK (Y397). Активация рецептора интегрина влияет на функцию, выживаемость и пролиферацию бета-клеток [19] и часто стимулирует фосфорилирование и накопление FAK в очаговых участках адгезии.Клетки Beta-TC6, культивированные в отсутствие NCSC, содержали многочисленные очаговые адгезии, окрашенные фосфо-FAK, локализованные на плазматической мембране (рис. 4). Эти фосфо-FAK-позитивные сайты встречались реже в NCSC. Интересно, что клетки бета-TC6 при совместном культивировании с NCSC и окружении с ними обнаруживают явное снижение количества фокальных адгезий фосфо-FAK (рис. 4). Таким образом, прямой контакт с NCSCs может эффективно препятствовать образованию фокальных адгезий бета-TC6 клеток, даже если ламинин продуцируется окружающими NCSC.
4). Таким образом, прямой контакт с NCSCs может эффективно препятствовать образованию фокальных адгезий бета-TC6 клеток, даже если ламинин продуцируется окружающими NCSC.
Рисунок 4. Совместное культивирование клеток бета-ТС6 с NCSC влияет на очаговые адгезии бета-ТС6 клеток.
Не-GFP-положительные клетки бета-TC6 культивировали либо отдельно (левые панели), либо вместе с GFP-положительными клетками NCSC (правые панели, зеленые клетки) на покровных стеклах, покрытых ламинином. Клетки обрабатывали 20 нг / мл IL-1β и 20 нг / мл IFN-γ в течение 48 часов, а затем фиксировали, пермеабилизировали и окрашивали на ламинин (оранжевый), интегрин альфа VI (оранжевый) или фосфо-FAK (Y397). ) (красный). Ядра были визуализированы Hoechst.Флуоресценцию фотографировали на флуоресцентном микроскопе с линзой 20x. Круглые кружки указывают на положения скоплений ламинина NCSC вблизи бета-TC6 клеток. Масштабная линейка показывает расстояние 50 мкм. Результаты являются репрезентативными для трех независимых экспериментов.
https://doi.org/10.1371/journal.pone.0061828.g004
Воздействие цитокинов не повлияло на характеристики иммуноокрашивания ламинина или альфа-интегрина в клетках бета-TC6 ни при культивировании отдельно, ни при совместном культивировании с NCSC (результаты не показано).Однако сигнал фосфо-FAK (Y397) оказался более сильным в клетках бета-TC6, подвергшихся воздействию цитокинов, по сравнению с клетками бета-TC6, не подвергавшимися воздействию цитокинов (результаты не показаны).
Влияние совместного культивирования бета-TC6 и NCSC клеток на фосфорилирование ERK, Akt и FAK
Чтобы еще больше подтвердить наш вывод о том, что прямые контакты с NCSC подавляют активацию FAK бета-TC6 клеток, мы затем проанализировали влияние совместного культивирования на ERK (T202 / Y204), Akt (S473), FAK (Y397) и совместное культивирование клеток TC6. Фосфорилирование FAK (Y576 / 577) проточной цитометрией и методом Becton-Dickinson Phosflow.Этот подход позволяет количественно определять уровни фосфобелка в конкретной популяции клеток, а не во всех клетках совместной культуры. Внутриклеточное фосфорилирование ERK, Akt и FAK изучали в бета-TC6 и NCSC с помощью FL1-стробирования не-GFP и GFP-положительных клеток по отдельности, а затем количественного определения FL2 (PE-конъюгированных фосфо-Akt-антител) и FL3 (PerCP-конъюгированных phospho-ERK) сигналов в двух популяциях клеток. Мы наблюдали значительное снижение уровней фосфо-ERK, фосфо-FAK (Y397) и фосфо-FAK (Y576 / 577) в клетках бета-ТС6, совместно культивируемых с NCSC, по сравнению с клетками бета-ТС6, культивируемыми отдельно (рис. 5).Также была тенденция к снижению фосфо-Akt, но это не достигло статистической значимости. Тем не менее, настоящие результаты проточной цитометрии хорошо соответствуют результатам иммуноокрашивания (рис. 4), поскольку оба метода сообщают о пониженной активации FAK.
Внутриклеточное фосфорилирование ERK, Akt и FAK изучали в бета-TC6 и NCSC с помощью FL1-стробирования не-GFP и GFP-положительных клеток по отдельности, а затем количественного определения FL2 (PE-конъюгированных фосфо-Akt-антител) и FL3 (PerCP-конъюгированных phospho-ERK) сигналов в двух популяциях клеток. Мы наблюдали значительное снижение уровней фосфо-ERK, фосфо-FAK (Y397) и фосфо-FAK (Y576 / 577) в клетках бета-ТС6, совместно культивируемых с NCSC, по сравнению с клетками бета-ТС6, культивируемыми отдельно (рис. 5).Также была тенденция к снижению фосфо-Akt, но это не достигло статистической значимости. Тем не менее, настоящие результаты проточной цитометрии хорошо соответствуют результатам иммуноокрашивания (рис. 4), поскольку оба метода сообщают о пониженной активации FAK.
Рисунок 5. Влияние совместного культивирования на фосфорилирование ERK, AKT и FAK бета-TC6 клеток.
NCSC помещали на планшеты или вставки, покрытые ламинином, и через три дня добавляли клетки бета-TC6. После 48 часов совместного культивирования IL-1β и IFN-γ были добавлены в течение еще 48 часов периода культивирования.Затем клетки анализировали на фосфорилирование ERK1 / 2 (T202 / Y204), Akt (T473), FAK (Y397) и FAK (Y576 / 577) с использованием метода Becton Dickinson Phosflow. Результаты представляют собой средние значения ± SEM для 3 независимых наблюдений. * обозначает p <0,05 с использованием парного t-критерия Стьюдента.
После 48 часов совместного культивирования IL-1β и IFN-γ были добавлены в течение еще 48 часов периода культивирования.Затем клетки анализировали на фосфорилирование ERK1 / 2 (T202 / Y204), Akt (T473), FAK (Y397) и FAK (Y576 / 577) с использованием метода Becton Dickinson Phosflow. Результаты представляют собой средние значения ± SEM для 3 независимых наблюдений. * обозначает p <0,05 с использованием парного t-критерия Стьюдента.
https://doi.org/10.1371/journal.pone.0061828.g005
Воздействие цитокинов в течение 48 часов не оказало значительного влияния на фосфорилирование ERK и Akt, но фосфорилирование FAK (Y576) стимулировалось цитокинами бета-TC6. клетки (рисунок 5).На уровни фосфорилирования не повлияли NCSC, культивированные отдельно или совместно с клетками бета-TC6 (результаты не показаны).
Влияние совместного культивирования бета-TC6 и NCSC клеток на иммунофлуоресценцию кадгерина и бета-катенина
Кадгериновые соединения являются сильными медиаторами выживания бета-клеток [20] и компетентности NCSC [21], [22]. Затем, используя антитело к пан-кадгерину, мы проанализировали микроскопический вид соединений кадгерина в совместных культурах бета-TC6 / NCSC. Мы зарегистрировали накопление кадгерина на плазматической мембране соседних клеток, как между клетками бета-TC6 (рис. 6, стрелки), так и между NCSC, а также между клетками β-TC6 и NCSC (рис. 6, кружки).Также бета-катенин, внутриклеточная связь между кадгериновыми соединениями и клеточным цитоскелетом, демонстрировал аналогичную картину окрашивания (рис. 6). Мы наблюдали, что периферические клетки бета-ТС6, которые не были полностью окружены другими клетками, по окружности клеточного монослоя не обладали скоплениями кадгерина и бета-катенина (фигура 6, квадраты). Напротив, и кадгерин, и катенин в большом количестве присутствовали в клетках бета-ТС6, когда клетки были плотно окружены NCSC (рис. 6, кружки).Это указывает на то, что клетки бета-ТС6 образуют функциональные кадгериновые соединения с NCSC, которые заполняют всю окружность клеток. На паттерны иммунофлуоресценции бета-катенина и кадгерина в клетках бета-TC6 и NCSC цитокины не влияли (результаты не показаны).
Затем, используя антитело к пан-кадгерину, мы проанализировали микроскопический вид соединений кадгерина в совместных культурах бета-TC6 / NCSC. Мы зарегистрировали накопление кадгерина на плазматической мембране соседних клеток, как между клетками бета-TC6 (рис. 6, стрелки), так и между NCSC, а также между клетками β-TC6 и NCSC (рис. 6, кружки).Также бета-катенин, внутриклеточная связь между кадгериновыми соединениями и клеточным цитоскелетом, демонстрировал аналогичную картину окрашивания (рис. 6). Мы наблюдали, что периферические клетки бета-ТС6, которые не были полностью окружены другими клетками, по окружности клеточного монослоя не обладали скоплениями кадгерина и бета-катенина (фигура 6, квадраты). Напротив, и кадгерин, и катенин в большом количестве присутствовали в клетках бета-ТС6, когда клетки были плотно окружены NCSC (рис. 6, кружки).Это указывает на то, что клетки бета-ТС6 образуют функциональные кадгериновые соединения с NCSC, которые заполняют всю окружность клеток. На паттерны иммунофлуоресценции бета-катенина и кадгерина в клетках бета-TC6 и NCSC цитокины не влияли (результаты не показаны).
Рисунок 6. Клетки Beta-TC6 и NCSC образуют кадгериновые соединения.
Не-GFP-положительные клетки бета-TC6 культивировали либо отдельно (левые панели), либо вместе с GFP-положительными клетками NCSC (правые панели, зеленые клетки) на покровных стеклах, покрытых ламинином.Затем клетки фиксировали, пермеабилизировали и окрашивали на пан-кадгерин или бета-катенин (красный). Ядра были визуализированы Hoechst. Затем флуоресценцию фотографировали на флуоресцентном микроскопе с линзой 20x. Стрелки указывают положения соединений кадгерин / бета-катенин между клетками бета-TC6, а кружки указывают положения соединений между клетками бета-TC6 и NCSC. Квадратные головки указывают положения внешней окружности бета-ТС6, не окруженной NCSC. Масштабная линейка показывает расстояние 5 мкм.Результаты являются репрезентативными для трех независимых экспериментов.
https://doi.org/10.1371/journal.pone.0061828.g006
Ингибирование активности щелевых соединений не влияло на индуцированную совместным культивированием защиту от индуцированной цитокинами гибели клеток бета-TC6
Поскольку сообщалось, что щелевые соединения способствуют выживанию бета-клеток [23], мы выполнили определения жизнеспособности клеток на совместных культурах бета-TC6 клеток / NCSC с ингибитором щелевых соединений карбеноксолоном. Мы наблюдали, что индуцированная цитокинами гибель клеток бета-ТС6 не зависела от ингибитора щелевых соединений ни при культивировании только клеток бета-ТС6, ни при совместном культивировании клеток бета-ТС6 с NCSC (фигура 7).
Мы наблюдали, что индуцированная цитокинами гибель клеток бета-ТС6 не зависела от ингибитора щелевых соединений ни при культивировании только клеток бета-ТС6, ни при совместном культивировании клеток бета-ТС6 с NCSC (фигура 7).
Рис. 7. Ингибитор щелевых соединений не влияет на защиту от цитокинов, индуцированную совместным культивированием.
NCSC помещали на планшеты, покрытые ламинином, и через три дня добавляли клетки бета-TC6. Еще через 48 часов добавляли IL-1β (20 нг / мл) и IFN-γ (20 нг / мл) отдельно или в комбинации с ингибитором щелевого соединения карбеноксолоном (50 мкМ). Спустя 48 часов клетки метили йодидом пропидия, трипсинизировали и анализировали на гибель клеток проточной цитометрией. GFP-положительные клетки (NCSC) и GFP-отрицательные клетки регистрировали и анализировали отдельно.Результаты представляют собой средние значения ± SEM для трех отдельных наблюдений.
https://doi.org/10.1371/journal.pone.0061828.g007
Обсуждение
Предыдущие исследования сообщили о стимулирующем влиянии совместного культивирования NCSC и бета-клеток на пролиферацию бета-клеток [15] и функцию бета-клеток после трансплантации диабетическим мышам [13]. Кроме того, также сообщалось, что выживаемость бета-клеток в ответ на цитокины увеличивается при совместном культивировании с NCSC [14].Хотя благоприятное влияние совместного культивирования на пролиферацию бета-клеток требует прямых межклеточных контактов [15], остается неизвестным, опосредована ли защита от индуцированной цитокинами гибели растворимыми факторами или опосредована прямым межклеточным взаимодействием. контакт. Однако в настоящем исследовании мы наблюдаем, что для повышения жизнеспособности после воздействия цитокинов требуются прямые межклеточные контакты. Таким образом, одни и те же межклеточные контакты могут опосредовать не только усиление пролиферации бета-клеток, но также улучшение функции и выживаемости бета-клеток в ответ на цитокины.
Кроме того, также сообщалось, что выживаемость бета-клеток в ответ на цитокины увеличивается при совместном культивировании с NCSC [14].Хотя благоприятное влияние совместного культивирования на пролиферацию бета-клеток требует прямых межклеточных контактов [15], остается неизвестным, опосредована ли защита от индуцированной цитокинами гибели растворимыми факторами или опосредована прямым межклеточным взаимодействием. контакт. Однако в настоящем исследовании мы наблюдаем, что для повышения жизнеспособности после воздействия цитокинов требуются прямые межклеточные контакты. Таким образом, одни и те же межклеточные контакты могут опосредовать не только усиление пролиферации бета-клеток, но также улучшение функции и выживаемости бета-клеток в ответ на цитокины.
Природа прямых межклеточных контактов ранее не была выяснена. Здесь мы проанализировали соединения кадгерина, взаимодействия интегрин-ECM и щелевые соединения между NCSC, островковыми клетками и этими клетками в совместных культурах. Мы сообщаем, что бета-TC6 и клетки NCSC связаны с помощью соединений кадгерина. Эти структуры, вероятно, состоят из нейронального кадгерина (N-кадгерина) и / или эпителиального кадгерина (E-кадгерина), поскольку известно, что как бета-клетки, так и NCSC экспрессируют эти гомофильные адгезионные молекулы, в зависимости от стадии развития два типа клеток [24] — [26].Кадгериновые соединения между бета-клетками способствуют повышению выживаемости и функции [20], и возможно, что кадгериновые соединения между NCSC и бета-TC6 клетками оказывают сходные эффекты. Индуцированные кадгерином сигнальные пути, которые опосредуют устойчивость к гибели клеток, вероятно, очень сложны и зависят от контекста, но исследования показали увеличение притока кальция [27], снижение ядерной транслокации бета-катенина [28] и снижение фосфорилирования ERK [29], насколько это возможно. сигнальные события.
Мы сообщаем, что бета-TC6 и клетки NCSC связаны с помощью соединений кадгерина. Эти структуры, вероятно, состоят из нейронального кадгерина (N-кадгерина) и / или эпителиального кадгерина (E-кадгерина), поскольку известно, что как бета-клетки, так и NCSC экспрессируют эти гомофильные адгезионные молекулы, в зависимости от стадии развития два типа клеток [24] — [26].Кадгериновые соединения между бета-клетками способствуют повышению выживаемости и функции [20], и возможно, что кадгериновые соединения между NCSC и бета-TC6 клетками оказывают сходные эффекты. Индуцированные кадгерином сигнальные пути, которые опосредуют устойчивость к гибели клеток, вероятно, очень сложны и зависят от контекста, но исследования показали увеличение притока кальция [27], снижение ядерной транслокации бета-катенина [28] и снижение фосфорилирования ERK [29], насколько это возможно. сигнальные события.
Помимо соединений кадгерина, мы в настоящее время также наблюдали, что прямые взаимодействия бета-TC6 / NCSC приводят к увеличению продукции ламинина и снижению передачи сигналов ERK / FAK. Неясно, почему присутствие NCSCs способствует снижению передачи сигналов ERK / FAK, но мы можем предвидеть два возможных объяснения. Во-первых, с использованием клеток островков поджелудочной железы сообщалось, что связывание альфа-интегрина с коллагеном IV стимулирует активацию ERK, тогда как связывание интегрина с ламинином вместо этого подавляет активацию ERK за счет снижения передачи сигналов рецептора фактора роста фибробластов-1 (FGFR1) [18] .Кроме того, клетки, продуцирующие инсулин INS-1, реагировали на коллаген IV, но не на ламинин, стимулированным фосфорилированием FAK [30]. Следовательно, ламинин, полученный из NCSC, мог существенно изменить состав внеклеточного матрикса, так что эффекты коллагена исчезают, а фосфорилирование FAK / ERK ослабляется. Во-вторых, также возможно, что сильные кадгериновые соединения между β-TC6 клетками и NCSCs посредством цис- и транслатеральных взаимодействий физически конкурируют с фокальными адгезиями β-TC6 клеток, что приводит к снижению передачи сигналов FAK / ERK.
Неясно, почему присутствие NCSCs способствует снижению передачи сигналов ERK / FAK, но мы можем предвидеть два возможных объяснения. Во-первых, с использованием клеток островков поджелудочной железы сообщалось, что связывание альфа-интегрина с коллагеном IV стимулирует активацию ERK, тогда как связывание интегрина с ламинином вместо этого подавляет активацию ERK за счет снижения передачи сигналов рецептора фактора роста фибробластов-1 (FGFR1) [18] .Кроме того, клетки, продуцирующие инсулин INS-1, реагировали на коллаген IV, но не на ламинин, стимулированным фосфорилированием FAK [30]. Следовательно, ламинин, полученный из NCSC, мог существенно изменить состав внеклеточного матрикса, так что эффекты коллагена исчезают, а фосфорилирование FAK / ERK ослабляется. Во-вторых, также возможно, что сильные кадгериновые соединения между β-TC6 клетками и NCSCs посредством цис- и транслатеральных взаимодействий физически конкурируют с фокальными адгезиями β-TC6 клеток, что приводит к снижению передачи сигналов FAK / ERK. Сильные кадгериновые соединения связаны с поддерживаемым эпителиальным фенотипом и устойчивостью к переходу от эпителия к мезенхиме [31]. Кроме того, также сообщалось, что взаимодействия клеток со специфическими ламининами противодействуют переходу эпителия в мезенхиму, тем самым предотвращая де-дифференцировку и потерю фенотипа бета-клеток [32], [33]. Т.о., как кадгериновые соединения, так и продуцируемый NCSCs ламинин могут модулировать передачу сигналов интегрина бета-TC6 клеток, так что передача сигналов ERK и FAK подавляется, эпителиально-мезенхимальный переход ингибируется и выживаемость сохраняется.Возникает соблазн предположить, что два альтернативных объяснения, то есть повышенный уровень кадгерина и повышенный ламинин, действуют синергетически, чтобы сохранить функцию бета-клеток и выжить. В самом деле, активированные ламинином интегрины, как сообщается, стабилизируют соединения E-cadherin в др. Клеточных системах [34].
Сильные кадгериновые соединения связаны с поддерживаемым эпителиальным фенотипом и устойчивостью к переходу от эпителия к мезенхиме [31]. Кроме того, также сообщалось, что взаимодействия клеток со специфическими ламининами противодействуют переходу эпителия в мезенхиму, тем самым предотвращая де-дифференцировку и потерю фенотипа бета-клеток [32], [33]. Т.о., как кадгериновые соединения, так и продуцируемый NCSCs ламинин могут модулировать передачу сигналов интегрина бета-TC6 клеток, так что передача сигналов ERK и FAK подавляется, эпителиально-мезенхимальный переход ингибируется и выживаемость сохраняется.Возникает соблазн предположить, что два альтернативных объяснения, то есть повышенный уровень кадгерина и повышенный ламинин, действуют синергетически, чтобы сохранить функцию бета-клеток и выжить. В самом деле, активированные ламинином интегрины, как сообщается, стабилизируют соединения E-cadherin в др. Клеточных системах [34].
Щелевые соединения также являются прямыми межклеточными контактами, которые являются потенциальными медиаторами выживания бета-клеток [23]. В бета-клетках коннексин 36 образует функциональные щелевые соединения, которые защищают от гибели клеток, вызванной цитокинами [23].С другой стороны, сообщалось, что NCSCs экспрессируют коннексин 43 до дифференцировки в зрелые типы клеток [35]. Поскольку существует множество примеров функциональных гетеромерных щелевых контактов, возможно, что они также присутствуют в наблюдаемых в настоящее время соединениях бета-TC6 / NCSC. Однако мы не наблюдали эффекта ингибитора щелевых соединений карбеноксолона на выживаемость клеток бета-TC6, что указывает на то, что предполагаемые щелевые соединения бета-TC6 / NCSC не опосредуют улучшение выживаемости клеток бета-TC6.
В бета-клетках коннексин 36 образует функциональные щелевые соединения, которые защищают от гибели клеток, вызванной цитокинами [23].С другой стороны, сообщалось, что NCSCs экспрессируют коннексин 43 до дифференцировки в зрелые типы клеток [35]. Поскольку существует множество примеров функциональных гетеромерных щелевых контактов, возможно, что они также присутствуют в наблюдаемых в настоящее время соединениях бета-TC6 / NCSC. Однако мы не наблюдали эффекта ингибитора щелевых соединений карбеноксолона на выживаемость клеток бета-TC6, что указывает на то, что предполагаемые щелевые соединения бета-TC6 / NCSC не опосредуют улучшение выживаемости клеток бета-TC6.
Таким образом, учитывая хорошо установленную роль соединений кадгерина в обеспечении выживания, вполне вероятно, что наблюдаемые в настоящее время прямые межклеточные контакты участвуют в защите от индуцированной цитокинами гибели клеток.Чтобы еще больше подтвердить эти находки, он, по-видимому, сначала имеет высокую мотивацию для блокирования соединений NCSC-to-beta-TC6 кадгерина с использованием антител, разрушающих кадгерин [27]. К сожалению, такая стратегия также нарушит соединения бета-ТС6 с бета-ТС6, что, вероятно, сделало бы интерпретацию любых предполагаемых результатов весьма проблематичной. Следовательно, необходимо разработать другие экспериментальные подходы для лучшего понимания прямых контактов NCSC / beta-TC6 клеток. Между тем, возможно, что испытания трансплантации островков выиграют от совместной трансплантации NCSC или от манипуляций, которые способствуют соединениям кадгерина бета-клеток с соседними клетками.
К сожалению, такая стратегия также нарушит соединения бета-ТС6 с бета-ТС6, что, вероятно, сделало бы интерпретацию любых предполагаемых результатов весьма проблематичной. Следовательно, необходимо разработать другие экспериментальные подходы для лучшего понимания прямых контактов NCSC / beta-TC6 клеток. Между тем, возможно, что испытания трансплантации островков выиграют от совместной трансплантации NCSC или от манипуляций, которые способствуют соединениям кадгерина бета-клеток с соседними клетками.
Вклад авторов
Задумал и спроектировал эксперименты: NW ENK. Проведены эксперименты: АН КТ СВ. Проанализированы данные: АН КТ СВ ЭНК СЗ. Написал бумагу: ENK NW.
Список литературы
- 1. Эйзирик Д.Л., Мандруп-Поулсен Т. (2000) Выбор смерти — передача сигнала иммуно-опосредованного апоптоза бета-клеток. Диабетология 44: 2115–2133.
- 2.
Суарес-Пинзон В., Соренсен О., Блекли Р. К., Эллиотт Дж. Ф., Раджот Р. В. и др.(1999) Разрушение бета-клеток у мышей NOD коррелирует с экспрессией Fas (CD95) на бета-клетках и экспрессией провоспалительных цитокинов в островках. Диабет 48: 21–28.
- 3. Уно С., Имагава А., Окита К., Саяма К., Мориваки М. и др. (2007) Макрофаги и дендритные клетки, инфильтрирующие островки с бета-клетками или без них, продуцируют фактор некроза опухоли альфа у пациентов с недавно начавшимся диабетом 1 типа. Диабетология 50: 596–601.
- 4. Welsh N (1996). Индуцированное интерлейкином-1бета образование церамида и диацилглицерина может приводить к активации c-Jun-Nh3-терминальной киназы и фактора транскрипции ATF2 в линии инсулин-продуцирующих клеток RINm5F.J Biol Chem 271: 8307–8312.
- 5. Ларсен С.М., Вадт К.А., Юль Л.Ф., Андерсен Х.У., Карлсен А.Е. и др. (1998) Индуцированный интерлейкином-1бета синтез NO островков поджелудочной железы крыс требует как p38, так и регулируемых внеклеточными сигналами митоген-активируемых протеинкиназ киназы 1/2. J Biol Chem 273: 15294–15300.
- 6.
Flodström M, Welsh N, Eizirik DL (1996) Цитокины активируют ядерный фактор kB (NF-kappaB) и индуцируют производство оксида азота в островках поджелудочной железы человека. FEBS Lett 385: 4–6.
- 7. Mokhtari D, Barbu A, Mehmeti I, Vercamer C, Welsh N (2009) Сверхэкспрессия субъединицы ядерного фактора — {kappa} B c-Rel защищает от гибели островковых клеток человека in vitro. Am J Physiol Endocrinol Metab 297: E1067-1077.
- 8. Райан Э.А., Пати Б.В., старший PA, Бигам Д., Альфадли Э. и др. (2005) Пятилетнее наблюдение после клинической трансплантации островков. Диабет 54: 2060–2069.
- 9. Davalli AM, Scaglia L, Zangen DH, Zangen DH, Hollister J, et al.(1996) Уязвимость островков в ближайшем посттрансплантационном периоде. Динамические изменения в структуре и функциях. Диабет 45: 1161–1167.
- 10. Nekrep N, Wang J, Miyatsuka T, German MS (2008) Сигналы от нервного гребня регулируют массу бета-клеток в поджелудочной железе. Развитие 135: 2151–2160.
- 11.
Козлова Е.Н., Янссон Л. (2005) Взаимодействия in vitro между инсулин-продуцирующими бета-клетками и ганглиями дорзального корешка эмбриона. Поджелудочная железа 31: 380–384.
- 12.Козлова Е.Н., Янссон Л. (2009) Дифференциация и миграция стволовых клеток нервного гребня стимулируются островками поджелудочной железы. Neuroreport 20: 833–838.
- 13. Олеруд Дж., Канайкина Н., Василовская С., Кинг Д., Сандберг М. и др. (2009) Стволовые клетки нервного гребня увеличивают пролиферацию бета-клеток и улучшают функцию островков совместно трансплантированных островков поджелудочной железы мыши. Диабетология 52: 2594–2601.
- 14. Ngamjariyawat A, Turpaev K, Welsh N, Kokzlova EN (2012) Совместное культивирование инсулин-продуцирующих клеток RIN5AH со стволовыми клетками нервного гребня частично защищает от гибели клеток, индуцированной цитокинами.Поджелудочная железа 41: 490–492.
- 15.
Grouwels G, Vasylovska S, Olerud J, Leuckx G, Ngamjariyawat A, et al. (2012) Дифференцирующиеся стволовые клетки нервного гребня индуцируют пролиферацию культивируемых бета-клеток островков грызунов. Диабетология. 55: 2016–25.
- 16. Hjerling-Leffler J, Marmigere F, Heglind M, Cederberg A, Koltzenburg M и др. (2005) Граничный колпачок: источник стволовых клеток нервного гребня, которые генерируют несколько подтипов сенсорных нейронов. Развитие 132: 2623–2632.
- 17.Альдскогиус Х., Беренс С., Канайкина Н., Ляховицкая А., Медвинский А. и др. (2009) Регулирование дифференцировки стволовых клеток нервного гребня пограничной крышки после трансплантации. Стволовые клетки 27: 1592–1603.
- 18. Kilkenny DM, Rocheleau JV (2008) Передача сигналов рецептора-1 фактора роста фибробастов в b-клетках панкреатических островков модулируется внеклеточным матриксом. Мол эндоцинол 22: 196–205.
- 19.
Парно Дж., Хаммер Э., Руиллер Д. Г., Арманет М., Халбан П. А. и др.(2006) Блокада взаимодействия b1 интергрина с ламинином-5 влияет на распространение и секрецию инсулина b-клетками крысы, прикрепленными к внеклеточному матриксу. Диабет 55: 1413–1420.
- 20. Парно Г., Гонель-Гисперт С., Морель П., Джованнони Л., Мюллер Ю. Д. и др. (2011) Вовлечение кадгерина защищает человеческие В-клетки от апоптоза. Эндокринология 152: 4601–4609.
- 21. Курчо К.Л., Маурер Дж., МакКаун С.Дж., Каттаросси Дж., Симадамор Ф. и др. (2010) Раннее приобретение компетенции Neural Crest во время нейрализации hESCs PLoS ONE.5: e13890.
- 22. Kléber M, Lee HY, Wurdak H, Buchstaller J, Riccomagno MM и др. (2005) Поддержание стволовых клеток нервного гребня с помощью комбинаторной передачи сигналов Wnt и BMP. J Cell Biol 169: 309–320.
- 23. Klee P, Allagnat F, Pontes H, Cerderroth M, Charollais A и др. (2011) Коннексины защищают В-клетки поджелудочной железы мыши от апоптоза. Дж. Клин Инвест 121: 4870–4879.
- 24. Dahl U, Sjödin A, Semb H (1996) Кадгерины регулируют агрегацию бета-клеток поджелудочной железы in vivo.Разработка 122: 2895–2902.
- 25.
Мур Р. , Ларю Л. (2004) Молекулы клеточной поверхности и онтогенез ствола нервного гребня: перспектива. Рождение Def Res 72: 140–150.
- 26. Йоханссон Дж. К., Восс Ю., Кесаван Г., Костецкий И., Виеруп Н. и др. (2010) N-кадгерин незаменим для развития поджелудочной железы, но необходим для обновления гранул бета-клеток. Бытие 48: 374–381.
- 27. Hauge-Evans AC, Squires PE, Persaud SJ, Jones PM (1999) Ответы бета-клеток поджелудочной железы на бета-клетки на стимулы питательных веществ: усиление Ca2 + и секреторных резонансов инсулина псевдоостровок MIN6.Диабет 48: 1402–1408.
- 28. Herzig M, Savarese F, Novatchkova M, Semb H, Christofori G (2007) Прогрессирование опухоли, вызванное потерей E-кадгерина, независимо от передачи сигналов Wnt, опосредованной бета-катенином / Tcf. Онкоген 26: 2290–2298.
- 29.
Iwashita J, Ose K, Ito H, Murata J, Abe T (2011) Ингибирование E-кадгерин-зависимого межклеточного контакта способствует производству муцина MUC5AC через активацию рецепторов эпидермального фактора роста. Biosci Biotechnol Biochem 75: 688–693.
- 30. Кришнамурти М., Ли Дж., Аль-Масири М., Ван Р. (2008) Экспрессия и функция интегринов алфавита 1 в бета-клетках поджелудочной железы (INS-1). J Cell Commun Signal 2: 67–79.
- 31. Heuberger J, Birchmeier W (2010) Взаимодействие опосредованной кадгерином клеточной адгезии и канонической передачи сигналов Wnt. Колд-Спринг-Харб Perspect Biol 2: a002915.
- 32. Chapman HA (2012) Эпителиальные реакции на повреждение легких: роль внеклеточного матрикса. Proc Am Thorac Soc 9: 89–95.
- 33. Banerjee M, Virtanen I, Palgli J, Korsgren O, Otonkoski T (2012) Пролиферация и пластичность бета-клеток человека на физиологически встречающихся изоформах ламинина. Эндокринол клеток Mol 355: 78–86.
- 34. Stipp CS (2010). Ламинин-связывающие интегрины и их тетраспаниновые партнеры как потенциальные антиметастатические мишени. Опыт Рев Мол Мед 12.
- 35.
Ли Дж. , Хаббс Х.В., Эйбергер Дж., Виллеке К., Дермицель Р. и др. (2006) Анализ экспрессии коннексина во время развития шванновских клеток мышей идентифицирует коннексин 29 как новый маркер перехода нервного гребня в клетки-предшественники.GLIA 55: 93–103.
 Диабет 48: 21–28.
Диабет 48: 21–28. FEBS Lett 385: 4–6.
FEBS Lett 385: 4–6. Поджелудочная железа 31: 380–384.
Поджелудочная железа 31: 380–384. Диабетология. 55: 2016–25.
Диабетология. 55: 2016–25.
 , Ларю Л. (2004) Молекулы клеточной поверхности и онтогенез ствола нервного гребня: перспектива. Рождение Def Res 72: 140–150.
, Ларю Л. (2004) Молекулы клеточной поверхности и онтогенез ствола нервного гребня: перспектива. Рождение Def Res 72: 140–150. Biosci Biotechnol Biochem 75: 688–693.
Biosci Biotechnol Biochem 75: 688–693. , Хаббс Х.В., Эйбергер Дж., Виллеке К., Дермицель Р. и др. (2006) Анализ экспрессии коннексина во время развития шванновских клеток мышей идентифицирует коннексин 29 как новый маркер перехода нервного гребня в клетки-предшественники.GLIA 55: 93–103.
, Хаббс Х.В., Эйбергер Дж., Виллеке К., Дермицель Р. и др. (2006) Анализ экспрессии коннексина во время развития шванновских клеток мышей идентифицирует коннексин 29 как новый маркер перехода нервного гребня в клетки-предшественники.GLIA 55: 93–103.Догоняют ли азиатские драконы и тигры?
Автор
Abstract
В данной статье исследуется процесс «наверстывания» доходов на душу населения у так называемых азиатских драконов и тигров. Он пополняет литературу несколькими способами. Во-первых, в нем проверяется гипотеза наверстывания с использованием самого длительного периода времени, который когда-либо рассматривался, с 1870 по 2014 год. Во-вторых, он документирует опыт этих двух групп стран и дает им возможные объяснения.В-третьих, анализ статьи использует алгоритм Кеджривала и Перрона (J Time Ser Anal 31: 305–328, 2010), который позволяет нам эндогенно оценивать множественные структурные сдвиги на уровне и тенденцию ряда без предварительного знания их интеграции. уровень. Это превосходит технические проблемы предыдущих эмпирических исследований. В-четвертых, он исследует, как азиатский финансовый кризис повлиял на процесс наверстывания отставания в экономиках Драконов и Тигров.
уровень. Это превосходит технические проблемы предыдущих эмпирических исследований. В-четвертых, он исследует, как азиатский финансовый кризис повлиял на процесс наверстывания отставания в экономиках Драконов и Тигров.
Рекомендуемое цитирование
DOI: 10.1007 / s00181-018-1517-5
Скачать полный текст от издателя
Поскольку доступ к этому документу ограничен, вы можете поискать его другую версию.
Ссылки на IDEAS
- Барри Эйченгрин, Донхён Парк и Кванхо Шин, 2012 г.« Когда быстрорастущие экономики замедляются: международные свидетельства и последствия для Китая »,
Азиатские экономические документы, MIT Press, vol. 11 (1), страницы 42-87, Winter / Sp.
- Каррион-и-Сильвестр, Хосеп Луис и Ким, Дукпа и Перрон, Пьер, 2009 г. « Тесты единичного корня на основе Gls с множественными структурными нарушениями как при нулевой, так и при альтернативной гипотезе », Эконометрическая теория, Cambridge University Press, vol. 25 (6), страницы 1754-1792, декабрь.
- Роберт Э.Лукас-младший, 2015. « Человеческий капитал и рост », Американский экономический обзор, Американская экономическая ассоциация, т. 105 (5), страницы 85-88, май.
- Ли, Чон-Вха и Хонг, Кисок, 2012 г. « Экономический рост в Азии: детерминанты и перспективы », Япония и мировая экономика, Elsevier, vol. 24 (2), страницы 101-113.
- Perron, Pierre & Zhu, Xiaokang, 2005. « Структурные разрывы с детерминированными и стохастическими тенденциями ,» Журнал эконометрики, Elsevier, т.129 (1-2), страницы 65-119.
- Бернард, Эндрю Б. и Дурлауф, Стивен Н., 1996.
« Интерпретация тестов гипотезы сходимости ,»
Журнал эконометрики, Elsevier, т. 71 (1-2), страницы 161-173.
- Стразичич, Марк С. и Ли, Джунсу и Дэй, Эдвард, 2004. « Сходятся ли доходы в странах ОЭСР? Данные временного ряда с двумя структурными сдвигами », Журнал макроэкономики, Elsevier, vol. 26 (1), страницы 131–145, март.
- Мохитош Кеджривал и Пьер Перрон, 2010 г.« Последовательная процедура для определения количества разрывов тренда с интегрированной или стационарной составляющей шума », Журнал анализа временных рядов, Wiley Blackwell, vol. 31 (5), страницы 305-328, сентябрь.
- Бернард, Эндрю Б. и Дурлауф, Стивен Н., 1995. « Конвергенция в международной продукции », Журнал прикладной эконометрики, John Wiley & Sons, Ltd., вып. 10 (2), страницы 97-108, апрель-июнь.
- Тарлок Сингх, 2010 г. « Причина экономического роста в международной торговле? Обзор «, Мировая экономика, Wiley Blackwell, vol.33 (11), страницы 1517-1564, ноябрь.
- Мохитош Кеджривал и Клод Лопес, 2013.
« основных единиц, сдвигов уровней и сдвигов в тенденциях производства на душу населения: надежная оценка »,
Эконометрические обзоры, Taylor & Francis Journals, vol. 32 (8), страницы 892-927, ноябрь.
- Мохитош Кеджривал и Клод Лопес, 2009. «Корни единиц, сдвиги уровней и сдвиги в тенденциях производства на душу населения: надежная оценка », Рабочие документы по экономике Университета Пердью 1227, Университет Пердью, факультет экономики.
- Кеджривал М. и Лопес К., 2011. «Корни единиц, сдвиги уровней и сдвиги в тенденциях производства на душу населения: надежная оценка », Рабочие бумаги 334, Банк Франции.
- Мохитош Кеджривал и Клод Лопес, 2010 г. « корней единиц, сдвиги уровней и сдвиги в тенденциях в выпуске на душу населения: надежная оценка », Университет Цинциннати, Серия рабочих документов по экономике 2010-02, Университет Цинциннати, факультет экономики.
- Кеджривал, Мохитош и Лопес, Клод, 2010 г.«Корни единиц, сдвиги уровней и сдвиги в тенденциях производства на душу населения: надежная оценка », Бумага MPRA 25204, Университетская библиотека Мюнхена, Германия.
- Ли, Qing & Papell, Дэвид, 1999. « Конвергенция международных результатов Данные временных рядов для 16 стран ОЭСР ,»
Международный обзор экономики и финансов, Elsevier, vol. 8 (3), страницы 267-280, сентябрь.
- Эванс, Пол и Ким, Джи Ук, 2011. « Стохастическая конвергенция скорости наверстывания и множественные структурные сдвиги в странах Азии », Economics Letters, Elsevier, vol.111 (3), страницы 260-263, июнь.
- Датта, Анусуа, 2003 г. « Тесты сходимости временных рядов и переходной динамики ,» Economics Letters, Elsevier, vol. 81 (2), страницы 233-240, ноябрь.
- Серена Нг и Пьер Перрон, 2001. « Выбор длины LAG и построение тестов единичного корня с хорошим размером и мощностью », Econometrica, Econometric Society, vol. 69 (6), страницы 1519-1554, ноябрь.
- Грегори Мэнкью, 1995. « Рост наций », Документы Брукингса об экономической деятельности, Программа экономических исследований, Институт Брукингса, т.26 (1, 25-я А), страницы 275-326.
- Перрон, Пьер и Родригес, Габриэль, 2003 г. « Детрендинг GLS, эффективные тесты единичного корня и структурные изменения »,
Журнал эконометрики, Elsevier, т. 115 (1), страницы 1-27, июль.
- Элвин Янг, 1995. « Тирания чисел: противостояние статистическим реалиям восточноазиатского опыта экономического роста «, Ежеквартальный журнал экономики, Oxford University Press, vol. 110 (3), страницы 641-680.
- Раделет Стивен и Сакс Джеффри и Ли Чон-Ва, 2001.» Детерминанты и перспективы экономического роста в Азии ,» Международный экономический журнал, Taylor & Francis Journals, vol. 15 (3), страницы 1-29.
- Ногера, Хосе, 2013. « Цены на нефть: разрывы и тенденции », Экономика энергетики, Elsevier, vol. 37 (C), страницы 60-67.
- Джейкоб Брауде и Игаль Менаше, 2011 г. « Азиатское чудо: капиталоемкое структурное изменение? », Журнал международной торговли и экономического развития, Taylor & Francis Journals, vol.20 (1), страницы 31-51.

 71 (1-2), страницы 161-173.
71 (1-2), страницы 161-173. 32 (8), страницы 892-927, ноябрь.
32 (8), страницы 892-927, ноябрь. « Конвергенция международных результатов Данные временных рядов для 16 стран ОЭСР ,»
Международный обзор экономики и финансов, Elsevier, vol. 8 (3), страницы 267-280, сентябрь.
« Конвергенция международных результатов Данные временных рядов для 16 стран ОЭСР ,»
Международный обзор экономики и финансов, Elsevier, vol. 8 (3), страницы 267-280, сентябрь. « Детрендинг GLS, эффективные тесты единичного корня и структурные изменения »,
Журнал эконометрики, Elsevier, т. 115 (1), страницы 1-27, июль.
« Детрендинг GLS, эффективные тесты единичного корня и структурные изменения »,
Журнал эконометрики, Elsevier, т. 115 (1), страницы 1-27, июль.Самые популярные товары
Это элементы, которые чаще всего цитируют те же работы, что и эта, и цитируются в тех же работах, что и эта.
- Атану Гошрей и Фаиза Хан, 2015. « Новые эмпирические данные о конвергенции доходов «, Эмпирическая экономика, Springer, т. 49 (1), страницы 343-361, август.
- Харви, Дэвид И. и Лейборн, Стивен Дж.И Тейлор, А. Роберт, 2012. «Тестирование корневого модуля при локальном прорыве в тенденции », Журнал эконометрики, Elsevier, т. 167 (1), страницы 140-167.
- Нуно Собрейра и Луис К. Нуньес и Пауло М. Родригес, 2014 г. « Характеризация траекторий экономического роста на основе новых тестов на структурные изменения », Экономическое расследование, Международная западная экономическая ассоциация, т. 52 (2), страницы 845-861, апрель.
- Tunali, Çidem Börke & Yilanci, Veli, 2010. « Сходятся или расходятся доходы на душу населения в странах MENA? », Physica A: Статистическая механика и ее приложения, Elsevier, vol.389 (21), страницы 4855-4862.
- Мацуки, Такаши, 2019.
« Конвергенция выпуска на душу населения в азиатских странах: данные ковариатного критерия единичного корня с эндогенным структурным разрывом »,
Экономическое моделирование, Elsevier, vol. 82 (C), страницы 99-118.
- Серранито, Франциско, 2013 г. « Гетерогенная технология и гипотеза технологического наверстывания: теория и оценка на примере стран MENA », Экономическое моделирование, Elsevier, vol. 30 (C), страницы 685-697.
- Амели Шарль и Оливье Дарне и Жан-Франсуа Оарау, 2019 г. « Насколько устойчива Ла Реюньон с точки зрения привлекательности международного туризма: оценка на основе тестов на единичный корень со структурными сдвигами в 1981–2015 гг. », Прикладная экономика, Taylor & Francis Journals, vol. 51 (24), страницы 2639-2653, май.
- Омид Ранджбар и Цангьяо Чанг, Чиен-Чианг Ли и Захра Мила Эльми, 2018. « Процесс наверстывания в странах с переходной экономикой », Экономические изменения и реструктуризация, Springer, т.51 (3), страницы 249-278, август.
- Франсиско Дельгадо и Мария Хосе Пресно, 2011 г.
« Конвергенция фискального давления в ЕС: подход временного ряда »,
Прикладная экономика, Taylor & Francis Journals, vol. 43 (28), страницы 4257-4267.
- Собрейра, Нуно и Нунес, Луис К. и Родригес, Пауло М. М., 2012. « Неоклассическая, полуэндогенная или эндогенная теория роста? Доказательства, основанные на новых тестах на структурные изменения », Рабочие документы Insper wpe_291, Рабочий документ Insper, Insper Instituto de Ensino e Pesquisa.
- Пьер Перрон, 2017. « Корневой элемент и структурные разрывы ,» Эконометрика, MDPI, Open Access Journal, vol. 5 (2), страницы 1-3, май.
- Стенгос, Танасис и Язган, М. Эге, 2014. « Постоянство в конвергенции ,» Макроэкономическая динамика, Cambridge University Press, vol. 18 (4), страницы 753-782, июнь.
- repec: kap: iaecre: v: 17: y: 2011: i: 3: p: 315-333 не указан в IDEAS
- Кунадо, Х. и Перес де Грасиа, Ф., 2006. « Реальная конвергенция в Африке во второй половине 20 века », Журнал экономики и бизнеса, Elsevier, vol.58 (2), страницы 153-167.
- Нильс Халдруп, Робинсон Круз, Тимо Терасвирта и Расмус Т. Варнесков, 2013.
« Единичные корни, нелинейности и структурные разрывы ,»
Chapters, in: Nigar Hashimzade & Michael A. Thornton (ed.), Handbook of Research Methods and Applications in Empirical Macroeconomics, Chapter 4, pages 61-94,
Эдвард Элгар Паблишинг.
- Батаа, Эрденебат и Иззельдин, Марван и Осборн, Дениз Р., 2016. « Изменения на мировом рынке нефти », Экономика энергетики, Elsevier, vol.56 (C), страницы 161-176.
- Эрденебат Батаа, Эндрю Вивиан и Марк Вохар, 2019. « Изменения во взаимосвязи между краткосрочной процентной ставкой, инфляцией и ростом: данные из Великобритании, 1820–2014 гг. », Бюллетень экономических исследований, Wiley Blackwell, т. 71 (4), страницы 616-640, октябрь.
- Мария Хесус Эррериас и Висенте Ортс, 2011 г. « Движущие силы роста Китая », Экономика переходного периода, Европейский банк реконструкции и развития, вып.19 (1), страницы 79-124, январь.
- Хункал Кунадо и Луис А. Хиль-Алана и Фернандо Перес де Грасиа, 2006. « Дополнительные эмпирические доказательства реальной конвергенции: дробно-интегрированный подход «,
Обзор мировой экономики (Weltwirtschaftliches Archiv), Springer; Institut für Weltwirtschaft (Кильский институт мировой экономики), т. 142 (1), страницы 67-91, апрель.
- Омид Ранджбар и Цангяо Чанг, Цзян-Чианг Ли и Захра (Мила) Эльми, 2016. « Возобновление дебатов о конвергенции, когда резкие разрывы и плавные сдвиги, среда, 1870-2010 гг. », Иранское экономическое обозрение (IER), экономический факультет Тегеранского университета.Тегеран, Иран, т. 20 (3), страницы 356-377, Summer.
- Зербо, Элеазар и Дарне, Оливье, 2019. « О стационарности выбросов CO2 в странах ОЭСР и БРИКС: метод последовательного тестирования », Экономика энергетики, Elsevier, vol. 83 (C), страницы 319-332.
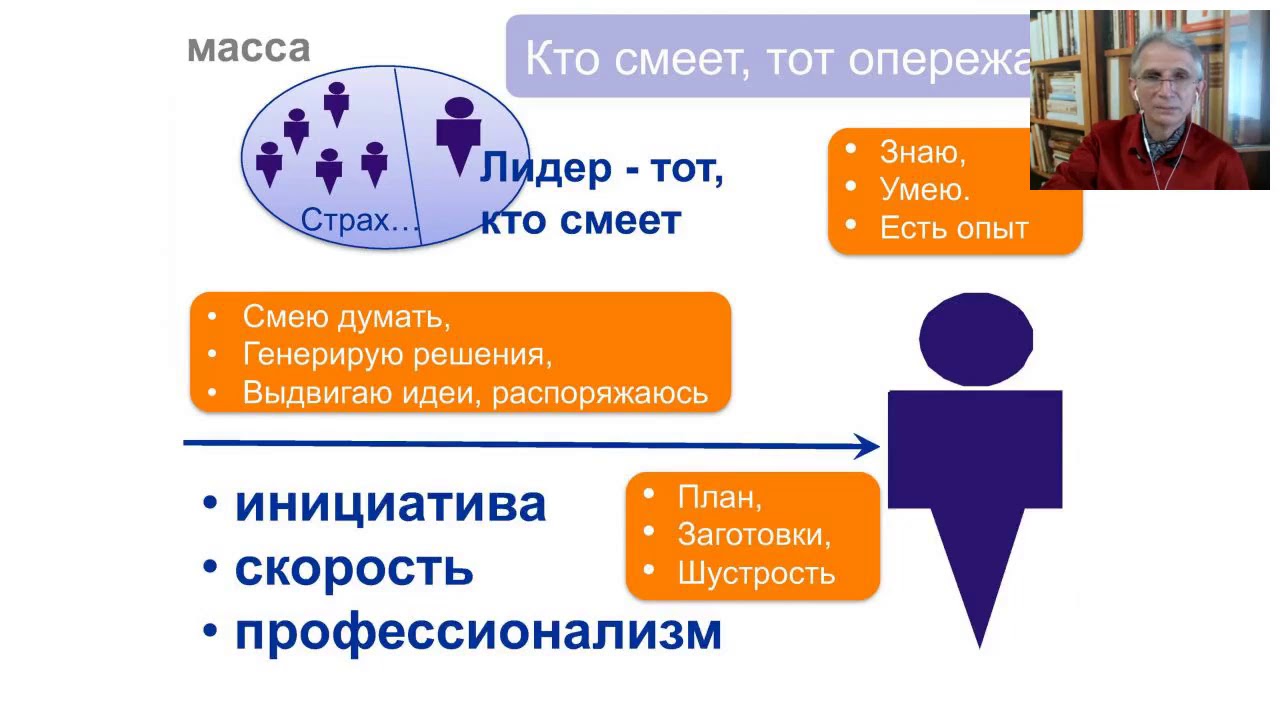 82 (C), страницы 99-118.
82 (C), страницы 99-118. 43 (28), страницы 4257-4267.
43 (28), страницы 4257-4267. Варнесков, 2013.
« Единичные корни, нелинейности и структурные разрывы ,»
Chapters, in: Nigar Hashimzade & Michael A. Thornton (ed.), Handbook of Research Methods and Applications in Empirical Macroeconomics, Chapter 4, pages 61-94,
Эдвард Элгар Паблишинг.
Варнесков, 2013.
« Единичные корни, нелинейности и структурные разрывы ,»
Chapters, in: Nigar Hashimzade & Michael A. Thornton (ed.), Handbook of Research Methods and Applications in Empirical Macroeconomics, Chapter 4, pages 61-94,
Эдвард Элгар Паблишинг. « Дополнительные эмпирические доказательства реальной конвергенции: дробно-интегрированный подход «,
Обзор мировой экономики (Weltwirtschaftliches Archiv), Springer; Institut für Weltwirtschaft (Кильский институт мировой экономики), т. 142 (1), страницы 67-91, апрель.
« Дополнительные эмпирические доказательства реальной конвергенции: дробно-интегрированный подход «,
Обзор мировой экономики (Weltwirtschaftliches Archiv), Springer; Institut für Weltwirtschaft (Кильский институт мировой экономики), т. 142 (1), страницы 67-91, апрель.Подробнее об этом товаре
Ключевые слова
Догонять; Структурные разрывы; Азиатские драконы; Азиатские тигры;Все эти ключевые слова.

Классификация JEL:
- C22 — Математические и количественные методы — — Модели с одним уравнением; Одиночные переменные — — — Модели временных рядов; Динамические квантильные регрессии; Модели динамического воздействия лечения; Диффузионные процессы
- O11 — Экономическое развитие, инновации, технологические изменения и рост — — Экономическое развитие — — — Макроэкономический анализ экономического развития
- O55 — Экономическое развитие, инновации, технологические изменения и рост — — Страновые исследования в масштабах экономики — — — Африка
Статистика
Доступ и загрузка статистикиИсправления
Все материалы на этом сайте предоставлены соответствующими издателями и авторами.Вы можете помочь исправить ошибки и упущения. При запросе исправления укажите идентификатор этого элемента: RePEc: spr: empeco: v: 57: y: 2019: i: 2: d: 10.1007_s00181-018-1517-5 . См. Общую информацию о том, как исправить материал в RePEc.
По техническим вопросам, касающимся этого элемента, или для исправления его авторов, названия, аннотации, библиографической информации или информации для загрузки, обращайтесь: (Sonal Shukla) или (Springer Nature Abstracting and Indexing). Общие контактные данные провайдера: http: // www.springer.com .
Если вы создали этот элемент и еще не зарегистрированы в RePEc, мы рекомендуем вам сделать это здесь. Это позволяет связать ваш профиль с этим элементом. Это также позволяет вам принимать возможные ссылки на этот элемент, в отношении которого мы не уверены.
Если CitEc распознал ссылку, но не связал с ней элемент в RePEc, вы можете помочь с этой формой .
Если вам известно об отсутствующих элементах, цитирующих этот элемент, вы можете помочь нам создать эти ссылки, добавив соответствующие ссылки таким же образом, как указано выше, для каждого элемента ссылки.Если вы являетесь зарегистрированным автором этого элемента, вы также можете проверить вкладку «Цитаты» в своем профиле службы авторов RePEc, поскольку там могут быть некоторые цитаты, ожидающие подтверждения.