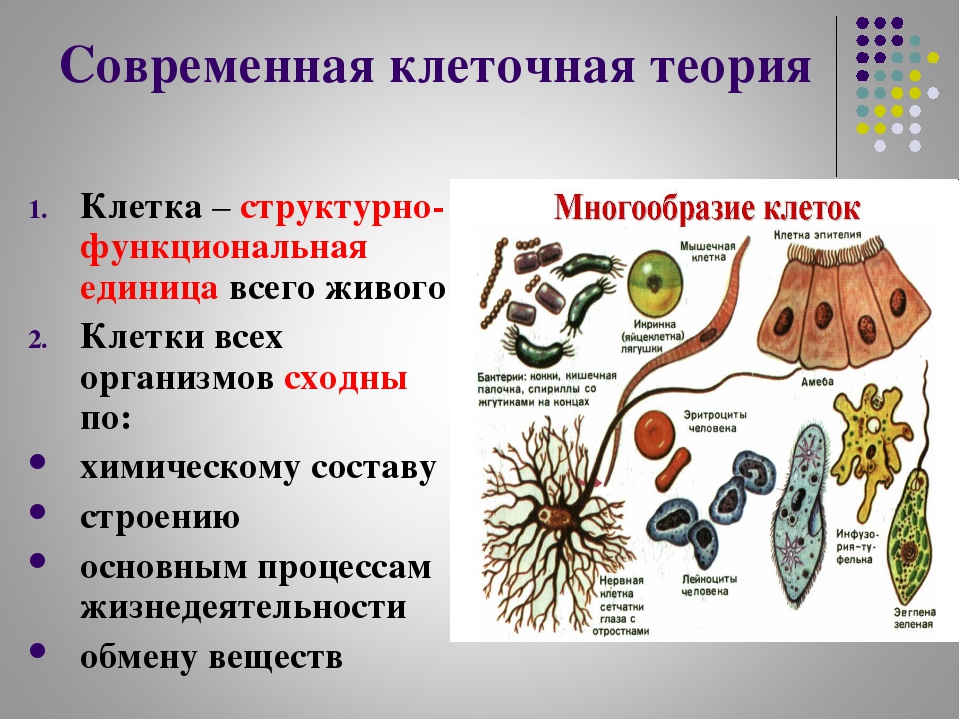
Обобщение ( Пример) по биологии за курс 7 класса 2 варианта
Биология 7 класс Вариант 2
1.Оболочка из клетчатки отсутствует в клетках
а) животных б) папоротников в) голосеменных г) покрытосеменных
2.Колониальные жгутиконосцы, имеющие форму шарика около 3 мм в диаметре, – это животные
организмы, называемые
а) инфузориями б) эвгленами в) амёбами г) вольвоксами
3.Животных, имеющих внутренний скелет из хрящевой или костной ткани, объединяют в единую
систематическую группу
а) класс Земноводные б) тип Хордовые в) отряд Бесхвостые г) семейство Лягушки
4.К типу моллюсков относят
А) речного рака б) ланцетника в) большого прудовика г) нереиду
5.Представитель какого класса типа Членистоногие изображён на рисунке?
а) многоножки б) паукообразные
в) ракообразные г) насекомые
6.Особенность покровов земноводных,
а) голая кожа, испаряющая много влаги б) влажная кожа, покрытая костной чешуёй
в) сухая кожа без желёз, с роговыми чешуями или щитками г) кожа с потовыми и сальными железами
7.Промежуточное положение между беспозвоночными и позвоночными занимают
а) ланцетник б) акулы в) скаты г) гаттерии
8.Кожное и лёгочное дыхание характерно для
а) ящериц б) крокодилов в) лягушек г) змей
9.Какой признак среди перечисленных отличает животных класса Пресмыкающиеся от животных класса
Млекопитающие?
а) использование для обитания наземно-воздушной среды б) непостоянная температура тела
в) замкнутая кровеносная система г) развитие без превращения
10.Формы проявления высшей нервной деятельности у некоторых видов птиц и млекопитающих – это
а) условные рефлексы б) мышление и разборчивая речь
в) безусловные рефлексы г) элементы рассудочного поведения
11. У млекопитающих газообмен происходит в
У млекопитающих газообмен происходит в
а) трахеях б) лёгочных пузырьках в) бронхиолах г) бронхах
12.В связи с развитием лёгочного дыхания у земноводных произошло формирование
а) трёхкамерного сердца б) грудной клетки в) ротового аппарата г)кожных желёз
13.Одновременно с широким распространением цветковых растений в эволюции животного мира
произошёл расцвет
а) насекомых б) земноводных в) пресмыкающихся г) птиц
14.Беспозвоночных животных, у которых в процессе эволюции впервые появилась кровеносная система,
относят к типу
а) плоских червей б) круглых червей в) кольчатых червей г) моллюсков
15.Выберите из перечня три элемента верного ответа.Какие позвоночные животные, встречающиеся в
водоёмах, являются земноводными?
а) обыкновенная плотва б) прудовая лягушка в) обыкновенный уж г) серая жаба д)обыкновенный тритон
16.Установите верную последовательность этапов цикла развития печёночного сосальщика, начиная с
яйца.
а) личинки прикрепляются к водным растениям и покрываются оболочкой (образуют цисты)
б) оплодотворённые яйца печёночного сосальщика попадают в воду
в) цисты проглатываются животным (или человеком) и попадают в его кишечник, а затем в печень
г) из яиц выходят микроскопические личинки, которые внедряются в тела улиток, размножаются в них и выходят в воду
Биология 7 класс Вариант 1
1.Какой буквой на рисунке, изображающем строение клетки животного, обозначена её оболочка?
а) А б) Б в) В г) Г
2.Среди обитающих в морях одноклеточных животных есть организмы, которые обладают наружным
скелетом, то есть
а) радиолярией б) фораминиферой в) раковиной г) цистой
3. Животные родственных отрядов объединяют в
Животные родственных отрядов объединяют в
а) типы б) классы в) роды г) семейства
4.Среди перечисленных групп беспозвоночных животных нервную систему диффузного типа имеют
а) Плоские черви б) Кишечнополостные в) Кольчатые черви г) Круглые черви
5.У паука, в отличие от клеща,
а) все членики тела срастаются между собой б) усики отсутствуют в) восемь ног
г) тело разделено на головогрудь и брюшко
6.У рыб клетки тела снабжаются кровью
а) смешанной б) насыщенной углекислым газом в) артериальной г) венозной
7.Жаберные щели, прикрытые жаберными крышками, имеют
а) ракообразные б) пресмыкающиеся в) костные рыбы г) хрящевые рыбы
8.Позвоночные, имеющие трёхкамерное сердце и голую кожу, выделяющую слизь, относятся к классу
а) рыб б) млекопитающих в) земноводных г) пресмыкающихся
9.У пресмыкающихся, в отличие от других позвоночных животных, кожа
а) сухая, покрыта роговыми чешуями или щитками б) служит дополнительным органом дыхания
в) влажная, покрыта костными чешуями г) влажная, богатая железами
10.Птицы отличаются от пресмыкающихся наличием
а) центральной нервной системы б) постоянной температуры тела
в) внутреннего оплодотворения г) двух кругов кровообращения
11.К млекопитающим относится
а) крокодил б) акула в) ихтиозавр г) кит
12.Древние пресмыкающиеся смогли полностью перейти к наземному образу жизни, так как у них
а) появились расчленённые конечности б) клетки тела снабжались смешанной кровью
в) появился костный скелет г) размножение происходило на суше
большинством пресмыкающихся?
а) четырёхкамерное сердце б) второй круг кровообращения
в) система кровообращения замкнутого типа г) разветвлённая сеть капилляров
14. Современные млекопитающие произошли от
Современные млекопитающие произошли от
а) динозавров б) древних птиц в) стегоцефалов г) зверозубых ящеров
15.Выберите из перечня три элемента верного ответа. Какие позвоночные животные имеют личиночную
стадию головастика?
а) тритон б) черепаха в) жаба г) карась д) лягушка
16.Установите соответствие между признаком животного и типом, для которого этот признак характерен: для каждой позиции из первого столбца подберите соответствующую позицию из второго, обозначенную цифрой.
ПРИЗНАК ЖИВОТНОГО
ТИП ЖИВОТНОГО
1)
Кольчатые черви
2)
Плоские черви
А)
Кровеносная система отсутствует.
Б)
Кровеносная система имеется.
В)
Играют роль в процессе почвообразования.
Г)
У многих видов пищеварительная система отсутствует.
Урок 6. теоретические методы исследования — Естествознание — 10 класс
Урок 6 Теоретические методы исследования
Перечень вопросов, рассматриваемых в теме:
- Что такое метод?
- Чем различаются анализ и синтез, индукция и дедукция?
- Какова взаимосвязь методов научного познания в реальной исследовательской деятельности?
Глоссарий по теме:
Метод — способ построения знания, форма практического и теоретического освоения действительности.
Анализ – разделение целостного предмета на составные части (стороны, признаки, свойства или отношения) с целью их всестороннего изучения;
Аналогия – прием познания, при котором на основе сходных объектов в одних признаках заключают об их сходстве и в других признаках;
Дедукция – логическое умозаключение от общего к частному.
Индукция – это логическое умозаключение от частных, единичных случаев к общему выводу, от отдельных фактов к обобщению.
Классификация – это разделение всех изучаемых предметов на отдельные группы в соответствии с каким – либо важным для исследователя признаком
Моделирование – изучение объекта (оригинала) путем создания и исследования его копии (модели), замещающей оригинал с определенных сторон, интересующих исследователя;
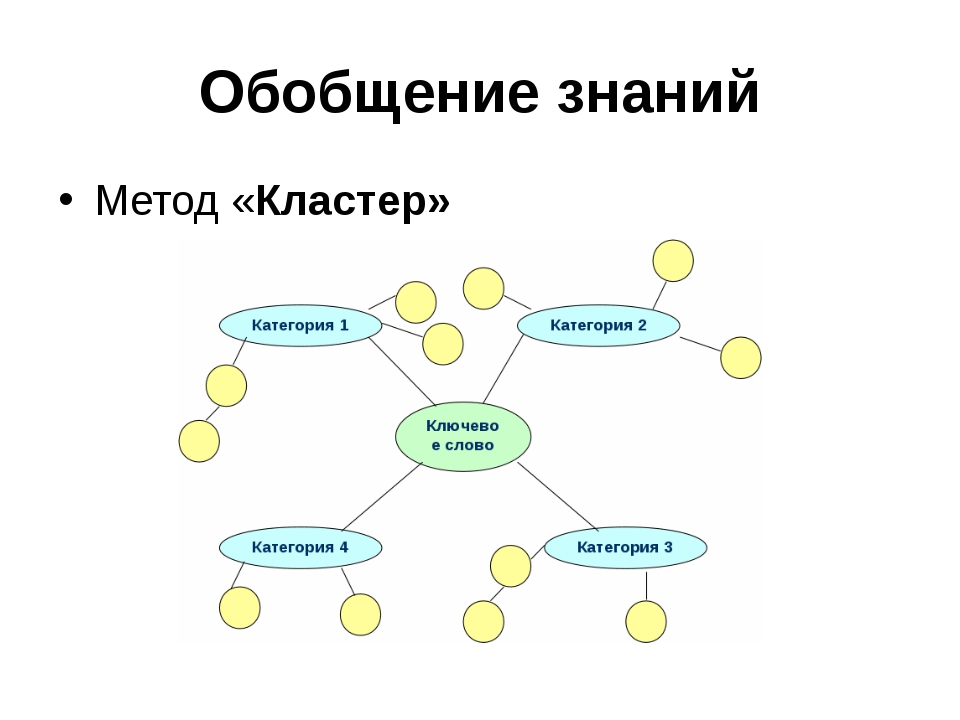
Обобщение – прием мышления, в результате которого устанавливаются общие свойства и признаки объектов;
Синтез – соединение ранее выделенных частей предмета в единое целое;
Систематизация — упорядочивание знания, т.е. приводит в систему наблюдения и экспериментальные факты.
Сравнение – методы измерений, в которых измеряемую величину сравнивают с величиной, воспроизводимой мерой.
Основная и дополнительная литература по теме урока:
- Естествознание. 10 класс [Текст]: учебник для общеобразоват. организаций: базовый уровень / И.Ю. Алексашина, К.В. Галактионов, И.С. Дмитриев, А.В. Ляпцев и др. / под ред. И.Ю. Алексашиной. – 3-е изд., испр. – М.: Просвещение, 2017. с 24 — 25.
Теоретический материал для самостоятельного изучения
Развитие научных представлений о мире шло сложным путем накопления фактов, наблюдений, открытий законов природы. Представление о мире, степень воздействия науки на жизнь человеческого общества время от времени резко менялись. Менялись и методы познания природных явлений. Роль методов как составной части науки сложно переоценить. Зачастую именно появление нового метода исследования определяет дальнейшее развитие науки. Метод в переводе с греческого языка означает «путь исследования, способ познания».
Немецкий ученый Готлиб Лейбниц (1646 — 1716) говорил: «На свете есть вещи поважнее самых прекрасных открытий – это знание метода, которым они были сделаны», а английский философ и ученый Фрэнсис Бэкон (1561 — 1626) сравнивал метод со светильником, освещающим путнику дорогу в темноте: «Даже хромой, идущий по дороге, опережает того, кто идет без дороги». Правильно подобранный метод ведет к определенной цели, дает достоверные результаты, обеспечивает получение объективных знаний.
Правильно подобранный метод ведет к определенной цели, дает достоверные результаты, обеспечивает получение объективных знаний.
Любая отрасль естествознания развивалась и развивается в два этапа. Сначала научные исследования имеют описательный характер. Происходит накопление фактических данных, составляющих основу конкретной науки. Особую ценность представляют те результаты, которые связывают воедино, приводят в систему ранее известные, но разрозненные факты. Такой период развития науки называют эмпирическим.
Эмпиризм (греч. empeiria – опыт) – учение, признающее опыт, основанный на ощущениях, единственным источником знаний. На данном этапе развития исследований естествоиспытатели ставят вопрос «как»: как протекает реакция, какое вещество получается, какие вещества необходимы и т. д.
На втором этапе развития исследования направляются на выяснение причинной связи, например, установления связи между строением вещества и его свойствами. Этот период развития науки называют рациональным. Рационализм (лат. rationalis – разумный, обоснованный, целесообразный) – направление в теории познания, признающее разум решающим источником знания. На данном этапе исследователи пытаются ответить на вопрос «почему».
Развитие в два этапа – эмпирический и теоретический (рациональный) – касается как науки в целом, так и отдельных ее разделов. Таким образом, выделяются эмпирические и теоретические методы.
Эмпирические методы: наблюдение, измерение эксперимент. Теоретические методы: анализ, синтез, абстрагирование, обобщение, индукция, дедукция, моделирование, классификация и др.
В реальном научном познании эти методы используют всегда в единстве. Например, при разработке эксперимента требуется предварительное теоретическое осмысление проблемы, формулирование гипотезы, а после проведения эксперимента — обработка результатов с использованием математических методов.
Рассмотрим особенности некоторых теоретических методов
Классификация и систематизация – это важнейшие функции любой науки.
Классификация – это разделение всех изучаемых предметов на отдельные группы в соответствии с каким – либо важным для исследователя признаком (особенно часто используется в описательных науках – многих разделах биологии, геологии, географии, кристаллографии и. т. п.). Примерами классификаций могут служить: 1) планеты Солнечной системы; 2) растения; 3) химических элементов.
К основным принципам классификации можно отнести следующие положения:
1. Все члены деления в совокупности должны составлять исходное понятие;
2. Разбиение на группы (классы) проводят по одному или небольшому числу признаков (основанию классификации), которое имеет существенное значение.
3. Члены деления не должны пересекаться, т.е. взаимно исключать друг друга.
4. Распределение на классы должно быть непрерывным, без скачков
Систематизация упорядочивает знания, т.е. приводит в систему наблюдения и экспериментальные факты. Для узкого круга фактов, например для веществ, она обычно сводится к классификации, то есть к распределению на классы. Таким образом, классификация – это тоже упорядочение, но более узкое и менее глубокое, чем систематизация.
Классификацию проводят по одному или небольшому числу признаков. Например, все химические элементы делят на металлы и неметаллы. Систематизация – это более глубокое, чем классификация, обобщение. Она отражает внутреннюю сущность объектов исследования. Например, Д. И. Менделеев систематизировал все химические элементы. В данном случае не имеет значения то, что внутренняя сущность химических элементов в то время не была ясной (электронное строение). Важно то, что суждение о внутренней связи химических элементов являлось в данной ситуации почти неизбежным следствием. Систематизация – столь широкое обобщение, что часто позволяет делать предсказания (для Периодической системы – свойства еще не открытых химических элементов), а это уже характерная черта научной теории. Таким образом, систематизация часто бывает первым шагом к научной теории.
Научные теории создаются, в основном индуктивным путем, т.е. на основе систематизации накопленной информации от частного к общему, от отдельных фактов к выводу об общей закономерности. Такой теоретический метод познания называют индукцией. Индукция как метод широко используется в науке. Можно сказать, что она является основным методом научного познания природы. Им пользуются ученые для создания и развития науки.
Например, изучение свойств отдельных химических элементов, в конце концов, привело Менделеева к созданию Периодической системы и открытию Периодического закона.
Способ рассуждения от общего к частному, то есть при котором из общих посылок с необходимостью следует заключение частного характера, называется дедукцией. Например, зная, что все селитры (нитраты щелочных и щелочноземельных металлов и нитрат аммония) хорошо растворимы в воде, мы можем сделать умозаключение, что нитрат калия KNO3 хорошо растворим в воде.
Теоретическая физика, современная математика основываются на системе аксиом, основополагающих утверждениях. Аксиомы являются фундаментом на котором строиться научного знание путем выведения умозаключений от общего к частному. Этот метод называют дедукцией. Его развивал французский философ и ученый Рене Декарт (1596-1650).
Примерам получения знания об одном предмете разными путями является открытие законов движения небесных тел. В начале XVII на основе большого количества данных наблюдений за движением планеты Марс методом индукции И. Кеплер открыл эмпирические законы движения планет в Солнечной системе. В конце этого же века на основе закона всемирного тяготения дедуктивным путем Ньютон вывел обобщенные законы движения небесных тел.
Большое значение в современной науке приобрели статистические методы, позволяющие определять средние значения, характеризующие всю совокупность изучаемых предметов. «Применяя статистический метод, мы не можем предсказать поведение отдельного индивидуума совокупности. Мы можем только предсказать вероятность того, что он будет вести себя некоторым определенным образом…
Мы можем только предсказать вероятность того, что он будет вести себя некоторым определенным образом…
Статистические законы можно применять только к большим совокупностям, но не к отдельным индивидуумам, образующим эти совокупности» (А. Эйнштейн, Л. Инфельд).
Важную роль при переходе от эмпирических исследований к созданию теории играет интуиция. Интуитивное прозрение обычно является результатом напряженной и обширной предшествующей работой мысли. Интуиция – это предчувствие, постижение истины без логического обоснования. Можно сказать, что интуиция – это высшая степень познания, когда подсознательно используется знание законов, правил, теорий, хотя между ними и есть логические пробелы. Высокое мастерство исследователя проявляется в умении заставить мозг работать на уровне интуиции.
Эйнштейн писал: “В моей жизни взгляд на мир глазами художника играл большую роль. В конце концов работа научного исследователя развивается на почве воображения. Как артист создает свои образы отчасти интуитивно, так и ученый должен обладать большой долей интуиции”.
Эту позицию разделяет и выдающейся физик XX века Макс Борна (1882-1970): “…мы, ученые, всегда должны помнить, что весь опыт базируется на чувствах. Теоретик, погрязший в своих формулах, забывший о явлениях, которые он собирался объяснить, – это уже не настоящий ученый – физик или химик; а если своими книгами он загораживается от красоты и разнообразия природы, то для меня он жалкий глупец. Ныне мы достигли разумного равновесия между экспериментом и теорией, между чувственной и интеллектуальной реальностью. И мы должны следить за тем, чтобы такое равновесие сохранилось”.
Выводы:
Методы – неотъемлемая составная часть науки, они развиваются вместе с наукой и во многом определяют ее развитие.
В реальной исследовательской деятельности методы научных исследований взаимосвязаны
Примеры и разбор решения заданий тренировочного модуля:
Задание 1. Метод, связанный с целенаправленным созданием ситуации, которая помогает изучить свойства и явления живой природы:
- естественный
- эмпирический
- теоретический
- исторический
Ответ: эмпирический
Задание 2. Начинающий исследователь целый месяц кормил одну группу из 10 крыс йогуртом, и все они прибавили в весе. Выделите цветом выводы, которые НЕ требуют дальнейшего экспериментального подтверждения?
Начинающий исследователь целый месяц кормил одну группу из 10 крыс йогуртом, и все они прибавили в весе. Выделите цветом выводы, которые НЕ требуют дальнейшего экспериментального подтверждения?
- Йогурт – это лучшее питание для крыс
- Некоторые крысы прибавляют в весе при питании йогуртом
- Йогурт не смертелен для данных животных
- Йогурт содержит все необходимые для крыс минеральные вещества и витамины
Ответ:
- Некоторые крысы прибавляют в весе при питании йогуртом
- Йогурт не смертелен для данных животных
Урок биологии 5 класс «Обобщение и систематизация знаний по теме Биология
Подготовила учитель биологии Юзвак Т. Д.
МОУСОШ № 9 им. Кирилла и Мефодия г. Каменки
План-конспект урока по биологии (5 класс). Урок обобщения и систематизации знаний по теме «Биология – наука о живом мире».
Тип урока: урок развивающего контроля.
Используемые технологии: здоровьесбережения, проблемного обучения, развивающего обучения.
Формируемые УУД: к. – самостоятельно организовывать учебное взаимодействие в группе; р. – составлять план решения проблемы, развивать умения применять полученные знания при выполнении самостоятельной работы; п. – выбирать наиболее эффективные способы решения задач в зависимости от конкретных условий; л. – формировать научное мировоззрение и мотивацию к дальнейшему изучению биологии, в. – воспитывать умение работать в заданном темпе и не теряться при выполнении самостоятельной работы.
Планируемые результаты: научиться оценивать свои результаты.
Оборудование: учебник, комплект карточек для задания «Потерянные буквы», комплект карточек для задания «Уберите лишнее», магниты, красные и зеленые фишки из цветной бумаги, ручки, белые листы бумаги, карточки с вопросами для самостоятельной работы по вариантам.
Ход урока.
Организационный момент. (2 мин.)
Учитель сообщает тему и задачи урока, объясняет роль фишек. (За правильный ответ учитель дает ответившему ученику зеленую фишку, за неполный ответ учитель дает красную фишку; затем ученики прикалывают заработанные фишки к листку с самостоятельной работой. Эти фишки дают возможность повысить балл за самостоятельную работу).
Мотивация к деятельности. Разминка (7 мин.)
Задание «Потерянные буквы». Учитель вывешивает на доску карточки со словами с пропущенными согласными буквами. Ученики должны отгадать слово. За каждый верный ответ учитель дает фишку: зеленую, если ответ верный или красную, если ответ не полный.
е – а – а (мембрана)
я – о (ядро)
и – о – а – а (цитоплазма)
у – а (лупа)
а – у – о (вакуоль)
Закрепление знаний. (Фронтальная работа)
Задание № 1. (7 мин.)
Учитель заранее на обратной стороне доски вывешивает карточки на магнитах. Из данных карточек необходимо убрать лишние:
Питание, обмен веществ, цвет, дыхание, движение, рост, объем, развитие, длина, раздражительность, размножение, ширина.
(лишние: цвет, объем, длина, ширина)
Учитель: Почему вы их убрали? Что осталось? (ответ учеников: остались признаки живых организмов)
Докажите, что это признаки живых организмов. (ответы учеников)
Ученикам, участвующим в этом задании, учитель дает фишки: зеленую, если ответ верный и красную, если ответ не полный.
Задание № 2. (7 мин.)
На каждой парте лежит лист с написанными словами. Их нужно разделить на две группы и объяснить, почему именно так разделили. Работа в паре.
Их нужно разделить на две группы и объяснить, почему именно так разделили. Работа в паре.
Объектив, штатив, сравнение, тубус, описание, предметный столик, измерение, окуляр, моделирование, эксперимент.
Ответы учеников: первая группа слов – методы изучения природы, вторая группа слов – основные части микроскопа.
За это задание учитель также дает фишки.
Физминутка. (3 мин.)
Упражнения для осанки: и.п. – стоя. Резко поднять плечи вверх, плавно отвести назад и плавно опустить, зафиксировать в таком положении на 3 секунды. Повторить 5 раз.
Дыхательное упражнение «Цветочный магазин»: и.п. – стоя. Закрыть глаза и представить, что вы находитесь в цветочном магазине. Вдыхайте медленно аромат цветов и медленно выдыхайте. Повторить 5 раз.
Самостоятельная работа. (15 мин.)
Учитель раздает каждому ученику лист с вариантом работы.
Вопросы:
1 вариант:
Чем живая природа отличается от неживой?
Какие методы изучения природы вы знаете?
Нарисуйте стадии деления клетки.
2 вариант:
Перечислите основные признаки живого организма.
Какие основные части микроскопа вы знаете?
Нарисуйте растительную клетку и подпишите основные части.
Рефлексия. (4 мин.)
Почему биологические знания нужны каждому образованному человеку?
Почему хозяйственная деятельность человека часто приводит к гибели живых организмов?
Почему жизнь живых организмов неразрывно связана неживой природой?
|
Самостоятельная работа «Вегетативные органы покрытосеменных растений» Вариант 1
Лист – это…. Почка – это….
Укажите правильную последовательность движения воды в растении, начиная от корня: а) сосуды древесины стебля б) жилки листа в) зона проведения г) устьица д) корневые волоски
|
Самостоятельная работа «Вегетативные органы покрытосеменных растений» Вариант 2
Стебель – это…
Побег – это….
а) газообмен; б) транспорт минеральных веществ; в) транспорт органических веществ г) выделение тепла.
Укажите последовательность процессов, происходящих при фотосинтезе: а) выделение кислорода б) образование сахаров в) освещение листьев солнцем г) разложение воды солнцем д) поглощение воды е) поглощение углекислого газа 4. Подпишите внутреннее строение стебля под № 1, 9, 3, 4, 5:
5. Раскройте биологическую роль листа в жизни растения. |

| 1 | 1 | Биологические термины и понятия | ||
| 2 | 1 | Биология как наука. Методы научного познания. Уровни организации живого | ||
| 3 | 1 | Генетическая информация в клетке. Хромосомный набор, соматические и половые клетки Хромосомный набор, соматические и половые клетки | ||
| 4 | 2 | Клетка как биологическая система. Жизненный цикл клетки | ||
| 5 | 2 | Клетка как биологическая система. Строение клетки, метаболизм. Жизненный цикл клетки | ||
| 6 | 1 | Моно- и дигибридное, анализирующее скрещивание | ||
| 7 | 2 | Организм как биологическая система. Селекция. Биотехнология | ||
| 8 | 2 | Организм как биологическая система. Селекция. Биотехнология | ||
| 9 | 2 | Многообразие организмов. Бактерии, грибы, растения, животные, вирусы | ||
| 10 | 2 | Многообразие организмов. Бактерии, грибы, растения, животные, вирусы | ||
| 11 | 2 | Многообразие организмов. Основные систематические категории, их соподчинённость | ||
| 12 | 2 | Организм человека. Гигиена человека | ||
| 13 | 2 | Организм человека | ||
| 14 | 2 | Организм человека | ||
| 15 | 2 | Эволюция живой природы | ||
| 16 | 2 | Эволюция живой природы. Происхождение человека | ||
| 17 | 2 | Экосистемы и присущие им закономерности. Биосфера | ||
| 18 | 2 | Экосистемы и присущие им закономерности. Биосфера | ||
| 19 | 2 | Общебиологические закономерности | ||
| 20 | 2 | Общебиологические закономерности. Человек и его здоровье | ||
| 21 | 2 | Биологические системы и их закономерности |
Комитет 1540 — О Комитете
Основные сведения
Резолюция 1540 (2004) Совета Безопасности Организации Объединенных Наций
28 апреля 2004 года Совет Безопасности Организации Объединенных Наций в соответствии с главой VII Устава Организации Объединенных Наций единогласно принял резолюцию 1540 (2004), в которой подтверждено, что распространение ядерного, химического и биологического оружия, а также средств его доставки представляет угрозу для международного мира и безопасности. В соответствии с резолюцией все государства обязаны, в частности, воздерживаться от оказания в любой форме поддержки негосударственным субъектам, которые пытаются разрабатывать, приобретать, производить, обладать, перевозить, передавать или применять ядерное, химическое или биологическое оружие и средства его доставки.
В соответствии с резолюцией все государства обязаны, в частности, воздерживаться от оказания в любой форме поддержки негосударственным субъектам, которые пытаются разрабатывать, приобретать, производить, обладать, перевозить, передавать или применять ядерное, химическое или биологическое оружие и средства его доставки.
Резолюция 1540 (2004) налагает на все государства обязательства в отношении принятия законодательства в целях предотвращения распространения ядерного, химического и биологического оружия и средств его доставки и предусматривает установление надлежащего национального контроля над относящимися к ним материалами в целях предотвращения их незаконного оборота. В ней также содержится призыв к расширению масштабов международного сотрудничества в контексте таких усилий. В резолюции заявлено о поддержке многосторонних договоров, нацеленных на ликвидацию или предотвращение распространения оружия массового уничтожения и подтверждается важность того, чтобы все государства этих договоров в полной мере выполняли их; в ней подтверждено, что никакие обязательства по резолюции 1540 (2004) не должны толковаться как противоречащие или изменяющие права и обязательства государств — участников Договора о нераспространении ядерного оружия, Конвенции о запрещении химического оружия и Конвенции о запрещении биологического и токсинного оружия, или меняющие сферу ответственности МАГАТЭ и ОЗХО.
27 апреля 2006 года Совет Безопасности продлил срок действия мандата Комитета 1540 на двухлетний период в резолюции 1673 (2006), в которой были подтверждены цели резолюции 1540 (2004) и говорилось о том, что Совет Безопасности заинтересован в активизации усилий Комитета по содействию полному выполнению этой резолюции.
25 апреля 2008 года Совет Безопасности принял резолюцию 1810 (2008), в которой он продлил мандат Комитета 1540 на трехлетний период, с дальнейшей поддержкой экспертов, до 25 апреля 2011 года. В резолюции 1810 (2008) Совет Безопасности настоятельно призвал Комитет 1540 продолжать укреплять его роль в содействии оказанию технической помощи, в том числе путем активного участия в подборе предложений и просьб об оказании помощи, тем самым повышая действенность и доступность такой помощи. Совет Безопасности также просил Комитет 1540 рассмотреть вопрос о всеобъемлющем обзоре хода осуществления резолюции 1540 (2004). В рамках этого всеобъемлющего обзора Комитет 1540 постановил провести открытое совещание с широким числом участников от государств — членов Организации Объединенных Наций и соответствующих международных организаций. Открытое совещание состоялось в Центральных учреждениях Организации Объединенных Наций в период с 30 сентября по 2 октября 2009 года, и его заключительный документ был опубликован на веб-сайте Комитета 1540.
Совет Безопасности также просил Комитет 1540 рассмотреть вопрос о всеобъемлющем обзоре хода осуществления резолюции 1540 (2004). В рамках этого всеобъемлющего обзора Комитет 1540 постановил провести открытое совещание с широким числом участников от государств — членов Организации Объединенных Наций и соответствующих международных организаций. Открытое совещание состоялось в Центральных учреждениях Организации Объединенных Наций в период с 30 сентября по 2 октября 2009 года, и его заключительный документ был опубликован на веб-сайте Комитета 1540.
20 апреля 2011 года Совет Безопасности принял резолюцию 1977 (2011), в которой он вновь подтвердил, что распространение ядерного, химического и биологического оружия, а также средств его доставки представляет собой угрозу для международного мира и безопасности, и продлил мандат Комитета 1540 на 10 лет — до 2021 года. Таким образом Совет Безопасности признал, что полное осуществление резолюции 1540 (2004) всеми государствами является долгосрочной задачей, которая потребует постоянных усилий на национальном, региональном и международном уровнях. В резолюции 1977 (2011) также предусмотрено проведение двух всеобъемлющих обзоров, одного — по истечении пяти лет и одного — до продления действия мандата. Кроме этого, Комитет 1540 в соответствии с резолюцией 1977 (2011) должен продолжать укреплять свою роль в содействии оказанию технической помощи и активизировать сотрудничество с соответствующими международными организациями. Комитет также должен продолжать совершенствовать его информационно-просветительские усилия и продолжать принимать меры по обеспечению транспарентности. 29 июня 2012 года Совет Безопасности принял резолюцию 2055 (2012), которая расширила состав группы экспертов в поддержку работы Комитета 1540 до девяти (9) экспертов.
15 декабря 2016 года, после представления Совету Безопасности доклада о всеобъемлющем обзоре, проведенном в 2016 году, была единогласно принята резолюция 2325 (2016). В этой резолюции были подтверждены обязанности, зафиксированные в резолюции 1540 (2004). В указанной резолюции, в частности, ко всем государствам был обращен призыв активизировать свои усилия по достижению полного осуществления резолюции 1540 (2004), уделяя особое внимание, когда и где это уместно, тем областям, в которых необходимо принять меры и повысить их эффективность.
В этой резолюции были подтверждены обязанности, зафиксированные в резолюции 1540 (2004). В указанной резолюции, в частности, ко всем государствам был обращен призыв активизировать свои усилия по достижению полного осуществления резолюции 1540 (2004), уделяя особое внимание, когда и где это уместно, тем областям, в которых необходимо принять меры и повысить их эффективность.
Следующий всеобъемлющий обзор должен быть проведен к 25 апреля 2021 года, когда истекает срок действия мандата Комитета 1540.
Обобщение передового педагогического опыта — биология, прочее
Развитие творческих способностей учащихся при изучении биологии и химии.
Р. Л. Фостенко,
учитель биологи и химии
МКОУ ООШ №14 г.Коркино,
ст. Дубровка, Челябинская область
Одним из эффективных способов развития креативного мышления у школьников может служить совершенствование умения первичной обработки учебной информации, способствующее активизации мыслительных процессов, повышению скорости и улучшению качества усвоения теоретического материала. В этом направлении большие возможности дает работа с учебным текстом.
Как показывают психологические исследования творческую личность отличает особое сочетание личностно-деловых качеств, т. е. способность создавать оригинальные ценности, принимать нестандартные решения. Способствовать развитию творческой личности мы можем, совершенствуя умения ученика:
1) видеть проблему;
2) выдвигать гипотезы, подлежащие проверке, и предлагать способы этой проверки;
3) намечать план решения этой проблемы, привлекая, имеющиеся знания, и объяснять свои решения;
4) создавать положительную систему нравственных ценностей.
Практика показывает, что многие дети, особенно учащиеся 5-6 классов, не умеют бегло и осмысленно читать учебные тексты, поэтому очень важно уделять внимание формированию умений и навыков самостоятельной работы с учебной литературой:
— выделять в тексте главную мысль;
— объяснять, почему текст так называется;
— кратко излагать текст;
— составлять список ключевых слов;
— конспектировать;
— ставить к тексту простые вопросы;
— составлять план параграфа;
— выстраивать пирамиды понятий.
Применение на уроках этих форм работы особенно актуально для тех учеников, которые не выполняют домашние задания. Объясняется это тем, что проработка текстов осуществляется по принципу «малого погружения»: на уроке ставится задача получить знания и умения, определенные стандартом образования, а домашнее задание призвано развивать полученные на уроке знания.
Исследования психологов показывают, что на 1-м (пассивном ) этапе человек запоминает лишь 10% от прочитанного, 20% от услышанного, 30% от увиденного, на 2 (активном) этапе – 50% от услышанного и увиденного, 70% от сказанного им самим, 90% от сделанного им самим.
Таким образом, чем более самостоятельным будет ребенок на уроке, тем выше будет уровень реализованной креативности. Для развития творческого мышления необходимо обучить, а затем и натренировать ребенка на применение мыслительных операций при изучении текста или термина:
анализ: мысленное разделение предметов на части или свойства;
синтез: мысленное объединение частей или свойств предметов и явлений в единое целое;
сравнение: сопоставление предметов и явлений, нахождение сходства и различий между ними;
обобщение: мысленное объединение предметов и явлений по общим признакам;
абстрагирование: выделение одних признаков и отвлечение от других (осуществляется на основе анализа).
Практика показала, что наиболее успешными являются следующие методы обработки учебной информации.
1.»Пирамида понятий».
Изучаемое понятие записывают посредине листа, под ним располагаются понятия, которые «поглощают» центральный термин, под ним – понятия, входящие в его состав. Получается пирамида с узкой вершиной и широким основанием. Упражнение помогает выявить взаимосвязи между терминами, включенными в пирамиду, и требуют от учащихся понимания всех используемых понятий.
Пример
Ткань (клетки + межклеточное вещество)
↑
Клетка
↓
Цитоплазма, ядро
↓
Комплекс Гольджи, рибосомы, митохондрии, лизосомы
2. «Понятийно-терминологическая карта».
«Понятийно-терминологическая карта».
Представляет собой карту с терминами, используя которую нужно составить текст любого жанра –статья, рассказ о каком-либо организме. Или, наоборот, используя текст параграфа, составить понятийно-терминологическую карту. Такую работу можно выполнять в конце изучения темы, раздела. Она очень полезна при работе в группе: одна группа работает с первым столбцом – составляет рассказ по терминам; другая использует материал лишь второго столбца, третья приводит примеры (последний столбец). В итоге весь материал прорабатывается.
Пример
Понятие | Сущность | Подробно |
1.Ткань | Группа клеток с окружающим их межклеточным веществом, объединенные общим происхождением, схожим строением и одинаковой функцией | В организме человека 4 вида тканей: эпителиальная, соединительная, нервная, мышечная, ткани внутренней среды организма. |
3. «Мысль по кругу».
Может применяться для отработки терминов. На парту кладется небольшой текст и несколько утверждений, среди которых есть как правильные, так и неправильные.
После небольшой подготовки первый ученик отвечает – верно ли первое в списке утверждение, затем второй соглашается или нет с этим утверждением и т. д. Когда обозначено отношение к последнему утверждению, следующий ученик возвращается к первому тезису – возможно его мнение отличается от предыдущего, и так, пока последний ребенок в классе не выскажется по данной теме Все результаты опроса записываются на доске.
4. «Дидактические игры со словами».
Это короткие, но емкие и информативные задания и упражнения, расширяющие общий кругозор. Они позволяют заинтересовать учащихся и заставить размышлять, поэтому удобны для отработки ключевых понятий и терминов.
Примеры игр
Составить разные предложения из набора ключевых слов.

Назвать как можно больше значений одного и того же термина (например, «клетка», «сосуд».
Составить из букв длинного слова как можно больше других слов.
Классифицировать термины и ключевые слова.
Составить краткий рассказ из слов на одну букву, сочетающихся по смыслу с предложенным текстом.
Восстановить деформированный текст.
Расставить термины и ударения.
Выразить мысль другими словами.

5. «Игра шифровка» — для передачи сообщения два преступника выдумали шрифт. Полиции удалось перехватить одно из сообщений. Помогите узнать, какую информацию зашифровали преступники.
Пример
Хищная корова хоботом и ластами грызет насекомых, а рукокрылая лошадь помогает обезьяне собирать яйца в сумку.
(Перечислены отряды Хищные, Парнокопытные, Хоботные, Ластоногие, Грызуны, Насекомоядные, Рукокрылые, Непарнокопытные, Приматы; Подклассы Яйцекладущие и Сумчатые).
6. Игра «Гусеница» — слова, относящиеся к какому-либо понятию, записываются слитно; необходимо их вычленить и записать в тетрадь.
Пример
Найти названия птиц:
(Соколибрибисойканарейказаркайральбатросоедроздрофазанандудодубоносорокакаду.
(Сокол, колибри, ибис, сойка, канарейка, казарка, кайра, альбатрос, осоед, дрозд, дрофа, фазан, нанду, удод, дубонос, сорока, какаду.)
На уроках химии, при изучении нового материала, использую учебные задачи, которые начинаются с практико- ориентированного проблемного вопроса «почему?». Проблемно- творческие интегрированные задания по химии снабжены научно-популярно информацией. С их помощью реализуется компетентностный подход к творческому саморазвитию личности в процессе обучения.
Задача 1. Почему при открывании бутылки с лимонадом происходит бурное выделение газа?
Научно-популярная информация- подсказка. Растворимость газов увеличивается с увеличением давления. Углекислый газ растворяется в лимонаде под давлением, поэтому при открывании бутылки избыточный газ выделяется из раствора.
Углекислый газ растворяется в лимонаде под давлением, поэтому при открывании бутылки избыточный газ выделяется из раствора.
Задание. Составьте уравнение химической реакции, происходящей при открывании бутылки с газированной водой.
(Ответ. Н2СО3 = СО2 + Н2О)
Творческое задание. Составьте карточку «Анкетные данные углекислого газа» (из семи пунктов: агрегатное состояние, запах, цвет и т. д.).
Образовательный продукт. Составление карточки.
Задача 2. Почему лук «без боли и печали» доводит до слез?
Научно-популярная информация-подсказка. Когда мы режем лук, происходит выделение летучего едкого вещества – тиопропиональдегид-S-оксида. Это соединение является лакриматором (от лат. Lakrima –слеза), именно оно вызывает слезы, когда мы режем сырой репчатый лук.
Задание. Установите молекулярную формулу тиопропиональдегид-S-оксида, если массовые доли элементов в нем составляют: С — 40%; Н – 6,6%; О – 17,8%; S – 35,6%; Мr = 90,14.
(Ответ. С3Н6ОS).
Творческое задание. Составить развернутый план текста параграфа «Сера» в учебнике.
Образовательный продукт. Составленный план текста учебника.
Задача 3. Почему потускневшие жемчужины давали склевать домашней птице?
Научно-популярная информационная подсказка. Известен способ реставрации потускневших жемчужин. Жемчуг на 86% состоит из карбоната кальция. Верхний потускневший или поцарапанный слой растворяемся в соляной кислоте желудочного сока петуха или другой живности, открывается блестящий слой.
Задание. Составьте уравнение химической реакции карбоната кальция с соляной кислотой.
(Ответ. СаСО3 + 2НСI = СаСI2 + Н2СО3)
Творческое задание. Отработать имитационный опыт «Обработка жемчуга соляной кислотой» и демонстрировать его перед аудиторией.
Образовательный продукт. План проведения имитационного опыта.
Задача 4.Почему зимой все время спать хочется?
Научно-популярная информация-подсказка. Холодной зимой воздух более разрежен, в нем содержится меньше кислорода, чем требуется для активности нашему организму. Кровь становится более густой, кровоток более медленным, увеличивается нагрузка на сердце и сосуды. Отсюда- усталость, головные боли, вызванные спазмами сосудов, зевота (кстати, она является характерным признаком гипоксии (недостатка кислорода) тканей мозга).
Задание. Больному с затрудненным дыханием предлагают сделать несколько вдохов из кислородной подушки. Во сколько раз примерно это облегчает его дыхание?
(Ответ. Примерно в 5 раз.)
Творческое задание. Подготовить вопросы автору учебника химии О. С. Габриеляну, которые помогли бы лучше и глубже понять смысл и значение параграфа «Неметаллы: кислород, озон, воздух. (О.С. Габриелян. 9кл. М.: Дрофа.2016г.)
Образовательный продукт. Составленные вопросы.
Сегодня важное значение в обучении приобретает индивидуальный подход, когда учебный процесс строится с учетом индивидуальных особенностей учащихся, создаются оптимальные условия для реализации потенциальных возможностей каждого ученика. Реализация принципа «обучать не всех, а каждого», приводит к активизации познавательной самостоятельности учащихся. На уроке, ограниченном по времени очень сложно этого добиться.
Средством индивидуализации обучения могут являться могут стать домашние задания, дифференцированные по уровню сложности, объему и виду творчества. Задания, которые интересны детям, чтобы их хотелось выполнить, чтобы сам процесс выполнения домашнего задания превратился в увлекательную и полезную с точки зрения самообразования ученика работу.
Заданий должно быть несколько, чтобы ученик с высоким интеллектом и высокими образовательными потребностями и ученик, не достигший успехов в обучении, смогли выбрать себе задания.
Креативные (творческие) задания
Тема «Простейшие»
1.Опишите амебу обыкновенную, используя только прилагательные. (Метод образного видения). В скобках указаны методы креативной педагогики (по А.В. Хуторскому.)
2.Представьте себе, что вы – эвглена зеленая. Опишите свои ощущения. (Метод вживания.)
Тема «Черви»
3. Найдите в тексте биологические ошибки. (Метод ошибок.)
Летом я жила в деревне. У бабушки большой огород, поэтому работы хватало всем. Мы пропалывали, поливали, выбирали из почвы дождевых червей, чтобы они не повредили корни растений. После работы мы мыли руки с мылом, ведь вместе с почвой под ногти могли попасть яйца печеночных сосальщиков – опасных паразитов человека. А однажды на бабушкину капусту напали белые планарии и стали объедать сочные нежные листья. Мы всех червей собрали и уничтожили. Летом мы поработали на славу, а осенью собрали небывалый урожай!
4. Что здесь лишнее? Почему? (Метод сравнения.)
А) печеночный сосальщик, свиной цепень, медицинская пиявка, белая планария.
Б) человеческая аскарида, свиной цепень, медицинская пиявка, нереида.
В) пескожил, белая планария, ложноконская пиявка, дождевой червь.
Тема «Насекомые»
5. Представьте, что вы – гусеница, превращающаяся в бабочку-белянку. Опишите свои ощущения. (Метод вживания.)
6.Нарисуйте зоошутку – необычное насекомое, которого не существует в природе: части тела возьмите от разных насекомых. Придумайте названия этому необычному насекомому. (Метод фактов.)
7. Как вы думаете, что произойдет, если вдруг исчезнут все насекомые? (Метод фактов.)
Тема «Рыбы. Земноводные. Пресмыкающиеся».
8.Придумайте ребусы: один о рыбах, второй о земноводных, третий о пресмыкающихся. (Метод символического видения.)
9. Составьте описание «особо опасного преступника» — змеи кобры. (Метод фактов. )
)
Тема «Млекопитающие»
10.Опишите уссурийского тигра и индийского слона, используя только прилагательные. (Метод символического видения.)
11. С детства знакомая картина: еж, несущий на своих иголках яблоко…
— Куда он его несет?
— К какому отряду относится еж?
— Чем же он питается?
— Может, он запасает яблоки на зиму?
— Как зимует еж?
— Нужно ли ему зимой пропитание?
Так нужны ли ежу яблоки? (Метод гипотез.)
Известно, что если то или иное событие, факт, слово, поступок задели самые сокровенные чувства ребенка, вызвали у него положительные или отрицательные эмоции (это также важно и даже необходимо, ибо все познается в сравнении), то запоминается надолго, возможно, навсегда. К том уже на таких уроках не развивается утомление, они способствуют раскрытию творческих способностей ребенка, помогают преодолеть однотипность уроков.
Ш. А. Амонашвили говорил: «Надо прогнать с уроков бога сна Морфея и чаще приглашать бога смеха Момуса».
Однообразие и скука – враги творчества!
Обобщение в экологии и эволюционной биологии: от гипотезы к парадигме
Андерсен, Н. Мёллер: 1982, Полуводные насекомые (Hemiptera: Gerromorpha): филогения, адаптации, биогеография и классификация, , Scandinavian Science Press Дания.
Google ученый
Allen, T.F.H. и Старр, Т.Б .: 1982, Иерархия, Перспективы экологической сложности , University of Chicago Press, Чикаго.
Google ученый
Арнквист, Г .: 1988, «Охрана спаривания и смещение сперматозоидов в водонагревателе Gerris lateralis Schumm. (Heteroptera: Gerridae) ’, Пресноводная биология 19 , 269–274.
(Heteroptera: Gerridae) ’, Пресноводная биология 19 , 269–274.
Артикул Google ученый
Арнквист, Г .: 1992a, «Драка перед копуляцией у водного долгонога: межполовой конфликт или оценка партнера», Поведение животных 43 , 559–567.
Артикул Google ученый
Арнквист, Г .: 1992b, «Влияние рабочего соотношения полов на относительный успех спаривания экстремальных мужских фенотипов у водного долгонога Gerris odontogaster (Zett.) (Heteroptera; Gerridae)», Animal Behavior 38 , 681–683.
Артикул Google ученый
Арнквист, Г.: 1997, «Эволюция систем спаривания водолазов: причины и последствия сексуальных конфликтов», в JC Choe и BJ Crespi (ред.), , Эволюция систем спаривания у насекомых и паукообразных , Cambridge University Press, Кембридж, стр. 146–163.
Google ученый
Белл, Г .: 1982, Шедевр природы , Крум-Хелм, Лондон.
Google ученый
Боннер, Дж.Т .: 1969, The Scale of Nature , World’s Work Ltd, Кингсвуд, Великобритания.
Google ученый
Басс, Л .: 1987, Эволюция индивидуальности , Princeton University Press, Princeton, NJ.
Google ученый
Коул, Л.К .: 1954, «Популяционные последствия явлений истории жизни», Quarterly Review of Biology 29 , 103–137.
Артикул Google ученый
Eldredge, N .: 1985, Unfinished Synthesis , Oxford University Press, Нью-Йорк.
Google ученый
Greenslade, P.J.M .: 1983, «Adversity Selection and the Habitat Templet», American Naturalist 122 , 352–365.
Артикул Google ученый
Хайла, Ю.: 1988, «Многоликая экологическая теория и данные», Oikos . 53 , 408–411.
Артикул Google ученый
Хайла, Ю.: 1990, «К экологическому определению острова: взгляд на северо-запад Европы», Journal of Biogeography 17 , 561–568.
Артикул Google ученый
Хайла, Ю.и Левинс, Р .: 1992, Человечество и природа — Экология, наука и общество , Плутон Пресс, Лондон.
Google ученый
Hairston, N.G. Ср .: 1989, Экологические эксперименты, цель, дизайн и выполнение , Cambridge University Press, Кембридж.
Google ученый
Яблонски П. и Вепсяляйнен К .: 1995, «Конфликт между полами у водного долгонога, Gerris lacustris : проверка двух гипотез охранного поведения самцов», Поведенческая экология 6 , 388–392.
Артикул Google ученый
Янзен, Д.: 1977, «Что такое одуванчики и тля?», Американский натуралист 111 , 586–589.
Артикул Google ученый
Klingenberg, C.P. и Спенс, Дж. Р.: 1997, «О роли размера тела в эволюции жизненного цикла», Ecological Entomology 22 , 55–68.
Артикул Google ученый
Kuhn, T .: 1962, Структура научных революций , University of Chicago Press, Чикаго.
Google ученый
Лентон Т.М .: 1998, «Гайя и естественный отбор», Nature 394 , 439–447.
Артикул Google ученый
Лернер, И.М .: 1954, Genetic Homeostasis , Dover Publications, New York.
Google ученый
Левинс, Р . : 1966, «Стратегия построения моделей в популяционной биологии», American Scientist 54 , 421–431.
: 1966, «Стратегия построения моделей в популяционной биологии», American Scientist 54 , 421–431.
Google ученый
Левинс, Р .: 1968, Эволюция в меняющихся средах , Princeton University Press, Princeton, NJ.
Google ученый
Левинс, Р .: 1974, «Качественный анализ частично определенных систем», Анналы Нью-Йоркской академии наук 231 , 123–138.
Артикул Google ученый
Левинс Р. и Левонтин Р.: 1985, Диалектический биолог , Издательство Гарвардского университета, Кембридж, Массачусетс.
Google ученый
Лием, К.Ф .: 1990, «Ключевые эволюционные инновации, дифференциальное разнообразие и симекоморфис», в M.H. Nitecki (ed.), Evolutionary Innovations , University of Chicago Press, Чикаго, стр. 147–170.
Google ученый
Лавлок, Дж. Э .: 1979, Гайя, Новый взгляд на жизнь на Земле , Oxford Univeristy Press, Оксфорд.
Google ученый
Лавлок, Дж.E .: 1991, Healing Gaia, Практическая медицина для планеты , Harmony Books, Нью-Йорк.
Google ученый
 org/Book»>
org/Book»>Макартур Р.Х. и Уилсон Е.О .: 1967, Теория биогеографии острова , Princeton University Press, Princeton, NJ.
Google ученый
McNaughton, S.J .: 1975, ‘ r — & K -selection in Typha ’, American Naturalist 109 , 251–261.
Артикул Google ученый
Мейнард Смит, Дж .: 1978, Эволюция пола , Cambridge University Press, Кембридж.
Google ученый
Nummelin, M., Vepsäläinen, K. и Spence, JR: 1988, «Инфекция яиц Gerrid (Heteroptera: Gerridae) паразитоидом Tiphodytes gerriphagus Marchal (Hymenoptera: Scelionidae) в Финляндии», Fennici 25 , 299–302.
Google ученый
О’Нил Р.В., Де Ангелис Д.Л., Вайде Дж.Б. и Аллен Т.Х.Ф .: 1986, Иерархическая концепция экосистем , Princeton University Press, Princeton, NJ.
Google ученый
Питерс Р.Х .: 1976, «Тавтология в эволюции и экологии», Американский натуралист 110 , 1–12.
Артикул Google ученый
Пьянка, Е.Р .: 1970, «О r и K Selection», American Naturalist . 104 , 592–597.
Артикул Google ученый
Pianka, E.R .: 1972, ‘ r and K Selection or b and d Selection’, American Naturalist 106 , 581–588.
Артикул Google ученый
Поппер, К.Р .: 1965, Домыслы и опровержения: рост научных знаний , Harper Torchbooks, Нью-Йорк.
Google ученый
Пуччиа, С.Дж. и Левинс, Р .: 1986, Качественное моделирование сложных биологических систем: Введение в анализ петель и усреднение по времени , Издательство Гарвардского университета, Кембридж, Массачусетс.
Google ученый
Рофф, Д.A .: 1992, Эволюция историй жизни: теория и анализ , Чепмен и Холл, Нью-Йорк.
Google ученый
Роу, Л .: 1992, «Комфортная многомужество у водного долгонога: конфликты при поиске пищи и контроль самками частоты совокупления и продолжительности сторожевого режима», Поведение животных 44 , 189–202.
Артикул Google ученый
Роу, Л., Арнквист, Г., Сих, А. и Крупа, Дж. Дж .: 1994, «Сексуальный конфликт и эволюционная экология моделей спаривания: водные долгоножки как модельная система», Trends in Ecology and Evolution 9 , 289–293.
Артикул Google ученый
Rubenstein, D.I .: 1989, «Конкуренция спермы у водного долгонога, Gerris remigis », Animal Behavior 38 , 631–636.
Артикул Google ученый
Salthe, S.N .: 1985, Evolving Hierarchical Systems , Columbia University Press, Нью-Йорк.
Google ученый
Satchell, J.E .: 1980, « r Worms и K Worms: основа для классификации стратегий, связанных с люмбрицидными дождевыми червями», в D.L. Диндал (ред.), Биология почвы в связи с практикой землепользования.Материалы 7-го Международного коллоквиума по почвенной зоологии , EPA, Вашингтон, стр. 848–854.
Google ученый
Саволайнен, Р .: 1990, «Успех колонии покорного муравья Formica fusca на территориях доминирующего Formica polyctena », Ecological Entomology 15 , 79–85.
Артикул Google ученый
Саволайнен, Р.: 1991, «Вмешательство деревянного муравья влияет на выбор размера и скорость извлечения добычи на Formica fusca », Поведенческая экология и социобиология 28 , 1–7.
Артикул Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Саволайнен Р. и Вепсяляйнен К .: 1988, «Иерархия конкуренции среди северных муравьев: влияние на разделение ресурсов и структуру сообщества», Oikos 51 , 135–155.
Артикул Google ученый
Саволайнен, Р. и Вепсяляйнен, К .: 1989, «Нишевая дифференциация видов муравьев на территориях древесных муравьев Formica polyctena », Oikos 56 , 3–16.
Артикул Google ученый
Саволайнен, Р., Вепсяляйнен, К. и Вуоренринне, Х .: 1989, «Муравьиные сообщества в таежном биоме: проверка роли территориальных древесных муравьев», Oecologia 81 , 481–486.
Артикул Google ученый
Шмальгаузен И.И .: 1938, Организация как Целое в Индивидуальном ном и Историческом Развитии, Издательство Академии Наук СССР, Москва. (на русском языке)
Google ученый
Sih, A .: 1988, «Влияние хищников на использование местообитаний, активность и брачное поведение полуводных насекомых», Animal Behavior 36 , 1846–1848.
Артикул Google ученый
Си, А., Крупа, Дж. И Трэверс, С .: 1990, «Экспериментальное исследование влияния риска хищничества и режима кормления на брачное поведение водного долгонога», American Nat Uralist 135 , 284–290.
Артикул Google ученый
Саутвуд, T.R.E .: 1977, «Среда обитания, храм экологических стратегий?», Journal of Animal Ecology 46 , 337–365.
Артикул Google ученый
Саутвуд, T.R.E .: 1988, «Tactics, Strategies and Templets», Oikos 52 , 3–18.
Артикул Google ученый
Спенс, J.R .: 1986a, «Взаимодействие между яичным паразитоидом Scelionid Tiphodytes gerriphagus (Hymenoptera) и его хозяевами Gerrid (Heteroptera)», Canadian Journal of Zoology 64 , 2728–2738.
Артикул Google ученый
Спенс, J.R .: 1986b, «Относительное влияние факторов смертности в полевых популяциях Gerris buenoi Kirkaldy (Heteroptera: Gerridae)», Oecologia 70 , 68–76.
Артикул Google ученый
Спенс, Дж. Р.: 1989, «Хабитат Храм и стратегии жизненного цикла прудовиков (Heteroptera: Gerridae): репродуктивный потенциал, фенология и диморфизм крыльев», Canadian Journal of Zoology 67 , 2432–2447.
Артикул Google ученый
Спенс, Дж.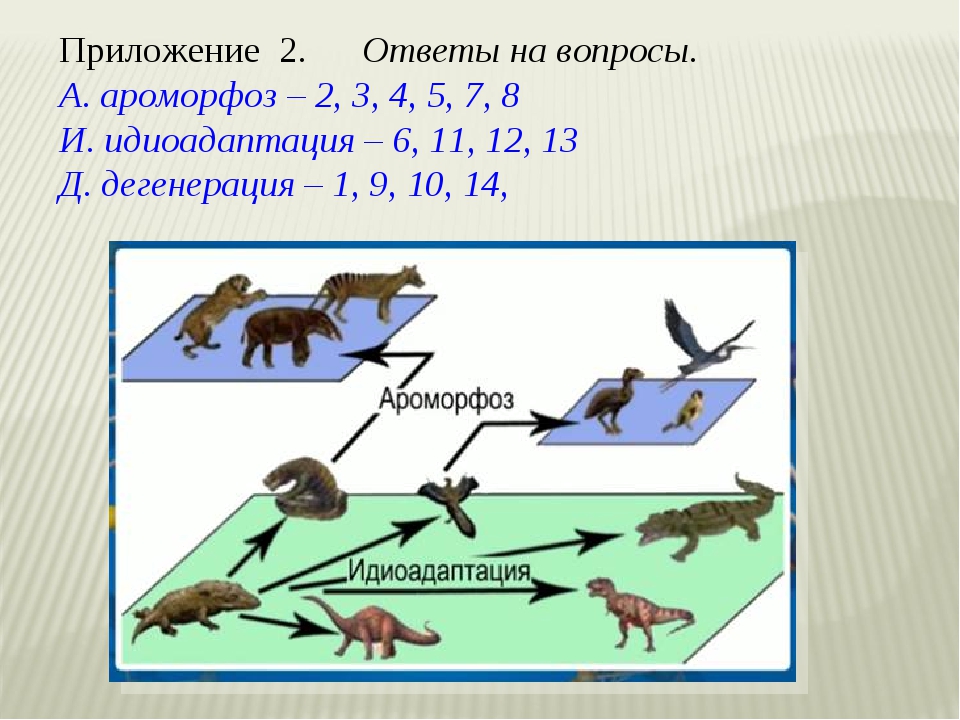 Р. и Андерсен, Н. М.: 1994, «Биология водно-болотных странников: взаимодействие между систематикой и экологией», Annual Review of Entomology 39 , 101–128.
Р. и Андерсен, Н. М.: 1994, «Биология водно-болотных странников: взаимодействие между систематикой и экологией», Annual Review of Entomology 39 , 101–128.
Артикул Google ученый
Спенс, Дж. Р. и Скаддер, Г. Г. Э .: 1980, «Среда обитания, жизненные циклы и структура гильдии водно-болотных странников (Heteroptera: Gerridae) на плато Фрейзер в Британской Колумбии», Канадский энтомолог 112 , 779–792.
Артикул Google ученый
Stearns, S.C .: 1976, «Life-history Tactics: A Review of the Ideas», Quarterly Review of Biology 51 , 3–47.
Артикул Google ученый
Stearns, S.C .: 1992, The Evolution of Life History , Oxford University Press, Oxford.
Google ученый
Тейлор П.: 1989, «Пересмотр моделей и генерирующая теория», Oikos . 54 , 121–126.
Артикул Google ученый
Торнхилл Р. и Олкок Дж .: 1983, Эволюция систем спаривания насекомых , Издательство Гарвардского университета, Кембридж, Массачусетс.
Google ученый
Vepsäläinen, K .: 1974, «Длина крыльев, репродуктивные стадии и среда обитания венгерского Gerris Fabr. Виды (Heteroptera, Gerridae) ’, Annales Academiae Scientiarum Fennicae, Series A IV, Biologica 202 , 1–18.
Виды (Heteroptera, Gerridae) ’, Annales Academiae Scientiarum Fennicae, Series A IV, Biologica 202 , 1–18.
Google ученый
Вепсяляйнен, К .: 1978, «Диморфизм крыла и диапауза у Gerris : определение и адаптивное значение», в Х. Дингле (ред.), Эволюция миграции и диапаузы насекомых , Springer-Verlag, Нью-Йорк, стр. 218–253.
Google ученый
Вепсяляйнен, К.и Ярвинен, О.: 1979, «Апомиктический партеногенез и структура окружающей среды», Американский зоолог . 19 , 739–751.
Google ученый
Вепсяляйнен, К. и Патама, Т .: 1983, «Распределение репродуктивной энергии в зависимости от структуры окружающей среды у пяти видов Gerris », в V.K. Браун и И. Ходек (ред.), Стратегии жизненного цикла , Dr. W. Junk bv Publ., Гаага, стр. 189–207.
Google ученый
Вепсяляйнен, К. и Писарски, Б .: 1982, «Ассамблея островных сообществ муравьев», Annales Zoologici Fennici 19 , 327–335.
Google ученый
Вепсяляйнен, К. и Саволайнен, Р .: 1990, «Влияние вмешательства муравьёв-муравьёв на добычу пищи Myrmica », Journal of Animal Ecology 59 , 643–654.
Артикул Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Вепсяляйнен, К. и Саволайнен, Р .: 1995, Рабочие соотношения полов и конфликт между полами у водного долгонога Gerris lacustris ’, American Naturalist 146 , 869–880.
Артикул Google ученый
Weizenbaum, J .: 1976, Компьютерная мощь и человеческий разум: от суждения к расчету , W.Х. Фриман и компания, Сан-Франциско.
Google ученый
Ямамура, Н .: 1987, «Эволюционно стабильные стратегии (ESS) Модели совокупления и защиты партнера», в Я. Ито, Дж. Л. Браун и Дж. Киккава (ред.), Общества животных: теории и факты , Японское научное общество, Токио, стр. 197–211.
Google ученый
(PDF) Обобщение в экологии и эволюционной биологии: от гипотезы к парадигме
236
Хейрстон, Н.Г.-старший: 1989, Экологические эксперименты, цель, дизайн и выполнение, Кембридж
University Press, Кембридж.
Яблонски, П. и Вепсяляйнен, К .: 1995, «Конфликт между полами у водолазов, Геррис
lacustris: проверка двух гипотез охранного поведения самцов», Поведенческая экология 6,
388–392.
Янзен, Д.: 1977, «Что такое одуванчики и тля?», American Naturalist 111, 586–589.
Klingenberg, C.P. и Спенс, Дж. Р.: 1997, «О роли размера тела в истории жизни
Эволюция», Экологическая энтомология 22, 55–68.
Кун, Т .: 1962, Структура научных революций, Издательство Чикагского университета, Чикаго.
Лентон Т.М .: 1998, «Гайя и естественный отбор», Nature 394, 439–447.
Лернер, I.M .: 1954, Генетический гомеостаз, Dover Publications, Нью-Йорк.
Левинс, Р .: 1966, «Стратегия построения модели в популяционной биологии», американский ученый
54, 421–431.
Левинс, Р .: 1968, Эволюция в меняющихся условиях, Princeton University Press, Princeton,
NJ.
Левинс, Р .: 1974, «Качественный анализ частично определенных систем», Annals of the New
York Academy of Sciences 231, 123–138.
Левинс, Р. и Левонтин, Р.: 1985, Диалектический биолог, издательство Гарвардского университета,
Кембридж, Массачусетс.
Лием, К.Ф .: 1990, «Ключевые эволюционные инновации, дифференциальное разнообразие и Symeco-
morphis», в M.H. Нитеки (ред.), Evolutionary Innovations, University of Chicago Press,
Chicago, стр.147–170.
Лавлок, Дж. Э .: 1979, Гайя, Новый взгляд на жизнь на Земле, Oxford Univeristy Press, Оксфорд.
Лавлок, Дж. Э .: 1991, Healing Gaia, Практическая медицина для планеты, Harmony Books, New
York.
MacArthur, R.H. и Wilson, E.O .: 1967, Theory of Island Biogeography, Princeton
University Press, Princeton, NJ.
McNaughton, S.J .: 1975, «r- & K-selection in Typha», American Naturalist 109, 251–261.
Мэйнард Смит, Дж.: 1978, Эволюция пола, Издательство Кембриджского университета, Кембридж.
Nummelin, M., Vepsäläinen, K. и Spence, JR: 1988, «Заражение яиц Gerrid (Heter-
optera: Gerridae) паразитоидом Tiphodytes gerriphagus Marchal (Hymenoptera:
Scelionidae) в Финляндии» Zoologici Fennici 25, 299–302.
О’Нил, Р.В., ДеАнджелис, Д.Л., Вайд, Дж. Б. и Аллен, Т. Х. Ф .: 1986, Иерархическая концепция
экосистем, Princeton University Press, Принстон, Нью-Джерси.
Питерс, Р.Х .: 1976, «Тавтология в эволюции и экологии», American Naturalist 110, 1–12.
Пьянка, E.R .: 1970, «Выбор r и k», American Naturalist 104, 592–597.
Пианка, ER: 1972, ‘r and K Selection or b and d Selection’, American Naturalist 106, 581–
588.
Поппер, KR: 1965, Предположения и опровержения: рост научных знаний, Harper
Torchbooks, Нью-Йорк.
Пуччиа, К. Дж. и Левинс, Р .: 1986, Качественное моделирование сложных биологических систем:
Дж. и Левинс, Р .: 1986, Качественное моделирование сложных биологических систем:
Введение в анализ петель и усреднение по времени, Издательство Гарвардского университета, Кембридж,
Массачусетс.
Рофф, Д.А.: 1992, Эволюция историй жизни: теория и анализ, Чепмен и Холл,
Нью-Йорк.
Роу, Л .: 1992, «Комфортная полиандрия у водного долгонога: конфликты при поиске пищи и самки
Контроль частоты совокупления и продолжительности сторожа», Animal Behavior 44, 189–
202.
Использование принципов теории обучения понять эволюцию развивающей организации
Связывание эволюции эволюционируемости с обобщением в обучающих системах
Объяснение того, как организмы приспосабливаются к новой селективной среде, занимает центральное место в эволюционной биологии [1–5].Живые организмы устойчивы и способны к изменениям. Первое свойство обеспечивает стабильность и надежную функциональность против генетических и экологических возмущений, в то время как последнее обеспечивает гибкость, позволяющую эволюционно приобретать новые потенциально адаптивные черты [5–9]. Эта способность организма производить подходящие фенотипические вариации для адаптации к новой среде часто определяется как предпосылка для эволюционируемости , то есть способности к адаптивной эволюции [7, 10, 11].Таким образом, важно понимать лежащие в основе вариационные механизмы, которые делают возможным производство адаптивных фенотипических вариаций [6, 7, 12–18].
Фенотипические вариации в значительной степени определяются внутренними тенденциями, налагаемыми генетической архитектурой и архитектурой развития [18–21]. Например, отклонения в развитии могут допускать высокую вариабельность одного фенотипического признака и ограниченную вариабельность другого или вызывать совместное изменение определенных фенотипических признаков [6, 15, 22–26]. Сами процессы развития также определяются предыдущим отбором.В результате мы можем ожидать, что прошлая эволюция могла бы адаптировать распределение фенотипов, исследованных будущим естественным отбором, чтобы усилить многообещающие вариации и избежать менее полезных, развивая архитектуры развития, которые предрасположены к эффективной адаптации [10, 13]. Однако отбор не может отдавать предпочтение чертам за преимущества, которые еще не реализованы. Более того, в ситуациях, когда отбор может контролировать фенотипическую изменчивость, он почти всегда снижает такую вариабельность, потому что он предпочитает канализацию гибкости [23, 27–29].
Однако отбор не может отдавать предпочтение чертам за преимущества, которые еще не реализованы. Более того, в ситуациях, когда отбор может контролировать фенотипическую изменчивость, он почти всегда снижает такую вариабельность, потому что он предпочитает канализацию гибкости [23, 27–29].
Канализация развития может показаться внутренне противоположной увеличению фенотипической изменчивости. Некоторые, однако, рассматривают эти понятия как две стороны одной медали, т.е. предрасположенность к развитию некоторых фенотипов, скорее, идет рука об руку с уменьшением предрасположенности к образованию других фенотипов [8, 30, 31]. Киршнер и Герхарт объединили результаты, подтверждающие эти идеи, в рамках единой модели облегченного варианта [8, 32]. Подобные идеи и концепции включают в себя вариационных свойств организмов [13], самооблегчение эволюции [20] и эволюцию как возня [33] и связанные с ними понятия [6, 7, 10, 12].В случае облегченной изменчивости ключевое наблюдение состоит в том, что внутренняя структура развития организмов смещает как количество, так и направление фенотипической изменчивости. Недавние работы в области облегченных вариаций показали, что для развития эволюционирующих структур необходимы множественные селективные среды [25, 27, 34–36]. Когда селективная среда содержит лежащие в основе структурные закономерности, возможно, что эволюция учится ограничивать фенотипическое пространство регионами, которые эволюционно более выгодны, способствуя открытию полезных фенотипов в одной или нескольких мутациях [35, 36].Но, как мы покажем, эти условия не обязательно повышают способность к развитию в новых средах. Таким образом, общие условия, которые способствуют возникновению адаптивных ограничений развития, повышающих способность к эволюции, не совсем понятны.
Чтобы решить эту проблему, мы изучаем условия, при которых эволюция путем естественного отбора может найти организации развития, которые производят то, что мы называем здесь обобщенными фенотипическими распределениями — т. е. прошлого, но они также могут производить новые фенотипы из того же семейства.Parter et al. уже было показано, что это возможно в конкретных случаях, изучая модели структур РНК и логических вентилей [34]. Здесь мы хотим понять более общие условия, при которых и в какой степени естественный отбор может усилить способность структур развития производить подходящие вариации для отбора в будущем. Мы следим за предыдущей работой по эволюции развития [25] с помощью компьютерного моделирования, основанного на моделях генно-регуляторной сети (GRN). Многие авторы отмечают, что GRN имеют общие функции с искусственными нейронными сетями [25, 37-40].Watson et al. продемонстрировал еще один результат, более важный для наших целей здесь; что эволюция регуляторных взаимодействий при естественном отборе математически эквивалентна тому, как нейронные сети изучают [25]. Во время эволюции GRN способна запоминать несколько фенотипов, которые соответствовали множеству прошлых селективных сред, усваивая их статистическую корреляционную структуру в своих онтогенетических взаимодействиях, точно так же, как обучающиеся нейронные сети хранят и вспоминают обучающие шаблоны.Фенотипы, которые соответствовали прошлому, затем могут быть воссозданы сетью спонтанно (при генетическом дрейфе без отбора) в будущем или как ответ на новые селективные среды, которые частично похожи на среды прошлого [25]. Важным аспектом упомянутых выше усовершенствованных систем является модульность. Модульность была ключевой особенностью работы по эволюционируемости [6, 29, 41, 42], направленной на облегчение изменчивости, которая уважает естественную разложимую структуру избирательной среды, т.е.е., держите вместе вещи, которые нужно держать вместе, и разделяйте вещи, которые независимы [6, 12, 20, 41]. Соответственно, система может выполнять простую форму обобщения, отделяя знания от контекста, в котором они первоначально наблюдались, и повторно применяя их в новых ситуациях.
е. прошлого, но они также могут производить новые фенотипы из того же семейства.Parter et al. уже было показано, что это возможно в конкретных случаях, изучая модели структур РНК и логических вентилей [34]. Здесь мы хотим понять более общие условия, при которых и в какой степени естественный отбор может усилить способность структур развития производить подходящие вариации для отбора в будущем. Мы следим за предыдущей работой по эволюции развития [25] с помощью компьютерного моделирования, основанного на моделях генно-регуляторной сети (GRN). Многие авторы отмечают, что GRN имеют общие функции с искусственными нейронными сетями [25, 37-40].Watson et al. продемонстрировал еще один результат, более важный для наших целей здесь; что эволюция регуляторных взаимодействий при естественном отборе математически эквивалентна тому, как нейронные сети изучают [25]. Во время эволюции GRN способна запоминать несколько фенотипов, которые соответствовали множеству прошлых селективных сред, усваивая их статистическую корреляционную структуру в своих онтогенетических взаимодействиях, точно так же, как обучающиеся нейронные сети хранят и вспоминают обучающие шаблоны.Фенотипы, которые соответствовали прошлому, затем могут быть воссозданы сетью спонтанно (при генетическом дрейфе без отбора) в будущем или как ответ на новые селективные среды, которые частично похожи на среды прошлого [25]. Важным аспектом упомянутых выше усовершенствованных систем является модульность. Модульность была ключевой особенностью работы по эволюционируемости [6, 29, 41, 42], направленной на облегчение изменчивости, которая уважает естественную разложимую структуру избирательной среды, т.е.е., держите вместе вещи, которые нужно держать вместе, и разделяйте вещи, которые независимы [6, 12, 20, 41]. Соответственно, система может выполнять простую форму обобщения, отделяя знания от контекста, в котором они первоначально наблюдались, и повторно применяя их в новых ситуациях.
Здесь мы показываем, что эта функциональная эквивалентность между обучением и эволюцией предсказывает эволюционные условия, которые делают возможной эволюцию обобщенных фенотипических распределений.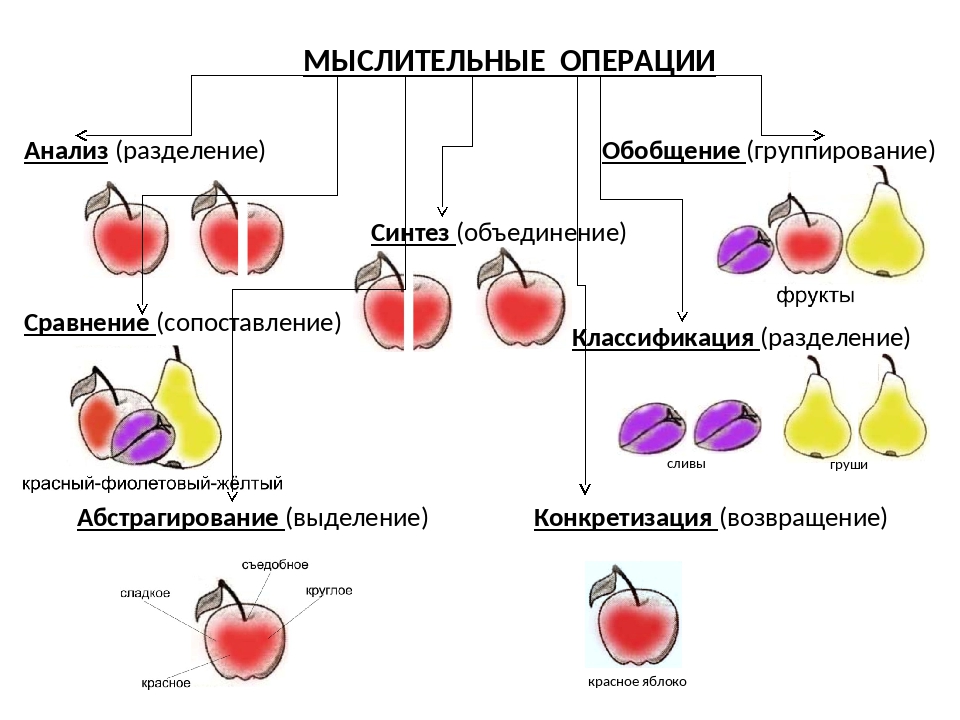 Мы проверяем эту аналогию между обучением и эволюцией, проверяя ее предсказания.В частности, мы разрешаем противоречие между канализацией фенотипов, которые были успешными в прошлых средах, и ожиданием фенотипов, которые подходят для будущих сред, признавая, что это эквивалентно предсказанию в обучающих системах. Такая прогностическая способность вытекает просто из способности представлять структурные закономерности в ранее увиденных наблюдениях (т. Е. В обучающей выборке), которые также верны в еще невидимых (т. Е. В тестовой выборке). В обучающих системах такое обобщение является обычным делом и не считается загадочным.Но также понятно, что успешное обобщение в обучающих системах не само собой разумеющееся и требует определенных хорошо понятных условий. Мы утверждаем, что понимание эволюции развития формально аналогично моделированию обучения и может предоставить полезные идеи и проверяемые гипотезы об условиях, которые усиливают эволюцию эволюционируемости при естественном отборе [42, 43]. Таким образом, признавая, что системы обучения на самом деле не « заглядывают в будущее », но тем не менее могут делать полезные прогнозы, обобщая прошлый опыт, мы демистифицируем представление о том, что близорукий естественный отбор может создавать новые фенотипы, которые подходят для ранее невидимой избирательной среды. и, что более важно, мы можем предсказать общие условия, в которых это возможно.Эта функциональная эквивалентность обучения и эволюции дает множество интересных, проверяемых прогнозов (таблица 1).
Мы проверяем эту аналогию между обучением и эволюцией, проверяя ее предсказания.В частности, мы разрешаем противоречие между канализацией фенотипов, которые были успешными в прошлых средах, и ожиданием фенотипов, которые подходят для будущих сред, признавая, что это эквивалентно предсказанию в обучающих системах. Такая прогностическая способность вытекает просто из способности представлять структурные закономерности в ранее увиденных наблюдениях (т. Е. В обучающей выборке), которые также верны в еще невидимых (т. Е. В тестовой выборке). В обучающих системах такое обобщение является обычным делом и не считается загадочным.Но также понятно, что успешное обобщение в обучающих системах не само собой разумеющееся и требует определенных хорошо понятных условий. Мы утверждаем, что понимание эволюции развития формально аналогично моделированию обучения и может предоставить полезные идеи и проверяемые гипотезы об условиях, которые усиливают эволюцию эволюционируемости при естественном отборе [42, 43]. Таким образом, признавая, что системы обучения на самом деле не « заглядывают в будущее », но тем не менее могут делать полезные прогнозы, обобщая прошлый опыт, мы демистифицируем представление о том, что близорукий естественный отбор может создавать новые фенотипы, которые подходят для ранее невидимой избирательной среды. и, что более важно, мы можем предсказать общие условия, в которых это возможно.Эта функциональная эквивалентность обучения и эволюции дает множество интересных, проверяемых прогнозов (таблица 1).
Таблица 1. Прогнозы, сделанные путем переноса ключевых уроков теории обучения в эволюционную теорию.
Подтверждено экспериментом: † Условия, способствующие обобщенному фенотипическому распределению, ‡ Как обобщение изменяется с течением времени эволюции, ◇ Условия, облегчающие обобщенное фенотипическое распределение, и and Анализ чувствительности к параметрам, влияющим на фенотипическое обобщение.
https://doi.org/10.1371/journal. pcbi.1005358.t001
pcbi.1005358.t001
В частности, следующие эксперименты показывают, что методы, улучшающие обобщение в машинном обучении, соответствуют эволюционным условиям, которые способствуют обобщенному фенотипическому распределению и, следовательно, повышают эволюционируемость. В частности, мы описываем, как хорошо известные методы машинного обучения, такие как обучение с шумом и штрафование за сложность модели, которые улучшают обобщающую способность моделей обучения, имеют биологические аналоги и могут помочь нам понять, как шумная избирательная среда и прямой выбор влияет на воспроизведение. Стоимость регуляторных взаимодействий генов может повысить эволюционируемость сетей регуляции генов.Это гораздо более сложная и мощная форма обобщения, чем предыдущие представления, которые просто экстраполируют предыдущий опыт. Система не просто расширяет свое усвоенное поведение за пределы своей прошлой «известной» области. Вместо этого нас интересуют ситуации, когда система может создавать новые знания, обнаруживая и систематизируя возникающие закономерности из прошлого опыта, и, что более важно, как система отделяет эти знания от контекста, в котором они первоначально наблюдались, чтобы их можно было повторно использовать. -используется в новых ситуациях.
Некоторые эволюционные механизмы и условия были предложены как важные факторы для улучшения эволюционируемости. Некоторые из них касаются модификации генетической изменчивости (например, [36, 44, 45] и [46]), в то время как другие касаются природы избирательных сред и организации развития, включая множественные избирательные среды [36], разреженность [47], прямое избирательное давление на стоимость соединений (которое может вызвать модульность [27, 44] и иерархию [48]), низкие отклонения и ограничения развития [49] и стохастичность в GRN [50].В этой статье мы сосредотачиваемся на механизмах и условиях, которые можно объединить и лучше понять в терминах машинного обучения, и, что более важно, на том, как мы можем использовать хорошо зарекомендовавшую себя теорию в обучении для характеристики общих условий, при которых эволюционируемость повышается. Таким образом, мы предлагаем первую теорию, которая характеризует общие условия, которые способствуют эволюции организаций развития, которые обобщают информацию, полученную в результате прошлого отбора, как требуется для повышения способности к развитию в новых условиях.
Таким образом, мы предлагаем первую теорию, которая характеризует общие условия, которые способствуют эволюции организаций развития, которые обобщают информацию, полученную в результате прошлого отбора, как требуется для повышения способности к развитию в новых условиях.
Экспериментальная установка
Основная экспериментальная установка включает нелинейный рекуррентный GRN, который развивает эмбриональный фенотипический паттерн, G , во взрослый фенотип, P a , на который может действовать отбор [25]. Фенотип взрослого человека представляет профиль экспрессии гена, который является результатом динамики GRN. Эта динамика определяется регуляторными взаимодействиями генов в сети, B [38, 39, 47, 53, 54] (см. Модель развития в приложении S1).Мы оцениваем приспособленность данной генетической структуры на основе того, насколько близок развитый фенотип к целевому фенотипическому паттерну, S . S характеризует направление отбора по каждому фенотипическому признаку, то есть элементу профиля экспрессии гена, в текущей среде. Динамика избирательной среды моделируется переключением с одного целевого фенотипа на другой каждые K поколений. K выбрано значительно меньше, чем общее количество смоделированных поколений.Ниже мы измеряем эволюционное время в эпох , где каждая эпоха обозначает N T × K поколений и N T соответствует количеству целевых фенотипов. (Обратите внимание, что эпоха — это термин, который мы заимствовали из машинного обучения, и он не представляет геологическую шкалу времени.)
В следующих экспериментах все фенотипические мишени выбираются из одного класса (как в [25, 34]).Этот класс состоит из 8 различных модульных паттернов, которые соответствуют различным комбинациям подшаблонов. Каждый субпаттерн выполняет разные функции, как показано на рис. 1. Эта модульная структура гарантирует, что среды (и, следовательно, наиболее подходящие фенотипы в этих средах) имеют общие закономерности, т. Е. Все они построены из различных комбинаций из одной и той же среды. набор модулей. Затем мы можем проверить, может ли система на самом деле «изучить» эту систематичность на ограниченном наборе примеров и, таким образом, обобщить их для создания новых фенотипов внутри того же класса.Наши эксперименты проводятся следующим образом. Популяция эволюционирует в результате воздействия ограниченного числа селективных сред (обучение). Затем мы анализируем условия, при которых производятся новые фенотипы из того же семейства (тест). В качестве примера задачи мы выбираем обучающий набор, состоящий из трех фенотипических паттернов из класса (см. Рис. 2а).
1. Эта модульная структура гарантирует, что среды (и, следовательно, наиболее подходящие фенотипы в этих средах) имеют общие закономерности, т. Е. Все они построены из различных комбинаций из одной и той же среды. набор модулей. Затем мы можем проверить, может ли система на самом деле «изучить» эту систематичность на ограниченном наборе примеров и, таким образом, обобщить их для создания новых фенотипов внутри того же класса.Наши эксперименты проводятся следующим образом. Популяция эволюционирует в результате воздействия ограниченного числа селективных сред (обучение). Затем мы анализируем условия, при которых производятся новые фенотипы из того же семейства (тест). В качестве примера задачи мы выбираем обучающий набор, состоящий из трех фенотипических паттернов из класса (см. Рис. 2а).
Рис. 1. Графическое изображение фенотипов.
(вверху) Схематическое изображение отображения последовательностей фенотипических паттернов на графические признаки.Каждый фенотипический «слот» представляет собой набор признаков (здесь 4), контролирующих определенный аспект фенотипа (например, передние крылья, жужжальца и усики). В пределах возможных конфигураций в каждом слоте (здесь 16) есть две конкретные конфигурации (состояние A и B), которые подходят для той или иной среды (см. Модель развития в Приложении S1). Например, «+ + −−» во втором слоте (сверху, зеленый) фенотипического паттерна кодирует пару передних крыльев (состояние B), а «- — ++» кодирует их отсутствие (состояние A ).Состояния A и B являются дополнением друг друга, то есть не соседями в пространстве фенотипов. Все остальные промежуточные состояния (здесь 14) представлены случайным мозаичным изображением состояний A и B на основе их соответствующего расстояния. d A указывает расстояние Хэмминга между данным состоянием и состоянием A. Соответственно, существуют потенциальные промежуточные состояния (т. Е. 4 для d A = 1, 6 для d A = 2 и 4 для d A = 3). (Внизу) Графическое изображение всех фенотипов, которые идеально адаптированы к каждой из восьми различных сред. Каждый целевой фенотип аналогичен насекомоподобному организму, имеющему 4 функциональных признака. Серые фенотипические мишени соответствуют побитовым дополнительным паттернам фенотипов в верхней половине пространства. Например, у самого правого верхнего насекомого присутствуют усики, передние и задние крылья, а хвоста нет. У самого правого, нижнего насекомого (побитовое дополнение насекомого над ним) усики, передние и задние крылья отсутствуют, но хвост присутствует.Мы определяем верхнюю строку как «класс» и игнорируем нижние дополнения как вырожденные формы обобщения.
(Внизу) Графическое изображение всех фенотипов, которые идеально адаптированы к каждой из восьми различных сред. Каждый целевой фенотип аналогичен насекомоподобному организму, имеющему 4 функциональных признака. Серые фенотипические мишени соответствуют побитовым дополнительным паттернам фенотипов в верхней половине пространства. Например, у самого правого верхнего насекомого присутствуют усики, передние и задние крылья, а хвоста нет. У самого правого, нижнего насекомого (побитовое дополнение насекомого над ним) усики, передние и задние крылья отсутствуют, но хвост присутствует.Мы определяем верхнюю строку как «класс» и игнорируем нижние дополнения как вырожденные формы обобщения.
https://doi.org/10.1371/journal.pcbi.1005358.g001
Рис. 2. Условия, способствующие обобщенному фенотипическому распределению.
Потенциальные фенотипические распределения, вызванные эволюционным процессом при 1) различных временных масштабах переключения среды, 2) шумах окружающей среды ( κ = 35 × 10 −4 ) и 3) давлении прямого отбора для слабых ( λ = 38) и разреженная связность ( λ = 0.22). Организмы подвергались воздействию трех селективных сред (а) из общего класса (i). Запоминание в процессе развития прошлых фенотипических целей явно зависит от временного масштаба изменения окружающей среды. Шумная среда и давление скупости усиливают обобщающую способность разработки, предрасполагая к созданию ранее невидимых целей из этого класса. Размер насекомоподобных существ описывает относительные частоты и указывает на склонность развития к выражению соответствующего фенотипа (фенотипы с частотой менее 0.01 были проигнорированы). Обратите внимание, что начальная структура развития в равной степени представляла все возможные фенотипические паттерны (здесь 2 12 возможных фенотипа).
https://doi.org/10.1371/journal.pcbi.1005358.g002
Один из способов оценить способность организаций развития к обобщению — это вывести популяцию в новую селективную среду и оценить сложившуюся предрасположенность системы развития к созданию подходящих фенотипы для этих сред (согласно [34]). Мы делаем это в конце экспериментального раздела. Мы также используем более строгий тест и исследуем спонтанное производство таких фенотипов, вызванное развитием в результате случайных генетических вариаций. В частности, мы исследуем, какие фенотипы предрасположены к возникновению сформировавшихся ограничений развития и предубеждений B , исходя из случайных исходных уровней экспрессии генов, G . Для этого мы проводим апостериорный анализ. Во-первых, мы оцениваем фенотипические распределения, вызванные развитой архитектурой развития при дрейфе.Поскольку мутация прямого воздействия на эмбриональные фенотипы ( G ) в этой модели намного больше, чем мутация на регуляторные взаимодействия ( B ) (см. Методы), мы оцениваем дрейф с равномерно случайным распределением по G (сохраняя B постоянная). Затем мы оцениваем, насколько успешно развитая система производит фенотипы с высокой приспособленностью, наблюдая, являются ли фенотипы, производимые развитыми корреляциями, B , членами общего класса (см. Методы).
Мы делаем это в конце экспериментального раздела. Мы также используем более строгий тест и исследуем спонтанное производство таких фенотипов, вызванное развитием в результате случайных генетических вариаций. В частности, мы исследуем, какие фенотипы предрасположены к возникновению сформировавшихся ограничений развития и предубеждений B , исходя из случайных исходных уровней экспрессии генов, G . Для этого мы проводим апостериорный анализ. Во-первых, мы оцениваем фенотипические распределения, вызванные развитой архитектурой развития при дрейфе.Поскольку мутация прямого воздействия на эмбриональные фенотипы ( G ) в этой модели намного больше, чем мутация на регуляторные взаимодействия ( B ) (см. Методы), мы оцениваем дрейф с равномерно случайным распределением по G (сохраняя B постоянная). Затем мы оцениваем, насколько успешно развитая система производит фенотипы с высокой приспособленностью, наблюдая, являются ли фенотипы, производимые развитыми корреляциями, B , членами общего класса (см. Методы).
Эволюция специализации — эволюционная биология
Знаковым документом в этой области в конце 20-го века является Футуйма и Морено 1988 г. (см. Также исторический обзор в Беренбауме, 1996 г.). Изучение специализации быстро развивалось в 1980-х годах и концентрировалось вокруг набора основных гипотез (например, гипотеза компромисса, гипотеза тупика; см. Компромиссы и специализация как эволюционный тупик). В последующие десятилетия появилось гораздо больше обзорных статей (например,g., Poisot, et al. 2011), что неудивительно, учитывая масштабность проблемы понимания эволюционной динамики экологической ниши. Эти обзорные статьи по-прежнему важны для определения исследовательских программ в различных областях (например, Bolnick, et al., 2003), но они не лишены своих предубеждений. Например, эта область часто фокусируется на специализации в трофических взаимодействиях, исключая специализацию в среде обитания или абиотических ассоциациях; и многие исследователи (в том числе и настоящие авторы) часто обращали внимание на травоядных насекомых (например,г. , Forister, et al. 2012, Jaenike 1990), не только из-за их крайнего разнообразия и специализации, но и из-за легкости, с которой ими можно манипулировать и подвергать их воздействию альтернативных ресурсов. Без сомнения, примеры из растений и других животных оказали влияние во многих случаях, что можно увидеть во многих цитируемых здесь обзорах.
, Forister, et al. 2012, Jaenike 1990), не только из-за их крайнего разнообразия и специализации, но и из-за легкости, с которой ими можно манипулировать и подвергать их воздействию альтернативных ресурсов. Без сомнения, примеры из растений и других животных оказали влияние во многих случаях, что можно увидеть во многих цитируемых здесь обзорах.
Беренбаум, М. Р. 1996. Введение в симпозиум: Об эволюции специализации. Американский натуралист 148: S78 – S83.
DOI: 10.1086 / 285903
Эта статья представляет собой краткое введение в эволюцию специализации с кратким историческим обзором до середины 1990-х годов. Он фокусируется на трофических взаимодействиях и описывает некоторые из основных трудностей в изучении специализации, в том числе то, как наши человеческие и культурные предубеждения могут повлиять на ее определение.
Болник Д. И., Р. Сванбэк, Дж. А. Фордайс и др. 2003. Экология людей: частота и последствия индивидуальной специализации. Американский натуралист 161.1: 1–28.
DOI: 10.1086 / 343878
Эта очень влиятельная статья ставит под сомнение предположение о том, что популяции обычно представляют собой однородные совокупности индивидов с одинаковыми степенями и типами специализации. Напротив, убедительные эмпирические данные свидетельствуют о том, что относительно обобщенные популяции часто представляют собой разнородные совокупности более специализированных людей. Исследуются теоретические, экологические, эволюционные и природоохранные последствия этих открытий.
Брукс, Д. Р. и Д. А. Макленнан. 2002. Природа разнообразия: эволюционное путешествие открытий . Чикаго: Univ. Чикаго Пресс.
DOI: 10.7208 / chicago / 9780226922478.001.0001
Эта книга дает обширный и глубокий обзор истории, теории, методов и эмпирических исследований, связанных с концепцией экологической специализации. Среди многих важных идей он вводит концепцию «ложных универсалов» для описания видов с явно широким диапазоном рациона, которые на самом деле специализируются на конкретном ресурсе и отслеживают этот же ресурс у нескольких видов хозяев или жертв.
Forister, M. L., L. A. Dyer, M. S. Singer, J. O. Stireman и J. T. Lill. 2012. Пересмотр эволюции экологической специализации с акцентом на взаимодействия насекомых и растений. Экология 93.5: 981–991.
DOI: 10.1890 / 11-0650.1
В этой статье дается более свежий взгляд на многие вопросы, рассмотренные в основополагающем обзоре этой темы тридцатью годами ранее в Futuyma and Moreno 1988. Системы насекомых-растений используются в качестве модели для решения теоретических проблем. и технический прогресс в исследованиях некоторых основных идей, связанных с эволюцией специализации.Предоставляется проспект для будущих исследований в данной области.
Футуйма Д. Дж. И Г. Морено. 1988. Эволюция экологической специализации. Ежегодный обзор экологии и систематики 19: 207–233.
DOI: 10.1146 / annurev.es.19.110188.001231
Это знаменательный обзор, в котором обобщены основные идеи для данной области, и из этого документа можно извлечь уроки. Например, авторы отметили, что межвидовые сравнения (например, производительности на разных ресурсах), как правило, говорят нам больше о последствиях, чем о причинах специализации, что по-прежнему является важной идеей для исследователей, работающих над «-омикой». »Специализированных фенотипов в сравнительной структуре.
Янике, Дж. 1990. Специализация хозяев на насекомых-фитофагах. Ежегодный обзор экологии и систематики 21: 243–273.
DOI: 10.1146 / annurev.es.21.110190.001331
Эта статья представляет собой классический обзор с точки зрения наиболее изученной группы организмов в рамках специализированных исследований: насекомых, питающихся растениями. Несмотря на относительно узкую направленность, извлеченные уроки носят общий характер, включая важные различия между механизмами специализации, зависящими от плотности и независимыми от плотности, и раннюю количественную обработку факторов, связанных с частотным распределением ширины диеты.
Джонсон, С. Д., и К. Э. Штайнер. 2000. Обобщение против специализации в системах опыления растений. Тенденции в экологии и эволюции 15.4: 140–143.
DOI: 10.1016 / S0169-5347 (99) 01811-X
Наряду с обзором питающихся растениями насекомых в Jaenike 1990, в этой статье представлен широкий обзор с точки зрения другой относительно хорошо изученной группы: растений и опылителей. Вопрос о том, была ли степень специализации исторически переоцененной, решается с важным выводом о том, что системы опыления растений занимают все точки в континууме от чрезвычайно обобщенного до чрезвычайно специализированного.
Пуазо Т., Дж. Д. Бевер, А. Немри, П. Х. Тралл и М. Э. Хохберг. 2011. Концептуальные основы эволюции экологической специализации. Письма по экологии 14.9: 841–851.
DOI: 10.1111 / j.1461-0248.2011.01645.x
Как Forister, et al. 2012 г., эта статья представляет собой недавний обзор за три десятилетия, прошедшие после основополагающей работы Футуйма и Морено 1988, с особым акцентом на биотические аспекты специализации. Авторы разрабатывают амбициозную концептуальную основу, которая синтезирует множество идей, чтобы понять, как и когда развивается специализация и, в частности, как возникают взаимодействия специализированных видов.
Возникновение кооперации путем эволюционного обобщения
Как правило, естественный отбор не поддерживает стратегии, поддерживающие общее благо. За исключением генетически связанных организмов, любое действие, влекущее за собой снижение приспособленности, отбирается противоположным образом, независимо от того, насколько оно полезно для других людей (1–3). Как следствие, естественный отбор приводит к «нагрузке на сотрудничество» — набору возможностей сотрудничества, которые упускаются в ходе эволюции.
Однако есть решение, которое может сделать, по крайней мере в принципе, адаптивным любое кооперативное поведение, пока оно дает чистый положительный эффект.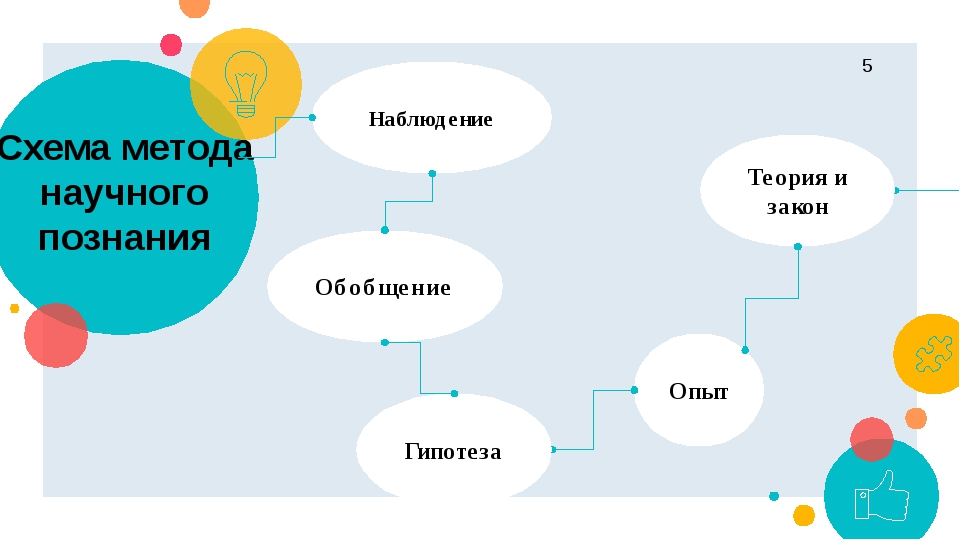 Его нужно стимулировать наградой (4–17). Если у человека есть возможность оплатить личные расходы c , чтобы принести выгоду b другому, последний может стимулировать сотрудничество, пообещав предоставить вознаграждение в размере r взамен. Это адаптивно для обоих лиц, пока r достаточно, чтобы компенсировать стоимость сотрудничества, но не настолько, чтобы перевесить его выгоду, то есть до тех пор, пока c
Его нужно стимулировать наградой (4–17). Если у человека есть возможность оплатить личные расходы c , чтобы принести выгоду b другому, последний может стимулировать сотрудничество, пообещав предоставить вознаграждение в размере r взамен. Это адаптивно для обоих лиц, пока r достаточно, чтобы компенсировать стоимость сотрудничества, но не настолько, чтобы перевесить его выгоду, то есть до тех пор, пока c
Однако с этим механизмом есть одна проблема. В отличие от рациональных людей, которые могут предвидеть последствия своих решений в будущем, биологическая эволюция близорука. Каждая генетическая мутация, связанная с биологической функцией, должна быть выгодной, когда она появляется. Как следствие, развитие взаимного сотрудничества порождает проблему самонастройки (18, 19). Сотрудничество не может быть адаптивным без вознаграждения, потому что оно стоит дорого; и вознаграждения не адаптируются, если значительная часть людей уже не желает сотрудничать при условии наличия вознаграждения.Следовательно, развитие взаимного сотрудничества потребует одновременного появления двух функций вместе, что маловероятно.
Коммуникация — это еще один тип биологического взаимодействия, который, как известно, подвержен тому же типу проблемы самонастройки. Для коммуникации должны присутствовать две взаимодополняющие черты: сигнал и реакция, причем ни одна из них не является адаптивной. По этой причине общение не может развиваться с нуля. Однако биологи-эволюционисты давно поняли, как эволюция может решить эту проблему.Коммуникация может развиваться, если одна сторона — сигнал или ответ — сначала развивается по другой причине, а затем превращается в коммуникативную функцию (20–22).
В случае взаимного сотрудничества решение должно быть таким же. Взаимное сотрудничество может развиваться, если одна сторона проблемы сначала развивается по независимой причине, а затем превращается в функцию взаимного сотрудничества (23, 24). Понимание возникновения взаимности и возможных ограничений, которые ограничивают ее масштабы в живом мире, поэтому включает понимание того, что делает возможным, а также какие ограничения, повторное использование биологической функции в другом контексте, чем это было изначально выбрано.Это то, к чему мы стремимся в данной статье.
Использование производительности когнитивных устройств за пределами диапазона контекстов, для которых они изначально были созданы, лежит в основе машинного обучения . Статистическая теория, лежащая в основе этого метода, показывает, что способность машин к обобщать не является случайным, удачным результатом обучения. Напротив, обобщение подчиняется систематической логике и происходит в конкретных обстоятельствах, которые можно понять (25).
Поскольку естественный отбор — это адаптивный процесс, как и обучение, концепция обобщения может быть легко распространена на биологическую эволюцию (26–35). В этой статье мы покажем, что взаимное сотрудничество может развиваться по той же причине, что и машины. Взаимное сотрудничество развивается, если факт сотрудничества и вознаграждения за сотрудничество других становится общими способностями, выходящими за пределы самого набора контекстов, для которых они были изначально выбраны, и условия, необходимые для такого эволюционного обобщения, могут быть поняты с помощью концепций машинного обучения.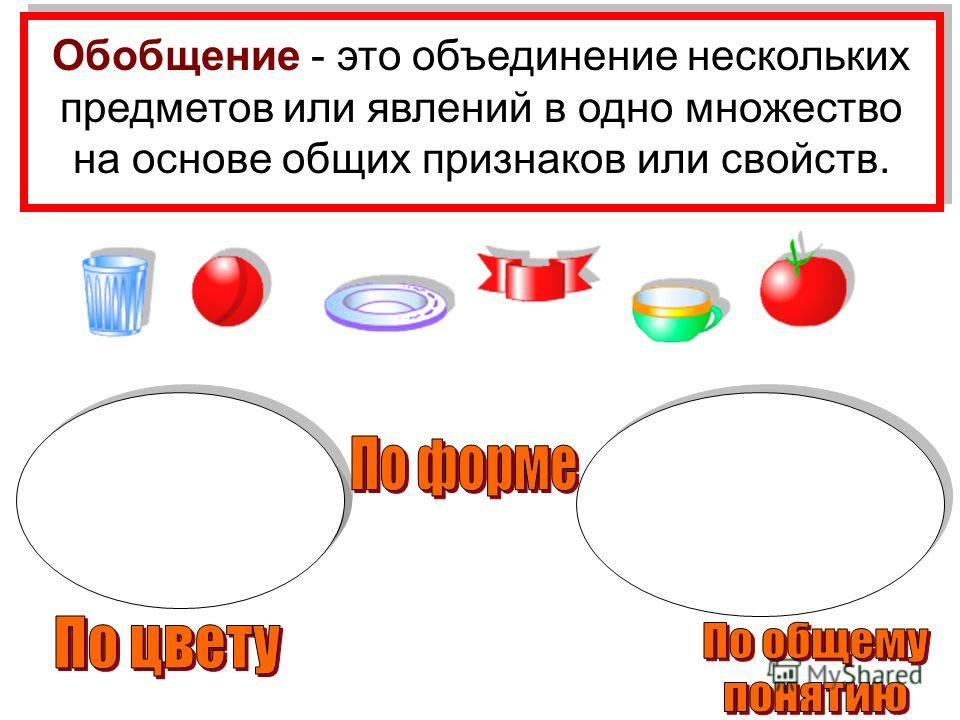 .Мы покажем, что это дает ключевое понимание природы взаимного сотрудничества в дикой природе, его ограничений у современных видов и его филогенетического распространения.
.Мы покажем, что это дает ключевое понимание природы взаимного сотрудничества в дикой природе, его ограничений у современных видов и его филогенетического распространения.
Жизненные игры
Теоретики эволюционных игр обычно изучают эволюцию социального поведения в одной игре, изолированной от всех других аспектов жизни людей. Это влечет за собой неявное предположение, что люди вырабатывают отдельные правила принятия решений, адаптированные к каждой игре, что плохо отражает то, как люди принимают решения на практике.Жизнь человека включает в себя множество ситуаций, и, хотя каждая ситуация влечет за собой определенную проблему, человек в конечном итоге должен принимать все решения с помощью одного и того же когнитивного устройства. Следовательно, модели должны отражать тот факт, что механизм принятия решений отдельных лиц выбирается для одновременной работы с набором игр, а не с одной (36–39). Мы увидим, что это имеет серьезные последствия для развития сотрудничества.
В нашей модели люди на протяжении своей жизни сталкиваются с набором ситуаций, выбранных в случайном порядке, каждая из которых называется игрой .В каждой игре участвуют два человека, доверенный и попечительский, оба характеризуются механизмом принятия решений, смоделированным как искусственная нейронная сеть, чьи веса связи развиваются в результате естественного отбора (подробнее см. Методы ). Для простоты мы предполагаем, что доверительный управляющий и доверительный управляющий взяты из двух разных групп, но в противном случае модель была бы почти идентичной. В каждой игре доверительный управляющий сначала решает, хочет ли он вложить деньги в игру или отказаться от нее. Если он решает инвестировать, он оплачивает личные расходы c , в то время как доверительный управляющий получает выгоду b .Затем доверительный управляющий имеет возможность принять решение о количественном вознаграждении r , которое он может передать доверительному управляющему взамен. Есть четыре типа игр в зависимости от значений b и c .
Есть четыре типа игр в зависимости от значений b и c .
В играх Selfish доверительный управляющий лично получает выгоду от инвестирования (то есть c <0), а доверительный управляющий может либо выиграть, либо не быть затронутым игрой (то есть b ≥ 0). Эгоистичные игры охватывают как ситуации, представляющие общий интерес, также называемые побочным продуктом сотрудничества (40), когда попечитель и попечитель извлекают выгоду из одного и того же действия, так и очень большое количество ситуаций, когда попечитель просто действует корыстно, не вовлекая попечитель.
В играх Wasteful инвестиции являются дорогостоящими для доверительного управляющего ( c> 0) и не приносят доверительному управляющему достаточной выгоды, чтобы компенсировать эти затраты ( b
В играх Cooperative инвестиции являются дорогостоящими для доверительного управляющего ( c> 0) и приносят прибыль b> c для доверительного управляющего.Следовательно, доверительный управляющий не имеет непосредственной выгоды от сотрудничества, но взаимовыгодное соглашение может быть заключено, если доверительный управляющий вознаграждает r ∈] c, b [. Кооперативные игры включают в себя все ситуации, в которых сотрудничество приносит чистую социальную выгоду, но создает эволюционную проблему самонастройки. В остальной части статьи для простоты мы будем говорить о «сотрудничестве» как о «инвестировании в кооперативную игру» (в несколько неортодоксальном использовании этого термина, см. 3). Наша цель — понять, как в этом смысле может развиваться сотрудничество.
Наконец, мы рассматриваем четвертую категорию игр, которую мы называем Взаимозависимые , охватывающие ситуации, когда (i) инвестиции приносят пользу доверительному управляющему и (ii) конечная чистая выгода для доверительного управляющего зависит от качества доверительного управляющего, то есть ситуации зависимых от партнера общих интересов (40–46). Например, когда колония муравьев вкладывает средства в защиту дерева акации, конечная выгода от инвестиций зависит от фактической ценности акации как укрытия (47).Когда опылитель посещает цветок, время, проведенное в цветке, может того стоить или нет, в зависимости от качества цветка (48). Когда человек тратит время и энергию на коллективные действия с сородичами (например, выполнение коллективной охоты, осмотр хищника и т. Д.), Конечная выгода от инвестиций зависит от качества партнера (ей) (например, 49). И т.д. Формально взаимозависимые игры характеризуются положительной стоимостью для доверительного управляющего ( c> 0), преимуществом b для доверительного управляющего и тем, что мы называем «автоматическим» вознаграждением r a , значение которого зависит от качества доверительного управляющего.На практике в нашей модели каждый раз, когда два игрока играют во взаимозависимую игру, значение автоматического вознаграждения просто выбирается случайным образом из двух возможных значений: r a 1 = 0 или r a 2 > c , каждый с вероятностью 0,5. Наконец, без ограничения общности, мы предполагаем, что b> c во всех взаимозависимых играх, так что всегда можно найти взаимовыгодное соглашение.
Например, когда колония муравьев вкладывает средства в защиту дерева акации, конечная выгода от инвестиций зависит от фактической ценности акации как укрытия (47).Когда опылитель посещает цветок, время, проведенное в цветке, может того стоить или нет, в зависимости от качества цветка (48). Когда человек тратит время и энергию на коллективные действия с сородичами (например, выполнение коллективной охоты, осмотр хищника и т. Д.), Конечная выгода от инвестиций зависит от качества партнера (ей) (например, 49). И т.д. Формально взаимозависимые игры характеризуются положительной стоимостью для доверительного управляющего ( c> 0), преимуществом b для доверительного управляющего и тем, что мы называем «автоматическим» вознаграждением r a , значение которого зависит от качества доверительного управляющего.На практике в нашей модели каждый раз, когда два игрока играют во взаимозависимую игру, значение автоматического вознаграждения просто выбирается случайным образом из двух возможных значений: r a 1 = 0 или r a 2 > c , каждый с вероятностью 0,5. Наконец, без ограничения общности, мы предполагаем, что b> c во всех взаимозависимых играх, так что всегда можно найти взаимовыгодное соглашение.
Во всех типах игр, помимо возможного автоматического вознаграждения, доверительный управляющий имеет возможность инвестировать в активных вознаграждений , r , чтобы предоставить дополнительную выгоду трастеру.Для простоты мы предполагаем, что это активное вознаграждение представляет собой консервативную передачу единиц пригодности с выгодой – для доверительного управляющего при точно таких же затратах для доверительного управляющего.
Обобщение создает скрытое вознаграждение
Мы предполагаем, что изначально, до того, как естественный отбор воздействует на поведение индивидов, попечители не награждают активно какую-либо игру (то есть r = 0 во всех случаях). В результате доверительные управляющие находятся в процессе отбора, чтобы отклонить все совместные игры, и, следовательно, их склонность вознаграждать эти игры не является предметом отбора, поскольку доверительные управляющие в любом случае никогда не вкладывают в них средства (см.1 и , методы ). В этом суть проблемы бутстрепа сотрудничества. Ключ к решению этой проблемы, однако, заключается в существовании скрытых вознаграждений , то есть вознаграждений, которые попечители предложили бы в совместных играх, если бы попечители действительно играли в них. Эволюция или неэволюция сотрудничества в решающей степени зависит от существования и ценности скрытых вознаграждений. Если скрытая награда в данной игре составляет r> c , то в этой игре может развиваться сотрудничество, поскольку эта награда представляет собой еще не обнаруженную выгоду, которую попечители отбирают для сбора.
В результате доверительные управляющие находятся в процессе отбора, чтобы отклонить все совместные игры, и, следовательно, их склонность вознаграждать эти игры не является предметом отбора, поскольку доверительные управляющие в любом случае никогда не вкладывают в них средства (см.1 и , методы ). В этом суть проблемы бутстрепа сотрудничества. Ключ к решению этой проблемы, однако, заключается в существовании скрытых вознаграждений , то есть вознаграждений, которые попечители предложили бы в совместных играх, если бы попечители действительно играли в них. Эволюция или неэволюция сотрудничества в решающей степени зависит от существования и ценности скрытых вознаграждений. Если скрытая награда в данной игре составляет r> c , то в этой игре может развиваться сотрудничество, поскольку эта награда представляет собой еще не обнаруженную выгоду, которую попечители отбирают для сбора.
(1) Доверительный управляющий информируется обо всех параметрах игры, которые не зависят от доверительного управляющего, и решает либо отклонить игру, либо «заплатить, чтобы увидеть» качество доверительного управляющего. (2) Если доверенное лицо отказывается, игра заканчивается без выплат, и следующая игра выбирается случайным образом. (3) Если доверитель решает «заплатить, чтобы увидеть», он платит небольшую стоимость мониторинга, м , чтобы узнать об общей награде ( руб. + руб. ), которую он в конечном итоге получит, если бы он решили сыграть в игру.Это предназначено для отражения всех разнообразных способов получения информации о своем партнере. Например, доверительный управляющий может собирать информацию, наблюдая за некоторыми аспектами фенотипа доверительного собственника, которые коррелируют с его качествами, играя в игру один раз, чтобы «проверить» доверительного собственника, узнавая о прошлом поведении доверительного собственника и т. Д. (4) На последнем этапе мы предполагаем, что трастер решает (5) продолжить игру, если r + r a > c или (6) отказаться, если нет.
Проблема в том, что эволюционная теория игр не может прямо сказать нам что-либо о скрытых вознаграждениях, поскольку они точно не подвергаются отбору. Тем не менее их эволюцию можно понять, если принять во внимание тот факт, что выбор фактических наград косвенно влияет на эволюцию скрытых наград. Чтобы понять этот момент, полезно провести параллель с машинным обучением. В машинном обучении машина (например, искусственная нейронная сеть) обучается отображать заданный набор входных данных, называемый обучающим набором, на определенные выходы.Но цель машинного обучения не только в том, чтобы создавать машины, которые могут давать правильные ответы в контекстах, с которыми они уже сталкивались. Смысл существования этой техники проистекает из того факта, что машины способны на обобщить за пределы своей обучающей выборки. То есть машины также могут принимать правильные решения в ситуациях, с которыми их никогда не учили. И эта способность к обобщению не является случайным, удачным результатом обучения. Напротив, обобщение подчиняется систематической логике и происходит в конкретных обстоятельствах, которые можно понять.
Будучи двумя адаптивными процессами, обучение и эволюция посредством естественного отбора обладают схожими свойствами, и поэтому обобщение имеет эволюционный аналог (32). Набор ситуаций, для которых популяция находится или находилась в отборе, соответствует обучающему набору популяции, то есть ситуациям, в которых популяция получила обратную связь от окружающей среды посредством естественного отбора. И наоборот, набор ситуаций, для которых популяция никогда не подвергалась отбору, потому что они возникают недостаточно часто, чтобы представлять собой давление отбора, соответствует набору тестов для этой популяции.Любой ответ на тестовой выборке — это непреднамеренное свойство способа обобщения организмов за пределами своей обучающей выборки.
В нашей модели обучающий набор опекунов состоит из набора игр, в которые доверительные управляющие действительно инвестируют. На протяжении всей своей жизни попечители сталкиваются с этими играми и должны принять решение о награде. Следовательно, как популяция, они получают обратную связь от естественного отбора. С другой стороны, набор для тестирования доверенных лиц состоит из набора игр, в которые доверенные лица никогда не вкладываются.Попечители никогда не должны принимать решения о награде в этих играх. Следовательно, они никогда не получают обратной связи от естественного отбора. Иными словами, набор для тестирования доверенных лиц состоит из всех игр, в которых их способность (или неспособность) вознаграждать вложения доверенных лиц составляет только скрытых . Поскольку развитие сотрудничества зависит от ценности этих скрытых вознаграждений, оно зависит от способности попечителей обобщать.
На протяжении всей своей жизни попечители сталкиваются с этими играми и должны принять решение о награде. Следовательно, как популяция, они получают обратную связь от естественного отбора. С другой стороны, набор для тестирования доверенных лиц состоит из набора игр, в которые доверенные лица никогда не вкладываются.Попечители никогда не должны принимать решения о награде в этих играх. Следовательно, они никогда не получают обратной связи от естественного отбора. Иными словами, набор для тестирования доверенных лиц состоит из всех игр, в которых их способность (или неспособность) вознаграждать вложения доверенных лиц составляет только скрытых . Поскольку развитие сотрудничества зависит от ценности этих скрытых вознаграждений, оно зависит от способности попечителей обобщать.
Чтобы изучить эволюцию скрытых вознаграждений в нашей мультиигровой структуре, мы сначала позволили попечителям развиваться, сохраняя поведение доверенных лиц как экзогенную константу.Мы предполагали, что доверительные управляющие всегда вкладывались в эгоистичные и взаимозависимые игры, и всегда отказывались от расточительных и кооперативных игр. На рисунке 2 показана эволюция поведения попечителей с течением времени. В состоянии равновесия попечители никогда не награждали эгоистичные игры, поскольку вознаграждение в этих играх было излишним, и они вознаграждали взаимозависимые игры добавлением активного вознаграждения r> c — r a , когда автоматическое вознаграждение оказалось недостаточным для компенсации попечителей ‘стоимость (то есть, когда r a
На оси ординат мы указываем процент игр каждого типа, для которых доверительные управляющие предоставляют достаточное вознаграждение, чтобы побудить их вкладывать средства. Сплошные линии показывают адаптацию вознаграждения попечителей во взаимозависимых и эгоистичных играх в течение эволюционного времени.Пунктирная зеленая линия показывает влияние этой адаптации на скрытую реакцию опекунов в совместных играх, для которых опекуны не подлежат отбору. Бесполезные игры здесь не показаны (см. SI Рисунок S1). Есть 2 5 взаимозависимых игр, N P = 1 вход для каждого параметра выплаты и N S = 50 ложных функций. Имею в виду более 30 симуляций. См. Методы для получения подробной информации о других параметрах. Обратите внимание, что в эгоистичных играх доверительный управляющий всегда имеет стимул инвестировать независимо от активного вознаграждения доверительного управляющего.Итак, здесь мы просто показываем среднее активное вознаграждение, предоставляемое в эгоистичных играх (синяя линия), которое всегда равно нулю.
Сплошные линии показывают адаптацию вознаграждения попечителей во взаимозависимых и эгоистичных играх в течение эволюционного времени.Пунктирная зеленая линия показывает влияние этой адаптации на скрытую реакцию опекунов в совместных играх, для которых опекуны не подлежат отбору. Бесполезные игры здесь не показаны (см. SI Рисунок S1). Есть 2 5 взаимозависимых игр, N P = 1 вход для каждого параметра выплаты и N S = 50 ложных функций. Имею в виду более 30 симуляций. См. Методы для получения подробной информации о других параметрах. Обратите внимание, что в эгоистичных играх доверительный управляющий всегда имеет стимул инвестировать независимо от активного вознаграждения доверительного управляющего.Итак, здесь мы просто показываем среднее активное вознаграждение, предоставляемое в эгоистичных играх (синяя линия), которое всегда равно нулю.
На рисунке 3 показано, как это обобщение зависело от обстоятельств. В частности, это в решающей степени зависело от преобладания взаимозависимых взаимодействий в экологии индивидов. Когда в среде было очень мало взаимозависимых игр, попечители почти никогда не делали обобщений в рамках эволюционного равновесия (рис. 3а). Они оптимально награждали в одной из немногих взаимозависимых игр, но ни при каких других обстоятельствах.С другой стороны, обобщение было более значительным, когда в экологии индивидов присутствовало множество взаимозависимых игр. Попечители не только оптимально вознаграждали во взаимозависимых играх, они также могли рассчитать правильное вознаграждение, которое они должны были предложить в некоторых, а иногда и во всех совместных играх, даже если они никогда не были выбраны для этого (рис. 3а).
Рис. 3. Степень обобщения в зависимости от экологии индивидов. (a, b) Мы предполагаем, что здесь развиваются только попечители, т.е.е. поведение трастера фиксировано (см. основной текст). Мы показываем процент кооперативных игр, для которых попечители в условиях эволюционного равновесия предоставляют скрытое вознаграждение, достаточное для стимулирования инвестиций доверительных управляющих. Каждая точка является результатом моделирования (для каждого набора параметров было выполнено 30 симуляций). Сплошная линия и заштрихованная область соответствуют наилучшему логистическому соответствию (среднее значение и стандартная ошибка). Результаты показаны для различного количества ложных и подлинных элементов ( N S и N P , см. Методы ). (c, d) Здесь мы предполагаем, что попечители и попечители развиваются совместно. Мы показываем процент кооперативных игр, в которые играют попечители и которые вознаграждаются попечителями при эволюционном равновесии. Условные обозначения и параметры такие же, как в (a), и (b), . (e, f) Гистограммы, показывающие распределение процента «обнаруженных» совместных игр по всем результатам моделирования, показанным, соответственно, в (c), и (d) .См. Методы для получения подробной информации о других параметрах.
Каждая точка является результатом моделирования (для каждого набора параметров было выполнено 30 симуляций). Сплошная линия и заштрихованная область соответствуют наилучшему логистическому соответствию (среднее значение и стандартная ошибка). Результаты показаны для различного количества ложных и подлинных элементов ( N S и N P , см. Методы ). (c, d) Здесь мы предполагаем, что попечители и попечители развиваются совместно. Мы показываем процент кооперативных игр, в которые играют попечители и которые вознаграждаются попечителями при эволюционном равновесии. Условные обозначения и параметры такие же, как в (a), и (b), . (e, f) Гистограммы, показывающие распределение процента «обнаруженных» совместных игр по всем результатам моделирования, показанным, соответственно, в (c), и (d) .См. Методы для получения подробной информации о других параметрах.
Причина, по которой небольшое количество взаимозависимых игр препятствует обобщению, та же, что и в хорошо известной проблеме чрезмерного согласования с несбалансированными данными в машинном обучении. Известно, что в задачах классификации, если обучающая выборка несбалансирована (например, в ней гораздо больше отрицательных, чем положительных примеров, или наоборот), то обобщение вряд ли возможно (50–54). Точнее, если количество положительных примеров в обучающем наборе очень мало, ИНС обычно учится классифицировать их точно, но слишком конкретно, снижая эффективность своей классификации в тестовом наборе, что называется переобучением.
Помимо преобладания взаимозависимости в экологии индивидов, возможность обобщения также зависела от когнитивной сложности задачи. Мы зафиксировали эту сложность с помощью двух параметров: N P и N S , которые представляли, соответственно, количество сенсорных входов, которые люди должны были интегрировать для оценки выигрыша в игре, и количество отвлекающих входов, которые могло быть ложно коррелировано с выигрышем в игре, но не допускало обобщения (см. Методы).Как это принято в машинном обучении, чем больше было N P и / или N S , тем больше было количество взаимозависимых игр, которые требовались для достижения полного обобщения (рис. 3a и 3b).
Методы).Как это принято в машинном обучении, чем больше было N P и / или N S , тем больше было количество взаимозависимых игр, которые требовались для достижения полного обобщения (рис. 3a и 3b).
Скрытые вознаграждения способствуют еще большему обобщению
В дополнение к эволюции попечителей мы затем учли тот факт, что доверительные управляющие также сформированы естественным отбором (см. Методы ). Обобщение — или эквивалентный процесс (см. Рис.4) — может происходить с обеих сторон, усиливая друг друга и приводя в конечном итоге к открытию большего количества совместных игр. Совместная эволюция сотрудничества и вознаграждения в этом случае схематически представлена на рисунке 4, а пример показан на рисунке 5. По сравнению с односторонним случаем, двусторонний случай привел к более высоким уровням обобщения (рисунок 3c).
Рис. 4. Эволюционные шаги, ведущие к возникновению сотрудничества. (1) Трастер выбран для инвестирования во взаимозависимые игры.(2) В результате попечитель выбирается для вознаграждения доверенного лица во взаимозависимых играх, когда само по себе автоматическое вознаграждение оказывается недостаточным. (3) В качестве побочного продукта попечитель также имеет скрытую способность вознаграждать некоторые совместные игры. (4) Доверительный управляющий выбирается, чтобы воспользоваться скрытыми вознаграждениями, инвестируя в эти Кооперативные игры. (5) Несмотря на то, что доверительный управляющий находится в процессе выбора, чтобы не вкладывать средства в другие совместные игры, на практике трудно избежать каждого ложного срабатывания.Следовательно, доверительный управляющий инвестирует в некоторые совместные игры, которые он еще не награждает. Эти ложные срабатывания играют ту же роль, что и скрытые вознаграждения, только наоборот. Их существование порождает давление выбора на попечителя, чтобы вознаградить игры, в которые попечитель случайно инвестирует. (6) В результате набор игр, для которых выбирается вознаграждение, увеличивается, а обобщение становится более сильным на стороне попечителя. (7) Поскольку появляется все больше и больше кооперативных игр, в которые можно инвестировать, доверитель делает дополнительные ложные срабатывания.
(7) Поскольку появляется все больше и больше кооперативных игр, в которые можно инвестировать, доверитель делает дополнительные ложные срабатывания.
Мы показываем по 8 игр каждого типа, выбранных случайным образом. Для взаимозависимых игр мы показываем только активное вознаграждение, r , которое предлагают доверенные лица, когда оказывается, что автоматическое вознаграждение отсутствует (т.е. когда r a = 0). Обратите внимание, что для каждой игры мы показываем масштабированное значение r / b для вознаграждения, так что это значение равно 1, когда попечители предоставляют достаточное вознаграждение.См. Методы для получения более подробной информации и параметров.
Помимо количества взаимозависимых игр и когнитивной сложности задачи, развитие сотрудничества также зависело от относительной важности взаимозависимых и кооперативных игр для экологии людей. Чем большую роль в экологии индивидов играет кооперация в целом, тем проще эволюционное обобщение (рис. 6).
Рис. 6. Экологическая важность сотрудничества и эволюционное обобщениеМы показываем равновесный процент совместных игр, в которые играют попечители и вознаграждаются попечителями, в зависимости от (i) доли взаимозависимых игр и (ii) доли взаимозависимых игр. относительная экологическая важность сотрудничества α (относительная важность взаимозависимых и кооперативных игр для фитнеса по сравнению с эгоистичными и расточительными играми, см. методы ).Случай α = 0 соответствует результатам на рис. 3c и 3d. Результаты показаны для двух значений числа N S ложных признаков и для двух значений числа N P входных данных для каждого параметра выплаты. Каждая точка — это среднее значение по 30 симуляциям.
Обсуждение
Эволюция путем естественного отбора требует, чтобы всегда один и тот же человек проявлял черту и извлекал из нее пользу. Каждый раз, когда это ограничение частично снималось в ходе эволюции — посредством генетического родства или посредством эволюции совместной репликации — происходил крупный переход, поскольку это расширяло диапазон адаптивных черт, которые могли развиваться (55, 56).В принципе, взаимность должна быть самым мощным средством снятия этого ограничения, поскольку она может вызвать эволюцию любой черты, с помощью которой индивид приносит пользу любому другому, при том единственном условии, что последний может вернуть некоторую выгоду. .
Каждый раз, когда это ограничение частично снималось в ходе эволюции — посредством генетического родства или посредством эволюции совместной репликации — происходил крупный переход, поскольку это расширяло диапазон адаптивных черт, которые могли развиваться (55, 56).В принципе, взаимность должна быть самым мощным средством снятия этого ограничения, поскольку она может вызвать эволюцию любой черты, с помощью которой индивид приносит пользу любому другому, при том единственном условии, что последний может вернуть некоторую выгоду. .
Однако то, что делает этот механизм таким мощным, имеет и обратную сторону эволюции. Взаимное сотрудничество сталкивается с проблемой курицы и яйца. Способность вознаграждать сотрудничество других является адаптацией к возможности сотрудничества других, но сама эта возможность является адаптацией к тому факту, что сотрудничество вознаграждается.Это, вероятно, объясняет, почему, несмотря на свою силу, взаимное сотрудничество играет гораздо более ограниченную роль в основных переходах, чем другие механизмы сотрудничества (57).
В этой статье мы попытались понять, какие эволюционные механизмы могут объяснить возникновение взаимного сотрудничества, несмотря на эту проблему. Мы показали, что это возникновение возможно при совместном воздействии двух механизмов: (i) селективного давления, возникающего в результате конкретных экологических ситуаций, которые играют роль триггеров, и (ii) эволюционного механизма, аналогичного обобщению.
Триггерные экологические ситуации — это социальные взаимодействия, при которых индивиды имеют степень зависимости от партнера общих интересов с другими. Типичными примерами этого являются коллективные действия, в которых заинтересованы оба партнера. Выгода, которую каждая сторона извлекает из коллективных действий, зависит от качества другой. Учитывая, что несколько партнеров или, в более общем плане, несколько различных возможностей для участия в коллективных действиях могут конкурировать друг с другом, естественно, что некоторые из них являются прибыльными, а некоторые — нет (58). Это означает возможность сотрудничества при условии качества партнера. В свою очередь, эта способность создает давление отбора в пользу людей высокого качества, то есть людей, которые дают другим достаточно большое вознаграждение, чтобы компенсировать стоимость их сотрудничества. Следовательно, в эволюционном равновесии люди взаимодействуют друг с другом, без каких-либо проблем с курицей и яйцом.
Это означает возможность сотрудничества при условии качества партнера. В свою очередь, эта способность создает давление отбора в пользу людей высокого качества, то есть людей, которые дают другим достаточно большое вознаграждение, чтобы компенсировать стоимость их сотрудничества. Следовательно, в эволюционном равновесии люди взаимодействуют друг с другом, без каких-либо проблем с курицей и яйцом.
С другой стороны, у этих реципрокных механизмов нет причин действовать вне конкретных взаимодействий, в которых они изначально развивались.Это объясняет, почему подавляющее большинство, а возможно, и все случаи взаимного сотрудничества, задокументированные за пределами людей, на самом деле соответствуют ситуациям, когда люди имеют некоторую степень общего интереса к сотрудничеству (46, 59). Взаимное сотрудничество ведет к количественной эскалации форм сотрудничества, которые уже были выгодными с самого начала при отсутствии взаимности. Он не допускает появления новых форм сотрудничества, которые обязательно должны основываться на взаимности, чтобы быть адаптивными.Иными словами, как отметили некоторые ученые, сотрудничество на природе чаще встречается в играх в снежные заносы, где у людей частично совпадают интересы, чем в дилеммах заключенных, где у индивидов нет общих интересов (59–61).
Следовательно, взаимное сотрудничество на самом деле не выполняет своего обещания привести к сотрудничеству во всех ситуациях, когда люди могут приносить пользу друг другу. На практике его объем значительно более ограничен. Если бы особи вида смогли преодолеть это ограничение, они бы резко расширили спектр выгод, которые они могут получить.
В этой статье мы показали, что это возможно, если экология человека включает не только несколько триггерных ситуаций, но и большое их количество в самых разных сферах их жизни. В этом случае отбор отдает предпочтение взаимным механизмам, которые могут обобщать ситуации, выходящие за рамки тех ситуаций, для которых они были изначально выбраны. Отдельные лица являются не только хорошими партнерами по охоте или хорошими производителями нектара для привлечения опылителей, они скорее могут в целом оценить затраты, уплачиваемые другими при сотрудничестве, и гарантировать, что они окупятся, сотрудничая с нужной суммой взамен.И наоборот, у людей нет причин сотрудничать только тогда, когда они непосредственно заинтересованы в этом. Скорее, они стремятся сотрудничать всякий раз, когда у них есть возможность принести пользу кому-то другому, при условии, что у нее есть возможность дать выгоду взамен.
Отдельные лица являются не только хорошими партнерами по охоте или хорошими производителями нектара для привлечения опылителей, они скорее могут в целом оценить затраты, уплачиваемые другими при сотрудничестве, и гарантировать, что они окупятся, сотрудничая с нужной суммой взамен.И наоборот, у людей нет причин сотрудничать только тогда, когда они непосредственно заинтересованы в этом. Скорее, они стремятся сотрудничать всякий раз, когда у них есть возможность принести пользу кому-то другому, при условии, что у нее есть возможность дать выгоду взамен.
Насколько нам известно, человеческий вид — единственный, в котором развились общие механизмы такого рода. Наше сотрудничество, основанное на справедливости, заключается в подсчете затрат, которые несут другие при сотрудничестве, какими бы они ни были, и их возмещении путем сотрудничества любым возможным способом (62–64).Это позволяет нам оказывать положительное влияние друг на друга в чрезвычайно широком диапазоне ситуаций, по сравнению с другими видами, в которых взаимное сотрудничество ограничивается несколькими ситуациями, в которых по случайным причинам проблема самонастройки может быть преодолена.
Методы
Инициализация
Мы запускаем симуляции, закодированные на Python. В каждой симуляции генерируется набор игр, который содержит 500 эгоистичных игр, 250 кооперативных игр, 250 расточительных игр и ряд взаимозависимых игр, которые варьируются от 1 до 2 10 в зависимости от симуляции.Для каждой игры мы генерируем случайные параметры выигрыша ( c ∈ [−1,1], b ∈ [0,1] и r a ∈ [0,1] для взаимозависимых игр) при условиях предоставлено в разделе Игры жизни .
В реальной жизни люди не имеют прямого представления о затратах и выгодах взаимодействия, в которое они вовлечены. Скорее они должны принимать решения на основе различных сенсорных сигналов из окружающей среды. Некоторые входные данные несут частичную информацию о параметрах выигрыша, в то время как другие представляют собой просто ложные характеристики, которые случайно коррелируют с текущей игрой. В наших симуляциях для каждого параметра выигрыша игроки получают не его фактическое значение y ∈ { b, c, r a }, а скорее число N P входных данных, из которых они могут сделать вывод. истинное значение параметра выплаты y . В начале каждой симуляции мы генерируем три функции (для трех параметров выигрыша b , c и r a ), которым игроки должны «учиться» в течение эволюционного времени, если они хотят основывать свои решения. от значений параметров выплат:
В наших симуляциях для каждого параметра выигрыша игроки получают не его фактическое значение y ∈ { b, c, r a }, а скорее число N P входных данных, из которых они могут сделать вывод. истинное значение параметра выплаты y . В начале каждой симуляции мы генерируем три функции (для трех параметров выигрыша b , c и r a ), которым игроки должны «учиться» в течение эволюционного времени, если они хотят основывать свои решения. от значений параметров выплат:
Параметры выбираются случайным образом, единственное ограничение состоит в том, что всегда существует набор, который удовлетворяет (1) для всех возможных значений параметра выплаты y .Число N P входов для каждого параметра выплаты определяет сложность задачи, которая состоит из вывода «истинного» значения параметра выплаты из сенсорных входных данных. Кроме того, для каждой игры мы случайным образом выбираем N S ложных входных данных, на которых игроки также могут основывать свои решения. Всего доверительный управляющий получает 2 N P + N S входных данных, а доверительный управляющий получает 3 N P + N S входных данных, поскольку доверительный управляющий не имеет доступа к автоматическое вознаграждение r a , когда он решает «заплатить, чтобы увидеть» возможное вознаграждение (см. рисунок 1).
Две искусственные нейронные сети (ИНС), доверенный и доверенный, генерируются с одним скрытым слоем из 10 нейронов и со случайно выбранными синаптическими весами. Для каждого вектора входных данных (входные данные для выплат и ложные характеристики отдельной игры) выход доверенного лица представляет собой вероятность p ∈ [0,1], чтобы «заплатить, чтобы увидеть» вознаграждение за эту игру. Обратите внимание, что мы явно моделируем только первое решение доверительного управляющего, которое должно решить, платить ли ему, чтобы увидеть вознаграждение доверительного управляющего (рис. 1).Мы предполагаем, что второе решение, которое состоит в том, чтобы инвестировать или отказаться после отслеживания вознаграждения, оптимизировано естественным отбором и что доверитель инвестирует iff r a + r — c . На выходе доверительного управляющего будет получено вознаграждение в размере рублей за эту игру.
1).Мы предполагаем, что второе решение, которое состоит в том, чтобы инвестировать или отказаться после отслеживания вознаграждения, оптимизировано естественным отбором и что доверитель инвестирует iff r a + r — c . На выходе доверительного управляющего будет получено вознаграждение в размере рублей за эту игру.
Фаза предварительного выбора
Может случиться так, что случайно сгенерированные ИНС отслеживают награды или предоставляют достаточно большое вознаграждение доверительному управляющему в некоторых совместных играх ( r> c ).Это могут быть случаи случайного развития сотрудничества, не обусловленного обобщением, а это именно то, что нас интересует. Чтобы избежать таких ситуаций, мы сначала обучаем ИНС никогда не отслеживать ( p = 0) и предоставлять нулевое вознаграждение в каждые игры ( r = 0).
Фаза выбора
Затем мы обучаем обе ИНС обеспечивать соответственно адаптивную вероятность «заплатить, чтобы увидеть» и адаптивное вознаграждение в каждой игре обучающего набора, как описано в разделе . Скрытые вознаграждения решают проблему начальной загрузки взаимного сотрудничества .Обратите внимание, что ИНС доверительного управляющего не обучается в некоторых играх: играх, в которые доверительный управляющий никогда не инвестирует ( p = 0).
Градиентный спуск
В предыдущих работах уже изучалась эволюция социального поведения с помощью ИНС (Арак и Энквист, 1993; Энквист и Арак, 1993; Джонстон, 1993; Андре и Нольфи, 2016; Дебов и др., 2017). В этих работах используются генетические алгоритмы, вдохновленные эволюцией путем естественного отбора. Однако существует формальная эквивалентность между естественным отбором и простыми формами обучения (Watson and Szathmáry 2016, Zador 2019; Hasson et al.2020). Например, многие методы машинного обучения основаны на градиентном спуске. Этот принцип также используется в классе моделей, которые широко использовались для описания эволюционного процесса: адаптивной динамики (Хофбауэр и Зигмунд, 1990; Гериц и др. , 1998). В случае естественного отбора признаки организмов постепенно развиваются в сторону максимизации приспособленности. В случае градиентного спуска синаптические веса ИНС постепенно изменяются в соответствии с градиентом функции ошибок, которая количественно определяет, насколько точна ИНС.Оба механизма приводят к машинам принятия решений, которые обеспечивают точный вывод для каждого набора входов, с которыми они столкнулись.
, 1998). В случае естественного отбора признаки организмов постепенно развиваются в сторону максимизации приспособленности. В случае градиентного спуска синаптические веса ИНС постепенно изменяются в соответствии с градиентом функции ошибок, которая количественно определяет, насколько точна ИНС.Оба механизма приводят к машинам принятия решений, которые обеспечивают точный вывод для каждого набора входов, с которыми они столкнулись.
В наших симуляциях мы используем простой алгоритм обратного распространения, то есть градиентный спуск, применяемый к ИНС (Goodfellow et al., 2016). Точнее, мы используем мини-пакетный градиентный спуск с размером пакета 50 и скоростью обучения 0,2. На каждой итерации мы вычисляем выходы доверительного управляющего ( p ) и доверительного управляющего ( r ) в 50 случайно выбранных играх.Из них мы выводим их адаптивные значения p * и r * в каждой игре (см. Раздел Эволюционно стабильная стратегия доверенного лица ) и выполняем один шаг обратного распространения обеих ИНС для обновления синаптических весов в направление, которое максимизирует приспособленность, то есть минимизирует разницу между фактическим выходом и его адаптивным значением. Мы используем следующую функцию ошибки для доверенного лица и для попечителя
Обратите внимание, что разница умножается на коэффициент, чтобы учесть тот факт, что в одних играх выбор сильнее, чем в других.Этот метод известен как обучение с учетом затрат (Кукар и Кононенко, 1998). На этапе обновления обратного распространения каждая игра вносит больший или меньший вклад в функцию ошибок в зависимости от своего типа. А именно, относительная экологическая важность сотрудничества α учитывается путем установки K i = 1 для взаимозависимых и кооперативных игр и K i = 1 — α для эгоистичных и расточительных игр. Если α = 0, сила выбора одинакова во всех играх.Напротив, если α = 1, только взаимозависимые и кооперативные игры влияют на приспособленность. Мы всегда предполагаем, что α = 0, за исключением результатов, показанных на рисунке 6. Кроме того, мы предполагаем, что выбор доверительного управляющего для предоставления вознаграждения в игре i линейно увеличивается с вероятностью p i , что трастер «платит, чтобы увидеть» награду в этой игре.
Мы всегда предполагаем, что α = 0, за исключением результатов, показанных на рисунке 6. Кроме того, мы предполагаем, что выбор доверительного управляющего для предоставления вознаграждения в игре i линейно увеличивается с вероятностью p i , что трастер «платит, чтобы увидеть» награду в этой игре.
Для каждой симуляции мы выполняем 10 3 минипакетов для фазы предварительного выбора и 10 6 для выбора.Значения равновесия, показанные на рисунках 3 и 6, представляют собой средние значения за последние 5 × 10 4 шагов, на которых мы убеждаемся, что алгоритм сходится.
Эволюционно устойчивая стратегия доверительного управляющего
Трастер должен решить для каждой игры, хочет ли он отказаться или играть условно, исходя из качества доверительного управляющего. Мы смоделировали это решение с вероятностью p ∈ [0,1], чтобы «заплатить, чтобы увидеть».
Наблюдение за доверительным управляющим в данной игре может быть оправдано только в том случае, если знак чистой выплаты доверительного управляющего в этой игре неуверен (то есть, если r a + r — c иногда бывает положительным и иногда отрицательный в той же игре, в зависимости от качества доверительного управляющего).В противном случае для доверенного лица всегда лучше сэкономить на мониторинге и безоговорочно сыграть или отклонить игру. Если знак r a + r — c неуверен, с другой стороны, оптимальная стратегия доверенного лица менее прямолинейна. Для простоты на протяжении всей статьи мы предполагаем, что в этом случае мониторинг всегда стоит того. То есть мы предполагаем, что взаимодействия продолжительны (или, в более общем смысле, важны с точки зрения отдачи), так что затраты на мониторинг м всегда ниже, чем ожидаемая выгода от полезных взаимодействий и ожидаемая стоимость дорогостоящих взаимодействий.
Следовательно, эволюционно стабильная стратегия (ESS) доверительного управления выглядит следующим образом:
Во всех играх, где r + r a — c всегда отрицательно, трастер должен отклониться ( p * = 0).
Во всех играх, где знак r + r a — c является неопределенным, доверительный управляющий должен заплатить млн за контроль качества доверительного управляющего ( p * = 1), а затем играть в игру iff r + r a — c > 0

Эволюционно стабильная стратегия доверительного управляющего
Доверительный управляющий должен активно выплачивать вознаграждение только тогда, когда оно того стоит, то есть только когда Трастер принимает во внимание ее награду, чтобы принять решение.Соответственно, эволюционно устойчивая стратегия попечителя заключается в следующем:
Во всех играх, где попечитель будет играть условно по знаку r + r a — c , попечитель должен предложить вознаграждение r * = c — r a + ϵ , где ϵ > 0, как раз достаточно, чтобы побудить доверенного лица играть в игру (см. 65).
Во всех играх, где доверительный управляющий в любом случае безоговорочно откажется от участия, решение доверительного собственника является нейтральным, поскольку оно никогда не происходит.Он представляет собой «скрытый» аспект стратегии доверительного управляющего.
Длительная генерализация, вызванная единичным испытанием на инвазивном раке Procambarus clarkii
РЕЗЮМЕ
Поведенческая гибкость позволяет адаптироваться к изменениям окружающей среды, с чем часто сталкиваются инвазивные виды при колонизации новых территорий. Такая гибкость проистекает из набора когнитивных механизмов, среди которых ключевую роль играет обобщение, поскольку оно позволяет переносить прошлые решения для решения аналогичных новых проблем.С помощью парадигмы привыкания мы изучили генерализацию у инвазивных раков Procambarus clarkii . После того, как раки привыкли к своей защитной реакции на конкретную струю воды, мы проверили, переносится ли привыкание на новый тип струи воды. Хотя привыкание не распространялось при первоначальном предъявлении нового стимула, оно неожиданно проявилось через 15 и 45 дней. Отсюда, что примечательно, в P . clarkii , одного представления нового события было достаточно, чтобы запустить длительную форму обобщения обучения на основе предыдущих подобных стимулов, когнитивную способность, которая может обеспечить адаптивные преимущества для этого инвазивного вида.
Хотя привыкание не распространялось при первоначальном предъявлении нового стимула, оно неожиданно проявилось через 15 и 45 дней. Отсюда, что примечательно, в P . clarkii , одного представления нового события было достаточно, чтобы запустить длительную форму обобщения обучения на основе предыдущих подобных стимулов, когнитивную способность, которая может обеспечить адаптивные преимущества для этого инвазивного вида.
Сноски
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих или финансовых интересов.
Вклад авторов
Концептуализация: C.C .; Методология: A.C., C.C.; Формальный анализ: A.D .; Расследование: A.C .; Написание — оригинальная черновик: н.э .; Написание — просмотр и редактирование: A.D., C.C .; Надзор: C.C .; Финансирование: C.C.
Финансирование
Эта работа была частично поддержана Fondo per la Ricerca di Ateneo (FRA) от Университета Триеста до К.C.
Доступность данных
Данные доступны из Open Science Framework (doi: 10.17605 / OSF.IO / FX67G): https://osf.io/fx67g/
- Получено 29 апреля , 2020.
- Принято 8 октября 2020 года.
- © 2020. Опубликовано The Company of Biologies Ltd
От специализации к обобщению: Васер, Николас М., Оллертон, Джефф: 9780226874005: Amazon.com: Книги
.a-tab-content> .a-box-inner {padding-top: 5px; padding-bottom: 5 пикселей; } #mediaTabs_tabSetContainer .a-tab-content {border-radius: 0px; } #mediaTabsHeadings {white-space: nowrap; переполнение: скрыто; } # mediaTabsHeadings.nonJSTabs {white-space: normal; } #mediaTabsHeadings ul. a-tabs {background: # f9f9f9; }
#mediaTabsHeadings .mediaTab_heading .mediaTab_logo {padding-left: 3px; вертикальное выравнивание: базовая линия; }
#mediaTabsHeadings #mediaTabs_tabSet {margin-top: 5px; плыть налево; граница справа: 0 пикселей; }
#mediaTabsHeadings.mediaTab_heading {маржа слева: -1px; }
#mediaTabsHeadings .mediaTab_heading a {color: # 111; граница справа: сплошной 1px #ddd; padding-top: 8 пикселей; padding-bottom: 7 пикселей; }
#mediaTabsHeadings .mediaTab_heading.a-active a {color: # c45500; маржа сверху: -5 пикселей; padding-top: 11 пикселей; граница слева: сплошной 1px #ddd; border-top-width: 3px;}
#mediaTabsHeadings .tabHidden {display: none! important; }
#bookDescription_feature_div {дисплей: встроенный блок; ширина: 100%;}
]]>
a-tabs {background: # f9f9f9; }
#mediaTabsHeadings .mediaTab_heading .mediaTab_logo {padding-left: 3px; вертикальное выравнивание: базовая линия; }
#mediaTabsHeadings #mediaTabs_tabSet {margin-top: 5px; плыть налево; граница справа: 0 пикселей; }
#mediaTabsHeadings.mediaTab_heading {маржа слева: -1px; }
#mediaTabsHeadings .mediaTab_heading a {color: # 111; граница справа: сплошной 1px #ddd; padding-top: 8 пикселей; padding-bottom: 7 пикселей; }
#mediaTabsHeadings .mediaTab_heading.a-active a {color: # c45500; маржа сверху: -5 пикселей; padding-top: 11 пикселей; граница слева: сплошной 1px #ddd; border-top-width: 3px;}
#mediaTabsHeadings .tabHidden {display: none! important; }
#bookDescription_feature_div {дисплей: встроенный блок; ширина: 100%;}
]]>Возможна отправка через 1-2 дня.
Поставляется и продается на Amazon.com.
Доступно по более низкой цене у других продавцов, которые могут не предлагать бесплатную доставку Prime. Доступно по более низкой цене у других продавцов, которые могут не предлагать бесплатную доставку Prime.