Свойства внимания, Переключаемость и объем внимания
Внимание в психологии подразделяется на 5 основных свойств.
Устойчивость.
Это способность человека в течение продолжительного времени удерживать внимание на каком-либо предмете, не отвлекаясь.
Устойчивость внимания связано с различными причинами. Один из факторов — это психологическое состояние человека на данный промежуток времени. Если он перевозбужден, то устойчивость внимания снижена, т.к. происходит сильное переутомления организма.
Индивидуальные физиологические особенности также могут повлиять на устойчивость, например, при слабой нервной системе у человека быстрей снижается устойчивость внимания.
Большую роль играет мотивация. Отсутствие интереса к информации, приводит к снижению устойчивости внимания, человек начинает чаще отвлекаться, ему стоит больших усилий перейти к изучению данного материала. При высокой мотивации может произойти полное «погружение» в изучаемый предмет.
Сосредоточенность.

Сосредоточенность внимания имеет и другое название — концентрация.
При повышенной концентрации, даже громкие звуки, разговор других людей не мешают изучать понравившийся нам материал.
Переключаемость.
Скорость, с которой человек способен перейти с одного предмета или вида деятельности на другой называется переключаемостью. Переключаемость может быть произвольной и непроизвольной.
При произвольном переключении человек сознательно переводит свое внимание.
А при непроизвольном внимании, человек начинает отвлекаться на объект, который его невольно заинтересовал.
Непроизвольное переключение внимания связано в первую очередь с неустойчивостью нервной системы человека, но это не всегда плохо. При непроизвольном переключении возникает временный отдых нашего организма. Происходит восстановление работоспособности нервной системы.
Распределение.
Распределение внимания — это способность выполнять параллельно несколько действий, при этом происходит расширение нашего внимания на несколько объектов.
Внешне это выглядит так, как будто мы одновременно выполняем несколько дел, но это не совсем так. Эта способность связана с быстротой переключения внимания.
Скорость переключения внимания зависит от физиологического и психологического состояния человека. При утомлении возникает сужение распределения внимания.
Объем.
Объем внимания определяет, какое количество информации способен человек сохранить при повышенном внимании.
Средний объем внимания у человека равен плюс/минус 5-7 единиц. Объем внимания отождествляют с кратковременной памятью.
Управление персоналом, образование, личное развитие. Тесты. Внимание. Память. IQ-тесты. Effecton Studio. Эффектон
Особенности внимания
В связи с разнообразием явлений и множеством свойств внимания возникает особая проблема их систематики. Задачи описания свойств и классификации видов внимания решались различными исследователями по-разному. Так, Джемс, классифицируя виды внимания, опирался на данные житейского опыта и самонаблюдения и наиболее существенной считал функцию отбора.
Задачи описания свойств и классификации видов внимания решались различными исследователями по-разному. Так, Джемс, классифицируя виды внимания, опирался на данные житейского опыта и самонаблюдения и наиболее существенной считал функцию отбора.
Давайте вместе понаблюдаем за собой. За кои веки вы выбрались на природу. Вначале все окружающее видится вам чем-то сказочно прекрасным. Потом Вы начинаете замечать деревья, траву, небо, птиц, переключая свое внимание с одного объекта на другой.
Вот Вы выражаете бурный восторг, увидев муравья, который тащит какую-то щепочку — и уже ваше внимание сконцентрировано на нем. Вы долго наблюдаете за его потугами, в этом проявляется устойчивость вашего внимания, потом ему на помощь прибегают его шустрые помощники, хватают щепочку, и Вы уже наблюдаете за каждым из них, распределяя свое внимание. Получается, что у вас одновременно несколько объектов (муравьев) для детального наблюдения — вот и объем внимания.
Таким образом, внимание не столь условно, как можно было бы подумать, и имеет некоторые особенности, которые у разных людей проявляются в разной степени его можно если не ощутить, то как-то измерить, оно имеет объем, концентрацию, переключаемость, устойчивость, распределение. Рассмотрим основные свойства внимания.
Рассмотрим основные свойства внимания.
Свойства внимания
Концентрация внимания
Концентрация внимания (сосредоточенность) — выделение сознанием объекта и направление на него внимания. Роль концентрированного внимания может быть различна. С одной стороны, оно необходимо для более полного и глубокого исследования того или иного объекта, а другой стороны, чрезмерная концентрация внимания приводит к резкому сужению поля внимания, что создает трудности в восприятии других важных объектов.
Устойчивость внимания
Устойчивость внимания — продолжительность времени, в течение которого человек может поддерживать свое внимание на объекте. Она особенно нужна в условиях однообразной и монотонной работы, когда длительное время выполняются сложные, но однотипные действия.
Интенсивность внимания
Опыты показывают, что интенсивное сорокаминутное внимание может сохраняться произвольно без заметного ослабления и непроизвольных переключений, хотя это и довольно утомительно.
Концентрация и устойчивость внимания
Особое значение для достижения успеха в любой деятельности имеет сосредоточенность и устойчивость внимания, которые характеризуют глубину, длительность и интенсивность психической деятельности человека. Именно сосредоточенность и устойчивость отличают людей, страстно увлеченных делом, умеющих ради основного отключиться от многочисленных побочных раздражителей.
Даже при очень устойчивом и сосредоточенном внимании всегда есть кратковременные непроизвольные изменения степени его интенсивности, напряженности. Это колебания внимания.
Как заставить себя несколько раз внимательно прочитать один и тот же текст? Этого можно добиться, если перед каждым повторным чтением ставить новые задачи. Сказать себе: «Первый раз читаю для общего ознакомления, теперь прочту, чтобы усвоить логику доказательств, которые приводит автор, дальше важно понять, как этот материал связан с предыдущим», и т. д. Т.е. надо учиться смотреть даже на знакомые явления с новой точки зрения.
д. Т.е. надо учиться смотреть даже на знакомые явления с новой точки зрения.
Объем внимания
Для некоторых профессий высокая интенсивность и большой объем внимания необходимы почти во все время трудовой деятельности, а двигательные навыки имеют гораздо меньшее значение. Эти профессии относятся к психологии труда к так называемым наблюдательным. Это диспетчеры, операторы аппаратуры и т.д.
Высокая интенсивность концентрированного внимания для других групп профессий нужна лишь в отдельные моменты работы.
Распределение внимания
Распределение — это способность одновременно выполнять несколько действий. Оно зависит от индивидуальных особенностей личности и от профессиональных навыков. Никто не сможет одновременно делать два дела, не умея делать каждое в отдельности.
Оно зависит от индивидуальных особенностей личности и от профессиональных навыков. Никто не сможет одновременно делать два дела, не умея делать каждое в отдельности.
Переключение внимания
Способность человека удерживать в центре внимания определенное число разнородных объектов одновременно позволяет совершать сразу несколько действий, сохраняя их в поле внимания.
Вспомним, например, феноменальные способности Юлия Цезаря, который, согласно преданию, мог одновременно делать семь не связанных между собой дел. Также Наполеон мог одновременно диктовать своим секретарям семь ответственных дипломатических документов.
Но как показывает жизненная практика, человек способен выполнить только один вид сознательной психической деятельности, а субъективное ощущение одновременности выполнения нескольких обязано быстрому последовательному переключению с одной на другую.
Еще В.Вундтом было показано, что человек не может сосредоточиваться на двух одновременно предъявляемых раздражителях. Однако иногда человек действительно способен выполнять одновременно два вида деятельности. На самом деле, в таких случаях один из видов выполняемой деятельности должен быть полностью автоматизирован, и не требовать внимания. Если же это условие не соблюдается, совмещение деятельности невозможно.
Однако иногда человек действительно способен выполнять одновременно два вида деятельности. На самом деле, в таких случаях один из видов выполняемой деятельности должен быть полностью автоматизирован, и не требовать внимания. Если же это условие не соблюдается, совмещение деятельности невозможно.
Распределение и переключение внимания
Большая группа профессий связанная с управлением движущимися механизмами (автомашины, краны, электровозы…) называется в психологии труда водительскими. Для них важны такие качества внимания, как широкое распределение и быстрое переключение, определяющие успешность управления механизмами в условиях многопланового воздействия в условиях внешнего мира.
Физиологический механизм распределения внимания связан с тем, что привычные действия, не вызывающие затруднений вследствие уже выработанных прочных систем временных связей, могут управляться участками коры, находящимися вне очага оптимального возбуждения.
Например, водитель-стажер не может одновременно распределять свое внимание между педалью сцепления, тормозом и в это же время слушать указания инструктора и наблюдать обстановку. В дальнейшем за счет автоматизации отдельных движений он научится распределять свое внимание. Благодаря тестированию при помощи тестов пакета «Внимание», время на обучение автоматизации действий человека уменьшится.
В дальнейшем за счет автоматизации отдельных движений он научится распределять свое внимание. Благодаря тестированию при помощи тестов пакета «Внимание», время на обучение автоматизации действий человека уменьшится.
Динамика любой работы приводит к необходимости постоянно менять объекты, на которые человек обращает внимание. Это выражается в переключении внимания.
Переключение внимания
Переключение — это сознательный перенос внимания с одного объекта на другой. Непроизвольное переключение внимания называется отвлечением внимания.
Физиологически произвольное переключение внимания объясняется перемещением по коре головного мозга участка с оптимальной возбудимостью. Высокая подвижность нервных процессов как индивидуальная черта темперамента позволяет легко и быстро переходить от одного объекта к другому. В таких случаях говорят о подвижном, гибком внимании.
Если же у человека недостаточная подвижность нервных волокон, то этот переход происходит с усилием, трудно и медленно. Значит у человека инертное внимание. Когда у человека совсем плохая переключаемость — это липкое внимание. Иногда плохая переключаемость у человека из-за плохой подготовленности к работе.
Значит у человека инертное внимание. Когда у человека совсем плохая переключаемость — это липкое внимание. Иногда плохая переключаемость у человека из-за плохой подготовленности к работе.
Рассеянность и внимательность
При рассеянности сознание человека не имеет определенной направленности, а переходит с одного предмета на другой, т.е. рассеивается.
Виды рассеянности
Можно выделить два основных вида рассеянности. Первый — результат общей неустойчивости внимания. Им, как правило, отличаются дети младшего возраста. Однако оно может быть и у взрослых в результате слабости нервной системы или большого утомления, недосыпания и т.д. Такой вид рассеянности появляется также при отсутствии привычки работать сосредоточенно.
Второй вид рассеянности имеет совсем иной характер. Он возникает потому, что человек сосредоточен на чем-то одном и поэтому не замечает ничего другого. Такой рассеянностью отличаются люди, увлеченные своим делом.
Внимательность
Если человек привыкает все делать внимательно, то внимание, становясь постоянной особенностью, перерастает во внимательность, которая, как черта личности, имеет большое значение в общем, психологическом облике человека. Тот, кто обладает этим качеством, отличается наблюдательностью, способностью лучше воспринимать окружающее. Внимательный человек реагирует на события быстрее и переживает их часто глубже, отличается большой способностью к обучению.
Тот, кто обладает этим качеством, отличается наблюдательностью, способностью лучше воспринимать окружающее. Внимательный человек реагирует на события быстрее и переживает их часто глубже, отличается большой способностью к обучению.
Внимательность связана с большим развитием свойств внимания: его объема, сосредоточенности, устойчивости, распределения. Обладая этим качеством, человек легко сосредотачивается, у него хорошо развито непроизвольное внимание. Даже при отсутствии интереса к работе внимательный человек может быстро мобилизовать произвольное внимание, заставить себя сосредоточиться на трудном и неинтересном занятии.
Психологическое тестирование при помощи пакета «Внимание» поможет установить свойства внимания каждого человека, выявить отклонения, изучая которые можно направить развитие внимания в нужное русло.
Эксклюзивный материал сайта «www.effecton.ru — психологические тесты и коррекционные программы». Заимствование текста и/или связанных материалов возможно только при наличии прямой и хорошо различимой ссылки на оригинал.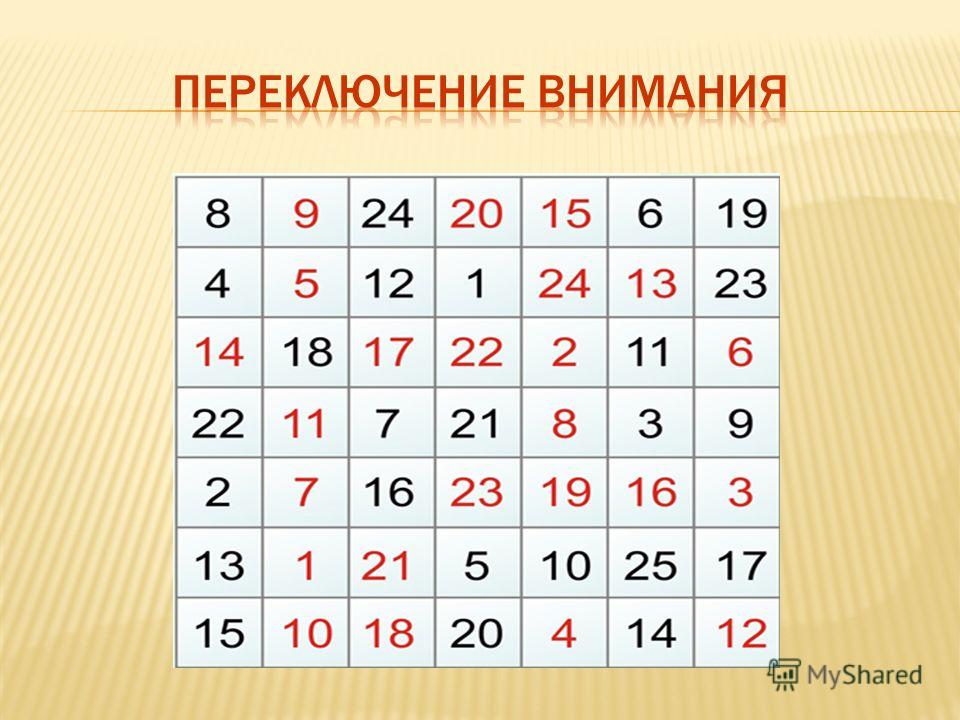 Все права защищены.
Все права защищены.
Особенности свойств внимания: концентрация, устойчивость и переключаемость у младших школьников
В статье рассматриваются свойства внимания у младших школьников и их особенности их проявления в учебной деятельности, представлены результаты эмпирического исследования.
Ключевые слова: внимание, свойства внимания у младших школьников, концентрация внимания, устойчивость внимания, переключаемость внимания.
Внимание имеет огромное значение в жизни человека. Внимание — необходимое условие выполнения любой деятельности. Именно внимание делает все наши психические процессы полноценными, дает возможность воспринимать окружающий нас мир. Внимание является основой успешной познавательной деятельности и принадлежит к числу важных психических процессов, влияющих на успешность обучения младших школьников. Именно от особенностей внимания зависит, может ли ребенок достаточно быстро перейти от одной деятельности к другой, как долго он может работать с одним и тем же материалом, насколько четко ему удается выделять объект и рассматривать, изучать его достаточно долгое время.
Исследования отечественных ученых показали роль и значение внимания и отдельных его свойств на развитие общего интеллекта ребенка. Между тем вопрос о механизмах влияния внимания на общую обучаемость учащихся до сих пор остается предметом острых дискуссий в психологической науке. От его решения зависит выбор стратегии развивающего обучения младших школьников.
Проблема внимания нередко рассматривается лишь в связи с другими психическими функциями: памятью, мышлением, воображением, восприятием. Действительно, проявления внимания нельзя увидеть отдельно от них, в чистом виде. Однако в настоящее время оно все чаще начинает рассматриваться учеными как принципиально важная для жизни и деятельности человека способность, без которой невозможно как его физическое выживание, так и достижение высот творческой деятельности.
Проблемы, связанные с развитием внимания у школьников, сегодня вызывают беспокойство и у педагогов, и у родителей, и у психологов, работающих с детьми. Многие взрослые жалуются на невнимательность детей, их неумение сосредоточиться, сколько либо долго удерживать внимание при решении учебных задач.
Многие взрослые жалуются на невнимательность детей, их неумение сосредоточиться, сколько либо долго удерживать внимание при решении учебных задач.
Учителю важно знать и изучать психологические и физиологические механизмы внимания учеников, так как именно организация внимания детей обеспечивает эффективность осуществления учебно-воспитательного процесса.
У детей младшего школьного возраста регулирующее влияние высших корковых центров постепенно совершенствуется, в связи, с чем и происходят существенные преобразования характеристик внимания, начинается интенсивное развитие его свойств, а именно: увеличивается объем внимания, повышается его устойчивость, переключаемость и распределение. Развитие произвольного внимания младшего школьника является важнейшим приобретением личности на данном этапе онтогенеза. Оно связано с формированием у ребенка волевых качеств и находится во взаимодействии с развитием интеллектуальных операций. Внимание поддерживается активной умственной деятельностью учащегося.
Внимание теснейшим образом связано с эмоционально-волевой сферой личности, что и определяет особенности формирования данного процесса у детей младшего школьного возраста. Индивидуальные особенности личности младших школьников оказывают влияние на характер внимания. У детей сангвинистического темперамента кажущаяся невнимательность проявляется в чрезвычайной активности. Сангвиники подвижны, непоседливы, много успевают сделать во время урока. Флегматики и меланхолики пассивны, вялы, часто кажутся невнимательными. На самом деле они сосредоточены на изучаемом предмете.
Преобладающим видом внимания младшего школьника является непроизвольное. В этом возрасте все еще сильная реакция на все новое, яркое, непривычное. Ребенок еще не умеет в достаточной степени управлять своим вниманием. Это объясняется преобладанием наглядно-образного характера мыслительной деятельности. Произвольное внимание в младшем школьном возрасте развивается вместе с развитием мотивов учения. Возможности волевого регулирования внимания в младшем школьном возрасте ограничены, младший школьник обычно может сосредоточенно работать лишь при наличии близкой мотивации (перспективы заработать высокую отметку, получить похвалу учителя).
Объем внимания младшего школьника меньше, чем у взрослого человека, распределение внимания — гораздо слабее. У учащихся начальных классов невысокая устойчивость внимания, это связано с возрастной слабостью процесса торможения. Первоклассники чаще всего имеют рассеянное и неразвитое внимание. Младшие школьники также не могут быстро переключать свое внимание с одного объекта на другой, такие свойства внимания, как переключение и распределение менее развиты в младшем школьном возрасте. На протяжении школьного возраста они интенсивно развиваются. Со временем внимание младшего школьника приобретает выраженный произвольный, преднамеренный характер.
Вообще, внимание является неразрывной частью познавательных процессов, чувств и воли, оно делает успешным весь процесс обучения. Внимание необходимо воспитывать. Возникая в активной деятельности ребенка, внимание не только вызывается, но и поддерживается рационально организованной деятельностью детей, и прежде всего их активным умственным трудом. Когда говорят о развитии внимания, имеют в виду совершенствование свойств внимания.
Когда говорят о развитии внимания, имеют в виду совершенствование свойств внимания.
Выделяют основные свойства внимания: объем, концентрация, распределение, устойчивость, переключение. Свойства внимания в большей степени независимы друг от друга. Внимание хорошее в одном отношении может быть не очень совершенным в другом. Основные свойства внимания можно развивать.
Организация учителем учебной деятельности детей, основанной на разных формах их мыслительной активности, формирует особую черту личности — внимательность. Подводя итог, можно сказать следующее, что современный учитель обладает всеми необходимыми средствами и возможностями для полноценного развития внимания младших школьников.
Исследованием проблемы внимания занимались выдающиеся ученые: Л. С. Выготский, П. Я. Гальперин, Д. Б. Эльконин, А. В. Петровский, Н. Ф. Добрынин, Н. Н. Ланге, В. Вундт, С. Л. Рубинштейн и другие.
Нами проведено эмпирическое исследование свойств внимания у младших школьников таких как: концентрация, устойчивость и переключаемость. В Таблице 1 представлены средние показатели по испытуемым по методике «Тест Бурдона» корректурная проба (буквенный вариант).
В Таблице 1 представлены средние показатели по испытуемым по методике «Тест Бурдона» корректурная проба (буквенный вариант).
Таблица 1
Средние показатели по методике «Тест Бурдона» концентрация, устойчивость ипереключаемость внимания умладших школьников
Исходя из выше представленных данных, можно констатировать следующее: концентрация внимания у младших школьников имеет относительно низкий уровень, поскольку концентрация внимания зависит от типа нервной системы и с трудом поддается корректировке; в свою очередь, устойчивость внимания имеет достаточно высокий показатель, так как к третьему классу оно уже достаточно хорошо сформировано у детей; переключаемость — находится на среднем уровне. Но наши данные имеют отношение только к данной выборке испытуемых.
Чтобы узнать зависит ли уровень развития свойств внимания от пола, необходимо сравнить между собой результаты исследования свойств внимания по группе мальчиков и группе девочек, применив t-критерий Стьюдента. Расчет t-критерия Стьюдента производится по формуле в программе Excel. В Таблице 2 приведены данные по расчету t-критерия Стьюдента по каждому свойству внимания у девочек и мальчиков.
Расчет t-критерия Стьюдента производится по формуле в программе Excel. В Таблице 2 приведены данные по расчету t-критерия Стьюдента по каждому свойству внимания у девочек и мальчиков.
Таблица 2
Данные расчета t-критерия Стьюдента по каждому свойству внимания удевочек имальчиков
При интерпретации данных расчета t-критерия Стьюдента следует, что статистических значимых различий между группами мальчиков и девочек нет, соответственно уровень развития свойств внимания не зависит от пола.
Внимание зависит, в том числе от потребностей, интересов, направленности, темперамента и волевых качеств младшего школьника, независимо от пола.
Целью данного экспериментального исследования было изучение свойств внимания: концентрация, устойчивость, переключаемость у младших школьников.
Одним из исследуемых нами свойств внимания являлась концентрация. По результатам диагностики мы констатировали достаточно низкий уровень концентрации внимания по группе исследуемых младших школьников. Другие раздражители будут отвлекать учащихся от выполняемого задания. Это может негативно сказываться на их учебной деятельности.
Другие раздражители будут отвлекать учащихся от выполняемого задания. Это может негативно сказываться на их учебной деятельности.
Следующим изучаемым свойством внимания младших школьников явилась устойчивость. По данным исследования — высокий уровень показателя. Это значит, что младшие школьники, принявшие участие в нашем исследовании, способны долгое время удерживать внимание на предложенном задании, доделывать задание до конца, не нарушая инструкции.
Ну и третьим исследуемым нами свойством внимания у младших школьников — переключаемость внимания имеет средний уровень. У исследуемых младших школьников нет особых трудностей при переходе с одного, выполняемого задания к другому. В целом, переключаемость внимания поддается тренировке легче, чем другие свойства.
Таким образом, можно сделать вывод, что свойства внимания взаимосвязаны между собой, и нарушение отдельного свойства может влиять на другие, а может компенсировать его. В то же самое время каждое из свойств можно рассматривать отдельно от других, и при нарушении одного свойства все прочие могут иметь достаточно высокий уровень выраженности.
Чем лучше у учащихся будет развито внимание, тем они будут более успешны в учебной деятельности.
Таким образом, мы видим, что цель проведенного эксперимента достигнута. У детей младшего школьного возраста свойства внимания имеют разную степень развития. Высокий уровень развития одного свойства внимания не означает высокого уровня развития других его свойств.
Литература:
- Веккер Л. М. Психика и реальность: единая теория психических процессов. М.: Смысл, 1998.
- Добрынин Н. Ф. О теории и воспитании внимания. М.: РПО, 1976.
- Дружинин В. Н., Ушаков Д. В. Когнитивная психология. М.: Пер Сэ, 2002.
- Ланге Н. Н. Психический мир. М.: МОДЭК, 1996.
- Макарова К. В., Таллина О. А. Психология человека. М.: Прометей, 2011.
- Рибо Т. А. Психология внимания. М.: УРСС, 2011.
- Шадриков В. Д. Диагностика познавательных способностей. Методика и тесты. М.: Альма Матер, 2009.

- Шадриков В. Д., Мазилов В. А. Общая психология. М.: Юрайт, 2018.

Основные термины (генерируются автоматически): свойство внимания, внимание, младший школьник, младший школьный возраст, школьник, концентрация внимания, расчет t-критерия, ребенок, учебная деятельность, долгое время.
Общая психопатология | Обучение | РОП
В норме внимание помогает индивиду воспринимать наиболее важные для него стимулы, игнорируя неважные, кроме того, внимание дает индивиду способность произвольно направлять и удерживать фокус своего внимания на интересующих его объектах, что позволяет продуктивно решать стоящие перед ним задачи.
Нарушения внимания могут быть достаточно разнообразными. Выделяют нарушения внимания, связанные с изменением состояния бодрствования (в этих случаях может быть нарушено, как произвольное, так и непроизвольное внимание), и нарушения характеристик произвольного внимания (его селективности, устойчивости, переключаемости, объема и пр. ).
).
Невнимание (апрозексия) — этот термин часто применятся сразу по отношению ко всей группе нарушений внимания, однако при более дифференцированном подходе его правильно использовать лишь в случаях общего угнетения процессов внимания, т.е. при отсутствии или затруднении фокусировки внимания как на важных, так и на неважных стимулах; в связи с этим восприятие всех стимулов нарушено, они игнорируются. В основе лежит снижение тонуса нервной системы, уровня бодрствования и готовности действовать. В таких состояниях человек выглядит вялым, недостаточно реагирующим на происходящее вокруг, его внимание трудно или невозможно чем-либо привлечь. Невнимание в той или иной степени выраженности наблюдается при угнетении сознания, интоксикациях седативными веществами, выраженном утомлении, апатии, грубых нарушениях интеллекта, органических поражениях головного мозга определенных локализаций. Например, при оглушении пациент воспринимает лишь малую часть окружающих его стимулов, он вял, аспонтанен, врачу приходится многократно повторять свой вопрос, чтобы пациент мог сфокусировать свое внимание на нем и на попытках на него ответить. Врач из-за чрезмерного утомления и сонливости, например во время ночного дежурства, может упустить признаки болезни у своего пациента, в том числе как важные, так и неважные, допустить ошибку в своих действиях. Водитель в состоянии алкогольного опьянения или под действием седативных препаратов из-за невнимательности может «просмотреть» изменение ситуации на дороге.
Врач из-за чрезмерного утомления и сонливости, например во время ночного дежурства, может упустить признаки болезни у своего пациента, в том числе как важные, так и неважные, допустить ошибку в своих действиях. Водитель в состоянии алкогольного опьянения или под действием седативных препаратов из-за невнимательности может «просмотреть» изменение ситуации на дороге.
Сверхотвлекаемость (гиперметаморфоз, сверхбодрствование) — фокусировка внимания сразу на всех действующих на человека стимулах, как на важных, так и на неважных. В основе — состояние, противоположное невниманию, — повышенная активация нервной системы и «чрезмерный» уровень бодрствования. В результате внимание становится крайне неустойчивым, сверхизменчивым, человек постоянно отвлекается на каждый новый стимул. В выраженных случаях нарушаются последовательность, цельность мыслительной деятельности, вплоть до развития состояния растерянности, недоумения. Наблюдается при маниакальных состояниях, интоксикациях стимуляторами и психодислептиками, иногда при галлюцинаторно-бредовых состояниях, органических поражениях головного мозга определенных локализаций.
Неустойчивость внимания (патологическая отвлекаемость) — неспособность человека удерживать фокус произвольного внимания на нужном ему объекте, в связи с чем страдает продуктивность его деятельности. При слабости произвольного внимания ведущую роль начинает занимать внимание непроизвольное, из-за этого человек постоянно отвлекается на посторонние стимулы, случайно попадающие в поле его внимания, случайные мысли («витает в облаках», «считает ворон»). Выраженность расстройства может в значительной степени варьировать.
Синдром дефицита внимания с гиперактивностью (СДВГ), основу которого составляет неустойчивость произвольного внимания, может проявляться целым спектром относительно негрубых нарушений внимания, отражающихся преимущественно на успеваемости в школе и работоспособности. Необходимо учитывать, что способность к удержанию фокуса произвольного внимания постепенно формируется у ребенка по мере его развития, поэтому для оценки этого нарушения у детей нужно принимать во внимание возрастные нормы.
При некоторых случаях органических поражений головного мозга (особенно дорсальной поверхности лобных долей) неустойчивость внимания может достигать такой степени, что она полностью дезорганизует деятельность взрослого человека.
Больной К. 35 лет несколько лет назад перенес тяжелую травму головного мозга во время погрузочных работ в порту, последние годы находится в психиатрической больнице. В отделении, если кто-то из медсестер громко вызывает любого из пациентов, К. тут же откликается, бросает свои дела и бежит к этой медсестре через весь коридор. Таким же образом встречает каждого, кто входит в отделение, начинает бойко и громко с ним разговаривать, но потом также быстро отходит, обратив внимание на что-то новое. Полноценно побеседовать с лечащим врачом он не в состоянии — ответив на один-два вопроса, он переключается на случайные темы, сам начинает задавать вопросы, а если рядом что-то происходит, теряет всякое внимание к беседе с врачом и, не завершив беседу, уходит в интересующем его направлении, например, посмотреть, как стригут других больных или что принесла процедурная медсестра из аптеки.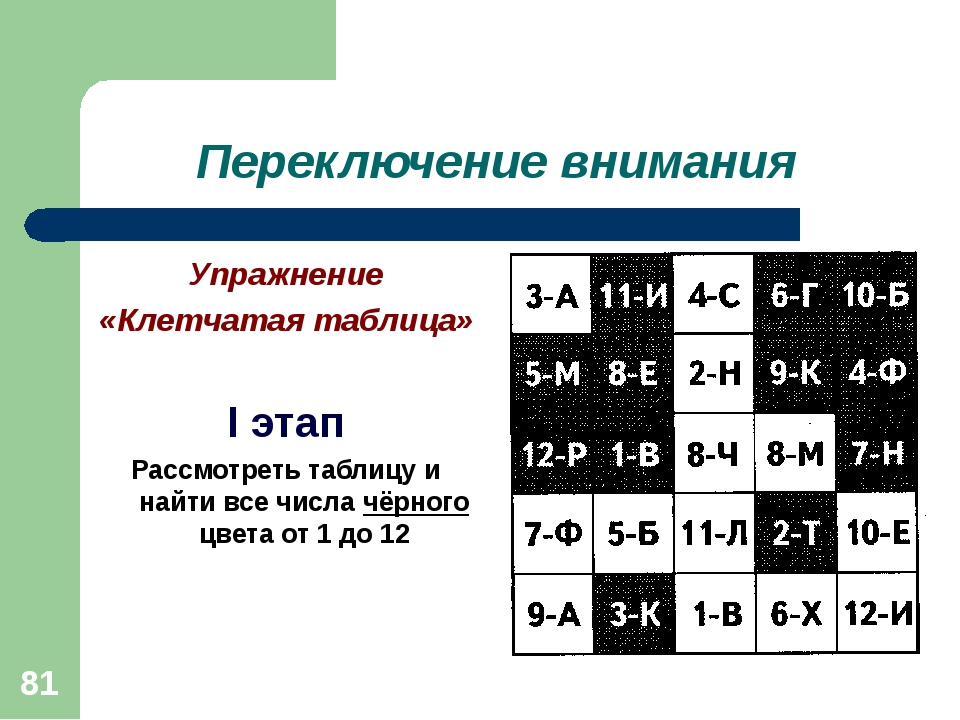
Истощаемость внимания — неспособность человека длительное время удерживать фокус произвольного внимания; первоначально внимание направлено на нужный индивиду объект, но по мере утомления оно все более истощается, а следовательно, непроизвольное внимание выходит на первый план. Является достаточно характерным признаком утомления и астении. Например, студент, утомленный недосыпанием на ночных дежурствах, садится заниматься: какое-то время он сосредоточенно читает учебник, но через несколько страниц замечает, что думает о чем-то постороннем, пытается вспомнить, что он только что читал, но не может. В специальных психологических тестах истощаемость внимания проявляется увеличением количества ошибок в монотонном задании по ходу его выполнения. Истощаемость внимания также наблюдается при синдроме дефицита внимания с гиперактивностью, органическом поражении головного мозга.
Тугоподвижность внимания (вязкость, патологическая прикованность) — трудности с переключением внимания с одной темы или объекта на другую тему или объект. Рассматривается как проявление замедленного мышления, или брадифрении. Отмечается при органических заболеваниях головного мозга, в том числе эпилепсии, сосудистых заболеваниях и пр.
Рассматривается как проявление замедленного мышления, или брадифрении. Отмечается при органических заболеваниях головного мозга, в том числе эпилепсии, сосудистых заболеваниях и пр.
Про чрезмерную прикованность внимания к той или иной теме, от которой человек не может отвлечься и которая заслоняет для него все происходящее вокруг, говорят также, когда описывают патологические суждения (бред, сверхценные идеи и навязчивые явления).
Сужение объема внимания — возможность одновременно удержать в фокусе внимания лишь небольшое количество объектов. Достаточно яркий пример — аффекты (аффективно суженное сознание) — остро развивающееся состояние, когда на фоне сильного эмоционального волнения, происходит резкое ограничение объема воспринимаемой информации (в том числе так называемое туннельное зрение) с сохранением фокуса внимания только на эмоционально значимых объектах; другие объекты, окружающие человека (в том числе те, которые помогли бы найти выход из сложившейся ситуации или дали бы возможность альтернативному пути действий), не воспринимаются (подробнее см. аффекты, аффективно-шоковые реакции).
аффекты, аффективно-шоковые реакции).
В случаях, когда у человека всегда отмечается некоторый недостаток объема и распределения внимания, проявляющиеся в виде невозможности удерживать в фокусе внимания необходимое для выполнения той или иной деятельности количество информации, эти нарушения в настоящее время часто трактуют, как недостаточность объема рабочей памяти. Существует целый ряд подходов к тренингу рабочей памяти, для повышения когнитивной эффективности, включая компьютеризированные варианты, доступные для мобильных устройств (н., n-back test).
Основные свойства внимания
Поскольку наличие внимания означает связь сознания с определенным объектом, его сосредоточенность на нем, прежде всего встает вопрос о степени этой сосредоточенности, т. е. о концентрированности внимания.
Концентрированностъ внимания — в противоположность его распыленности — означает наличие связи с определенным объектом или стороной деятельности и выражает интенсивность этой связи. Концентрация — это сосредоточенность, т. е. центральный факт, в котором выражается внимание. Концентрированность внимания означает, что имеется фокус, в котором собрана психическая или сознательная деятельность.
Концентрация — это сосредоточенность, т. е. центральный факт, в котором выражается внимание. Концентрированность внимания означает, что имеется фокус, в котором собрана психическая или сознательная деятельность.
Наряду с этим пониманием концентрации внимания под концентрированным вниманием часто в психологической литературе понимают внимание интенсивной сосредоточенности на одном или небольшом числе объектов. Концентрированность внимания в таком случае определяется единством двух признаков — интенсивности и узости внимания.
Объединение в понятии концентрации интенсивности и узости внимания исходит из той предпосылки, что интенсивность внимания и его объем обратно пропорциональны друг другу. Эта предпосылка в общем правильна, лишь когда поле внимания состоит из элементов, друг с другом не связанных. Но когда в него включаются смысловые связи, объединяющие различные элементы между собой, расширение поля внимания дополнительным содержанием может не только не снизить концентрированности, но иногда даже повысить ее. Мы потому определяем концентрацию внимания только интенсивностью сосредоточения и не включаем в нее узости внимания. Вопрос об объеме внимания, т. е. количестве однородных предметов, которые охватывает внимание, — особый вопрос.
Мы потому определяем концентрацию внимания только интенсивностью сосредоточения и не включаем в нее узости внимания. Вопрос об объеме внимания, т. е. количестве однородных предметов, которые охватывает внимание, — особый вопрос.
Для определения объема внимания пользовались до сих пор главным образом тахистоскопическим методом. В тахистоскопе на короткое, точно измеряемое время выставлялись подлежащие наблюдению экспонаты, как-то: буквы, цифры, фигуры.
Согласно ряду исследований, обнаруживших при этом существование довольно значительных индивидуальных различий в объеме внимания, объем внимания взрослого человека достигает в среднем примерно до 4—5, максимум 6 объектов; у ребенка он равен в среднем не более 2—3 объектам. Речь при этом идет о числе друг от друга не зависимых, не связанных между собой объектов (чисел, букв и т. п.). Количество находящихся в поле нашего внимания связанных между собой элементов, объединенных в осмысленное целое, может быть много больше. Объем внимания является поэтому изменчивой величиной, зависящей от того, насколько связано между собой то содержание, на котором сосредоточивается внимание, и от умения осмысленно связывать и структурировать материал. При чтении осмысленного текста объем внимания может оказаться существенно отличным от того, который дает его измерение при концентрации на отдельных осмысленно между собой не связанных элементах. Поэтому результаты тахистоскопического изучения внимания на отдельные цифры, буквы, фигуры не могут быть перенесены на объем внимания в естественных условиях восприятия связанного осмысленного материала. В практике, в частности педагогической, школьной, следовало бы, тщательно учитывая доступный учащимся объем внимания, не создавая в этом отношении непосильной перегрузки, расширять объем внимания, систематизируя предъявляемый материал, вскрывая его взаимосвязи, внутренние отношения.
Объем внимания является поэтому изменчивой величиной, зависящей от того, насколько связано между собой то содержание, на котором сосредоточивается внимание, и от умения осмысленно связывать и структурировать материал. При чтении осмысленного текста объем внимания может оказаться существенно отличным от того, который дает его измерение при концентрации на отдельных осмысленно между собой не связанных элементах. Поэтому результаты тахистоскопического изучения внимания на отдельные цифры, буквы, фигуры не могут быть перенесены на объем внимания в естественных условиях восприятия связанного осмысленного материала. В практике, в частности педагогической, школьной, следовало бы, тщательно учитывая доступный учащимся объем внимания, не создавая в этом отношении непосильной перегрузки, расширять объем внимания, систематизируя предъявляемый материал, вскрывая его взаимосвязи, внутренние отношения.
С объемом внимания тесно связана и распределяемость внимания. Говоря об объеме, можно, с одной стороны, подчеркивать ограничение поля внимания. Но оборотной стороной ограничения, поскольку оно не абсолютно, является распределение внимания между тем или иным числом разнородных объектов, одновременно сохраняющихся в центре внимания. При распределении внимания речь, таким образом, идет о возможности не одного, а много-, по крайней мере двухфокального внимания, концентрации его не в одном, а в двух или большем числе различных фокусов. Это дает возможность одновременно совершать несколько рядов действий и следить за несколькими независимыми процессами, не теряя ни одного из них из поля своего внимания. Наполеон мог, как утверждают, одновременно диктовать своим секретарям семь ответственных дипломатических документов. Некоторые шахматисты могут вести одновременно с неослабным вниманием несколько партий. Распределенное внимание является профессионально важным признаком для некоторых профессий, как, например, для текстильщиков, которым приходится одновременно следить за несколькими станками. Распределение внимания очень важно и для педагога, которому нужно держать в поле своего зрения всех учеников в классе.
Но оборотной стороной ограничения, поскольку оно не абсолютно, является распределение внимания между тем или иным числом разнородных объектов, одновременно сохраняющихся в центре внимания. При распределении внимания речь, таким образом, идет о возможности не одного, а много-, по крайней мере двухфокального внимания, концентрации его не в одном, а в двух или большем числе различных фокусов. Это дает возможность одновременно совершать несколько рядов действий и следить за несколькими независимыми процессами, не теряя ни одного из них из поля своего внимания. Наполеон мог, как утверждают, одновременно диктовать своим секретарям семь ответственных дипломатических документов. Некоторые шахматисты могут вести одновременно с неослабным вниманием несколько партий. Распределенное внимание является профессионально важным признаком для некоторых профессий, как, например, для текстильщиков, которым приходится одновременно следить за несколькими станками. Распределение внимания очень важно и для педагога, которому нужно держать в поле своего зрения всех учеников в классе.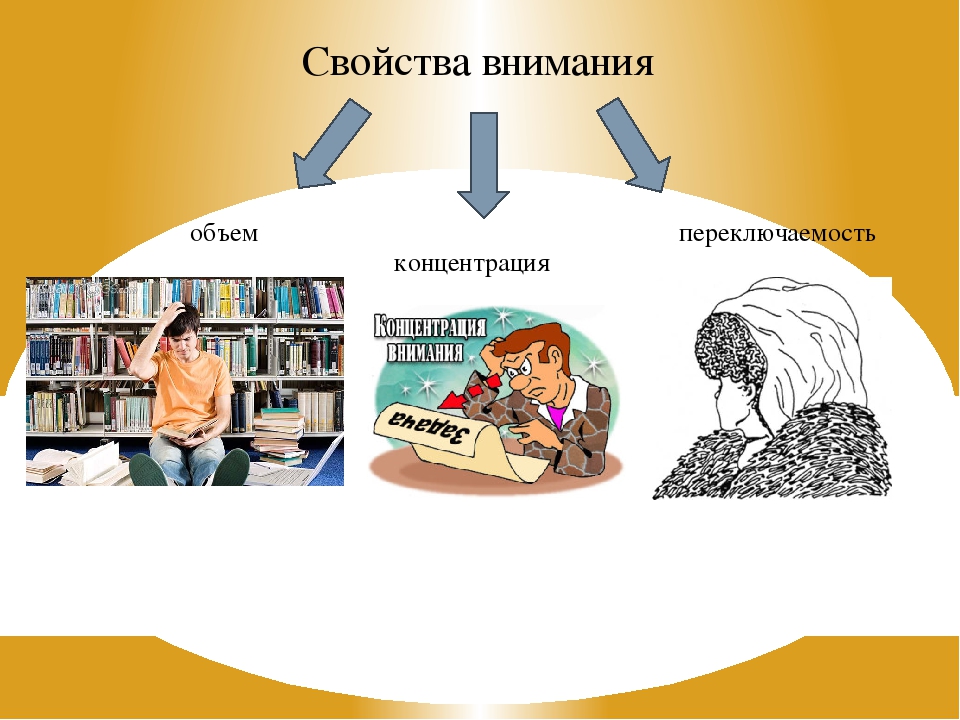
Распределение внимания зависит от ряда условий, прежде всего от того, насколько связаны друг с другом различные объекты и насколько автоматизированы действия, между которыми должно распределяться внимание. Чем теснее связаны объекты и чем значительнее автоматизация, тем легче совершается распределение внимания. Способность к распределению внимания весьма упражняема.
При определении концентрированности и объема внимания необходимо учитывать не только количественные условия. Из качественных моментов, в частности, один играет особенно значительную роль: связность смыслового содержания. Внимание — как и память — подчиняется различным законам, в зависимости от того, на каком материале оно осуществляется. Очень рельефно это сказывается на устойчивости внимания.
Устойчивость внимания определяется длительностью, в течение которой сохраняется концентрация внимания, т. е. его временной экстенсивностью. Экспериментальное исследование показало, что внимание первично подвержено периодическим непроизвольным колебаниям. Периоды колебаний внимания по данным ряда прежних исследований, в частности Н. Ланге, равны обычно 2—3 сек., доходя максимум до 12 сек. К колебаниям внимания относились, во-первых, колебания сенсорной ясности. Так, часы, которые держат неподвижно на одном и том же расстоянии от испытуемого, кажутся ему, если он их не видит, то приближающимися, то удаляющимися, в силу того, что он то более, то менее явственно слышит их биение.
Периоды колебаний внимания по данным ряда прежних исследований, в частности Н. Ланге, равны обычно 2—3 сек., доходя максимум до 12 сек. К колебаниям внимания относились, во-первых, колебания сенсорной ясности. Так, часы, которые держат неподвижно на одном и том же расстоянии от испытуемого, кажутся ему, если он их не видит, то приближающимися, то удаляющимися, в силу того, что он то более, то менее явственно слышит их биение.
Эти и подобные им случаи колебания сенсорной ясности, очевидно, непосредственно связаны с утомлением и адаптацией органов чувств. Иной характер носят колебания внимания, сказывающиеся при наблюдении многозначных фигур; в них попеременно то одна, то другая часть выступает как фигура: глаз соскальзывает с одного поля на другое. В этом можно убедиться если посмотреть на рисунок, в котором мы поочередно видим то вазу, то два профиля. Такой же эффект дает изображение усеченной пирамиды, стоит более длительное время на нее посмотреть, чтобы убедиться в том, что усеченное основание то выступает вперед, то отступает назад.
Однако традиционная трактовка проблемы устойчивости внимания, связанная с установлением периодических его колебаний, требует некоторой ревизии.
Положение с этой проблемой аналогично тому, какое создалось в психологии памяти в связи с установленной Эббингаузом и его последователями кривой забывания. Учебная работа была бы бесплодным, сизифовым трудом, если бы кривая Эббингауза отражала общие закономерности забывания всякого материала. Учебная и производственная работа была бы вообще невозможна, если бы пределы устойчивости внимания определялись периодами, установленными в опытах с элементарными сенсорными раздражителями. Но в действительности такие малые периоды колебания внимания, очевидно, ни в коем случае не составляют всеобщую закономерность. Об этом свидетельствуют наблюдения на каждом шагу. Очевидно, проблема устойчивости внимания должна быть поставлена и разработана заново. При этом существенно не столько экспериментально установить собственно очевидный факт значительно большей устойчивости внимания, сколько вскрыть конкретные условия, которыми объясняются частые периодические колебания в одних случаях, значительная устойчивость — в других.
Наша гипотеза заключается в следующем: наиболее существенным условием устойчивости внимания является возможность раскрывать в том предмете, на котором оно сосредоточено, новые стороны и связи. Там, где в связи с поставленной перед собой задачей мы, сосредоточиваясь на каком-нибудь предмете, можем развернуть данное в восприятии или мышлении содержание, раскрывая в нем новые аспекты в их взаимосвязях и взаимопереходах, внимание может очень длительное время оставаться устойчивым. Там, где сознание упирается как бы в тупик, в разрозненное, скудное содержание, не открывающее возможности для дальнейшего развития, движения, перехода к другим его сторонам, углубления в него, там создаются предпосылки для легкой отвлекаемости и неизбежно наступают колебания внимания.
Подтверждение этого положения имеется еще в одном наблюдении Гельмгольца. Изучая борьбу двух полей зрения (см. рис.), Гельмгольц отметил замечательный факт, в котором заключается ключ для объяснения устойчивости внимания, несмотря на периодические колебания сенсорных установок. «Я чувствую, — пишет Гельмгольц, — что могу направлять внимание произвольно то на одну, то на другую систему линий и что в таком случае некоторое время только одна эта система сознается мною, между тем как другая совершенно ускользает от моего внимания. Это бывает, например, в том случае, если я попытаюсь сосчитать число линий в той или другой системе. Крайне трудно бывает надолго приковать внимание к одной какой-нибудь системе линий, если только мы не связываем предмета нашего внимания с какими-нибудь особенными целями, которые постоянно обновляли бы активность нашего внимания. Так поступаем мы, задаваясь целью сосчитать линии, сравнить их размеры и т. п. Внимание, предоставленное самому себе, обнаруживает естественную наклонность переходить от одного нового впечатления к другому; как только его объект теряет свой интерес, не доставляя никаких новых впечатлений, внимание, вопреки нашей воле, переходит на что-нибудь другое. Если мы хотим сосредоточить наше внимание на определенном объекте, то нам необходимо постоянно открывать в нем все новые и новые стороны, в особенности когда какой-нибудь посторонний импульс отвлекает нас в сторону» (см.
«Я чувствую, — пишет Гельмгольц, — что могу направлять внимание произвольно то на одну, то на другую систему линий и что в таком случае некоторое время только одна эта система сознается мною, между тем как другая совершенно ускользает от моего внимания. Это бывает, например, в том случае, если я попытаюсь сосчитать число линий в той или другой системе. Крайне трудно бывает надолго приковать внимание к одной какой-нибудь системе линий, если только мы не связываем предмета нашего внимания с какими-нибудь особенными целями, которые постоянно обновляли бы активность нашего внимания. Так поступаем мы, задаваясь целью сосчитать линии, сравнить их размеры и т. п. Внимание, предоставленное самому себе, обнаруживает естественную наклонность переходить от одного нового впечатления к другому; как только его объект теряет свой интерес, не доставляя никаких новых впечатлений, внимание, вопреки нашей воле, переходит на что-нибудь другое. Если мы хотим сосредоточить наше внимание на определенном объекте, то нам необходимо постоянно открывать в нем все новые и новые стороны, в особенности когда какой-нибудь посторонний импульс отвлекает нас в сторону» (см. рис. выше, на этой стр.). Эти наблюдения Гельмгольца вскрывают самые существенные условия устойчивости внимания. Наше внимание становится менее подверженным колебаниям, более устойчивым, когда мы включаемся в разрешение определенных задач, в интеллектуальных операциях раскрываем новое содержание в предмете нашего восприятия или нашей мысли. Сосредоточение внимания — это не остановка мыслей на одной точке, а их движение в едином направлении. Для того чтобы внимание к какому-нибудь предмету поддерживалось, его осознание должно быть динамическим процессом. Предмет должен на наших глазах развиваться, обнаруживать перед нами все новое содержание. Лишь изменяющееся и обновляющееся содержание способно поддерживать внимание. Однообразие притупляет внимание, монотонность угашает его.
рис. выше, на этой стр.). Эти наблюдения Гельмгольца вскрывают самые существенные условия устойчивости внимания. Наше внимание становится менее подверженным колебаниям, более устойчивым, когда мы включаемся в разрешение определенных задач, в интеллектуальных операциях раскрываем новое содержание в предмете нашего восприятия или нашей мысли. Сосредоточение внимания — это не остановка мыслей на одной точке, а их движение в едином направлении. Для того чтобы внимание к какому-нибудь предмету поддерживалось, его осознание должно быть динамическим процессом. Предмет должен на наших глазах развиваться, обнаруживать перед нами все новое содержание. Лишь изменяющееся и обновляющееся содержание способно поддерживать внимание. Однообразие притупляет внимание, монотонность угашает его.
На вопрос о том, благодаря чему ему удалось прийти к открытию законов тяготения, Ньютон ответил: «Благодаря тому, что я непрестанно думал об этом вопросе». Ссылаясь на эти слова Ньютона, Кювье определяет гений как неустанное внимание. Основание гениальности Ньютона он видит в устойчивости его внимания. Но обратная зависимость более существенна. Богатство и содержательность его ума, открывавшего в предмете его мысли все новые стороны и зависимости, было, очевидно, существенным условием устойчивости его внимания. Если бы мысль Ньютона при размышлении о тяготении уперлась в одну неподвижную точку, будучи не в силах развернуть этот вопрос, раскрывая в нем новые перспективы, его внимание быстро иссякло бы.
Основание гениальности Ньютона он видит в устойчивости его внимания. Но обратная зависимость более существенна. Богатство и содержательность его ума, открывавшего в предмете его мысли все новые стороны и зависимости, было, очевидно, существенным условием устойчивости его внимания. Если бы мысль Ньютона при размышлении о тяготении уперлась в одну неподвижную точку, будучи не в силах развернуть этот вопрос, раскрывая в нем новые перспективы, его внимание быстро иссякло бы.
Но если бы мысль лишь переходила с одного содержания на другое, можно было бы скорее говорить о рассеянности, чем о сосредоточенности внимания. Для наличия устойчивого внимания необходимо, очевидно, чтобы изменяющееся содержание было объединено совокупностью отношений в одно единство. Тогда, переходя от одного содержания к другому, оно остается сосредоточенным на одном предмете. Единство предметной отнесенности соединяется с многообразием предметного содержания. Устойчивое внимание — это форма предметного сознания. Оно предполагает единство предметной отнесенности многообразного содержания. Таким образом, осмысленная связанность, объединяющая многообразное, динамическое содержание в более или менее стройную систему, сосредоточенную вокруг одного центра, отнесенную к одному предмету, составляет основную предпосылку устойчивого внимания.
Оно предполагает единство предметной отнесенности многообразного содержания. Таким образом, осмысленная связанность, объединяющая многообразное, динамическое содержание в более или менее стройную систему, сосредоточенную вокруг одного центра, отнесенную к одному предмету, составляет основную предпосылку устойчивого внимания.
Если бы внимание при всех условиях было подвержено таким колебаниям, какие имеют место, когда нам даны разрозненные и скудные по содержанию чувственные данные, никакая эффективная умственная работа не была бы возможна. Но оказывается, что самое включение умственной деятельности, раскрывающей в предметах новые стороны и связи, изменяет закономерности этого процесса и создает условия для устойчивости внимания. Устойчивость внимания, будучи условием продуктивной умственной деятельности, является в известной мере и ее следствием.
Осмысленное овладение материалом, раскрывающее посредством анализа и синтеза систематизацию материала и т. д., внутренние связи четко расчлененного содержания, существенно содействует высшим проявлениям внимания.
д., внутренние связи четко расчлененного содержания, существенно содействует высшим проявлениям внимания.
Устойчивость внимания зависит, конечно, помимо того, от целого ряда условий. К числу их относятся: особенности материала, степень его трудности, зна-комости, понятности, отношение к нему со стороны субъекта — степени его интереса к данному материалу и, наконец, индивидуальные особенности личности. Среди последних существенна прежде всего способность посредством сознательного волевого усилия длительно поддерживать свое внимание на определенном уровне, даже если то содержание, на которое оно направлено, не представляет непосредственного интереса, и сохранение его в центре внимания сопряжено с определенными трудностями.
Устойчивость внимания не означает его неподвижности, она не исключает его переключаемости. Переключаемость внимания заключается в способности быстро выключаться из одних установок и включаться в новые, соответствующие изменившимся условиям. Способность к переключению означает гибкость внимания — весьма важное и часто очень нужное качество.
Способность к переключению означает гибкость внимания — весьма важное и часто очень нужное качество.
Переключаемость, как и устойчивость, и объем внимания, и как внимание в целом, не является какой-то самодовлеющей функцией. Она — сторона сложной и многообразно обусловленной сознательной деятельности, в отличие от рассеяния или блуждания ни на чем не концентрированного внимания и от внимания неустойчивого, попросту неспособного длительно удержаться на одном объекте. Переключаемость означает сознательное и осмысленное перемещение внимания с одного объекта на другой. В таком случае очевидно, что переключаемость внимания в сколько-нибудь сложной и быстро изменяющейся ситуации означает способность быстро ориентироваться в ситуации и определить или учесть изменяющуюся значимость различных в нее включающихся элементов.
Легкость переключения у разных людей различна: одни — с легкой переключаемостью — легко и быстро переходят от одной работы к другой; у других «вхождение» в новую работу является трудной операцией, требующей более или менее длительного времени и значительных усилий. Легкая или затруднительная переключаемость зависит от целого ряда условий. К числу их относятся соотношение между содержанием предшествующей и последующей деятельности и отношение субъекта к каждой из них: чем интереснее предшествующая и менее интересна последующая деятельность, тем, очевидно, труднее переключение; и оно тем легче, чем выраженное обратное соотношение между ними. Известную роль в быстроте переключения играют и индивидуальные особенности субъекта, в частности его темперамент. Переключаемость внимания принадлежит к числу свойств, допускающих значительное развитие в результате упражнения. Рассеянность в житейском смысле слова является по преимуществу плохой переключаемостью. Имеется бесчисленное множество более или менее достоверных анекдотов о рассеянности ученых. Тип рассеянного профессора не сходит со страниц юмористических журналов. Однако, вопреки прочно укоренившемуся в обывательском понимании представлению, «рассеянность» ученых является, наоборот, выражением максимальной собранности и сосредоточенности; но только сосредоточены они на основном предмете своих мыслей.
Легкая или затруднительная переключаемость зависит от целого ряда условий. К числу их относятся соотношение между содержанием предшествующей и последующей деятельности и отношение субъекта к каждой из них: чем интереснее предшествующая и менее интересна последующая деятельность, тем, очевидно, труднее переключение; и оно тем легче, чем выраженное обратное соотношение между ними. Известную роль в быстроте переключения играют и индивидуальные особенности субъекта, в частности его темперамент. Переключаемость внимания принадлежит к числу свойств, допускающих значительное развитие в результате упражнения. Рассеянность в житейском смысле слова является по преимуществу плохой переключаемостью. Имеется бесчисленное множество более или менее достоверных анекдотов о рассеянности ученых. Тип рассеянного профессора не сходит со страниц юмористических журналов. Однако, вопреки прочно укоренившемуся в обывательском понимании представлению, «рассеянность» ученых является, наоборот, выражением максимальной собранности и сосредоточенности; но только сосредоточены они на основном предмете своих мыслей. Поэтому при столкновении с рядом житейских мелочей они могут оказаться в том смешном положении, которое живописуют анекдоты. Для того чтобы уяснить себе наличие сосредоточенности у «рассеянного» ученого, достаточно сравнить его внимание с вниманием ребенка, который выпускает из рук только что привлекшую его игрушку, когда ему показывают другую; каждое новое впечатление отвлекает его внимание от предыдущего; удержать в поле своего сознания оба он не в состоянии. Здесь отсутствуют и концентрированность, и распределяемость внимания. В поведении рассеянного ученого также обнаруживается дефект внимания, но он заключается, очевидно, не в легкой отвлекаемости, так как его внимание, наоборот, очень сосредоточено, а в слабой переключаемости. Рассеянность в обычном смысле слова обусловлена двумя различными механизмами — сильной отвлекаемостью и слабой переключаемостью.
Поэтому при столкновении с рядом житейских мелочей они могут оказаться в том смешном положении, которое живописуют анекдоты. Для того чтобы уяснить себе наличие сосредоточенности у «рассеянного» ученого, достаточно сравнить его внимание с вниманием ребенка, который выпускает из рук только что привлекшую его игрушку, когда ему показывают другую; каждое новое впечатление отвлекает его внимание от предыдущего; удержать в поле своего сознания оба он не в состоянии. Здесь отсутствуют и концентрированность, и распределяемость внимания. В поведении рассеянного ученого также обнаруживается дефект внимания, но он заключается, очевидно, не в легкой отвлекаемости, так как его внимание, наоборот, очень сосредоточено, а в слабой переключаемости. Рассеянность в обычном смысле слова обусловлена двумя различными механизмами — сильной отвлекаемостью и слабой переключаемостью.
Различные свойства внимания — его концентрация, объем и распределяемость, переключаемость и устойчивость — в значительной мере независимы друг от друга: внимание хорошее в одном отношении может быть не столь совершенным в другом. Так, например, высокая концентрация внимания может, как об этом свидетельствует пресловутая рассеянность ученых, соединяться со слабой переключаемостью.
Так, например, высокая концентрация внимания может, как об этом свидетельствует пресловутая рассеянность ученых, соединяться со слабой переключаемостью.
Мы охарактеризовали внимание как проявление избирательной направленности психической деятельности, как выражение избирательного характера процессов сознания. Можно было бы к этому прибавить, что внимание выражает не только как бы объем сознания, поскольку в нем проявляется избирательный характер сознания, но и его уровень — в смысле степени интенсивности, яркости.
Внимание неразрывно связано с сознанием в целом. Оно, поэтому, естественно, связано со всеми сторонами сознания. Действительно, роль эмоциональных факторов ярко сказывается в особенно существенной для внимания зависимости его от интереса. Значение мыслительных процессов, особенно в отношении объема внимания, а также его устойчивости, была уже отмечена. Роль волы находит себе непосредственное выражение в факте произвольного внимания.
Диагностика внимания
Один из самых частых запросов родителей на консультации у психолога связан с диагностикой и коррекцией внимания у детей. При этом, жалобы родителей весьма разнообразны: ребенок невнимателен на уроках, часто «летает в облаках, с трудом сосредотачивается на задании, легко отвлекается, делает «глупые» ошибки на письме, при выполнении счетных операций не замечает когда сбивается и получает неверный ответ. Что же значит быть внимательным?
Внимание — это важнейшее качество, которое характеризует процесс отбора нужной информации и отбрасывания лишней. Дело в том, что в человеческий мозг ежесекундно поступают тысячи сигналов из внешнего мира. Если бы не существовало внимания (своеобразного фильтра), то наш мозг не смог бы избежать перегрузки. Внимание обладает определенными свойствами (характеристиками): объемом, устойчивостью, концентрацией, избирательностью, распределением, переключаемостью и произвольностью.
Устойчивость — это длительность привлечения внимания к одному и тому же объекту или к одной и той же задаче.
Объем внимания — количество одновременно отчетливо осознаваемых объектов — может колебаться от 5 до 7 объектов.
Распределение внимание— способность человека удерживать в центре внимания несколько разнородных объектов одновременно. Историки любят рассказывать о феноменальных способностях Юлия Цезаря, Наполеона, которые по приданиям, могли выполнять семь не связанных между собой дел. Их внимание, безусловно, было очень развито, но, кроме того, часть действий была отработана до автоматизма, т. е. не требовала к себе внимания. Таким образом, распределяемость внимания является обратной стороной его переключаемости.
Переключаемость внимания определяется скоростью перехода от одного вида деятельности к другому.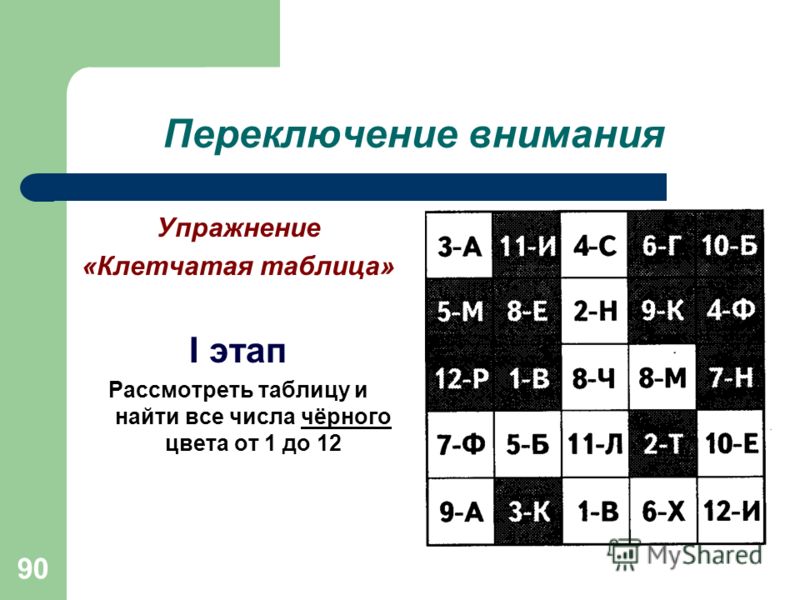 Это свойство теснейшим образом связано с концентрацией внимания, которое можно рассматривать как степень погружения человека в проблему.
Это свойство теснейшим образом связано с концентрацией внимания, которое можно рассматривать как степень погружения человека в проблему.
Избирательность внимания— это способность к сосредоточению на наиболее важных объектах.
Высокий уровень развития основных свойств внимания приводит к появлению произвольного внимания.
Нарушение каждого из перечисленных свойств приводит к отклонениям в поведении и деятельности ребенка. К. Д. Ушинский отмечал огромную роль внимания в психической деятельности: «…внимание есть именно та дверь, через которую проходит все, что только входит в душу человека из внешнего мира». Легко представить, что если эта «дверь» закрыта, то ни о памяти, ни о мышлении, ни о каких других процессах не может быть и речи. Поэтому и любое обучение начинается прежде всего с внимания. Этот процесс является основой любой интеллектуальной деятельности. Своего особого содержания внимание не имеет, но является стороной всех познавательных процессов, проявляясь внутри восприятия, мышления, памяти и др.
Каждая из перечисленных характеристик при диагностике может рассматриваться отдельно и вместе с тем, как частная оценка внимания в целом. Для диагностики внимания предлагаются различные тесты и методики. Наиболее известные и популярные из них, это: «Найди и вычеркни», «Проставь значки», «Запомни и расставь точки», «Переплетенные линии», «Круги», «Корректурная проба», Тест Тулуз-Пьерона, Тест «Кольца Ландольта», «Найди отличия», Тест Мюнстерберга (модификации), Методика «Красно-черная таблица», Таблицы Шульте. Из полученных диагностических данных можно вывести общую, интегральную оценку уровня развития активного внимания.
Особое место в диагностике внимания у детей занимает младший школьный возраст, т.к. именно в этом возрасте формируются многие познавательные процессы. В этот период необходимо более внимательно присматриваться к развитию детей, вовремя отслеживать особенности усвоения ими учебного материала, выявлять те свойства внимания, которые имеют низкие показатели.
СПИСОК ИСПОЛЬЗУЕМОЙ ЛИТЕРАТУРЫ:
1. Л.П. Баданина «Психология познавательных процессов».-М. :Флинта: МПСИ, 2008.
2. С.В. Курдюкова, А.В. Сунцова «Развиваем внимание».- Эксмо, 2010.
3. Л.В. Черемошкина «Развитие внимания детей».- Академия развития, 1997
Педагог-психолог Кривуля А.П.
Персональный сайт автора
Дата публикации — 22.11.2019
Устойчивость внимания: меньше, чем у золотой рыбки?
- Саймон Мейбин
- Всемирная служба Би-би-си, программа «More or Less»
Автор фото, Getty Images
Вы наверняка даже не сможете дочитать до конца эту статью, ведь все знают, что устойчивость внимания современного человека становится все короче — это же так естественно! Или нет?
Трудно фокусироваться на чем-то одном в нынешней действительности, состоящей из соцсетей, смартфонов и гипертекстовых ссылок, вторгающихся именно в тот момент, как вы что-то читаете.
Статистика это тоже подтверждает.
По данным различных исследований, концентрация нашего внимания сократилась с 12 секунд в 2000 году до 8 секунд в настоящий момент — то есть стала меньше, чем у среднестатистической аквариумной рыбки, которая в состоянии поддерживать свое внимание в течение 9 секунд.
Но если вы обратите внимание на то, откуда поступают эти цифры, картина оказывается не такой простой.
Все эти утверждения основаны на отчете, подготовленном в 2015 году группой, изучавшей потребительские предпочтения для канадского отделения компании Microsoft, на основании опроса 2000 канадцев и исследования активности головного мозга у 112 человек во время того, как те занимались разнообразными делами.
Те цифры, которые все издания потом подхватили — о нашей сокращающейся устойчивости внимания, — на самом деле не были получены во время исследования Microsoft. Они действительно впервые появились в этом отчете, но были взяты из другого источника — Statistic Brain.
Автор фото, iStock
Быстрый поиск в Google выводит нас на первоисточник. Видно, что вебсайт Statistic Brain тоже выглядит вполне заслуживающим доверия. Там даже говорится, что авторы сайта «любят цифры, их чистоту и то, о чем они говорят» — как раз с такими людьми мы в программе More or Less и любим иметь дело.
И в качестве доказательства любители цифр с сайта Statistic Brain приводят источники всех своих цифр — откуда они взяты. Однако источники эти своей туманностью только раздражение вызывают.
А когда мы связались с упомянутыми источниками — Национальным центром по биотехнологической информации в Национальной библиотеке США по медицине, а также с Associated Press — никто не смог предоставить никакого свидетельства проведенных исследований, которые бы подтверждали указанные цифры.
Попытки связаться с самим сайтом Statistic Brain тоже ни к чему не привели.
Я поговорил с несколькими людьми, которые посвятили свою жизнь изучению проблем человеческого внимания, но они также не могли понять, откуда появились эти данные.
Автор фото, Getty Images
Подпись к фото,Профессор Бриггс давно изучает то, как выполнение нескольких дел сразу влияет на концентрацию внимания — например, когда человек за рулем пользуется мобильным телефоном
Все зависит от выполняемой задачи
На самом деле, по мнению этих специалистов, утверждения о том, что устойчивость нашего внимания сокращается, совершенно не верны.
«Я совершенно так не думаю, — сказала Джемма Бриггс, доктор психологических наук, преподающая в британском Открытом университете (Open University). — Просто потому, что я не думаю, что психологи или люди, которые интересуются этой проблемой, будут стараться замерять и подсчитывать это таким вот способом».
Джемма изучает внимание у водителей и очевидцев преступления и говорит, что искать среднестатистическую величину «устойчивости внимания» бессмысленно.
«Все зависит от того, чем вы занимаетесь. То, насколько мы концентрируем внимание на выполнении задачи, варьируется от того, что требует от нас эта задача», — говорит она.
Был проведен ряд исследований, когда изучалось как раз, что происходит с концентрацией внимания во время выполнения одной какой-то задачи, например, прослушивания лекции.
Однако предполагать, что существует какое-то определенное количество времени, в течение которого люди обычно сосредотачивают внимание даже на одном каком-то деле, тоже не совсем верно.
«То, как мы распределяем свое внимание на различные задачи, зависит очень во многом от того, что каждый человек привносит в эту конкретную ситуацию», — объясняет Бриггс.
«У нас имеется множество разной информации по поводу того, что обычно случается в каждой конкретной ситуации, что мы можем ожидать от нее. И вот эти наши ожидания и то, что мы испытываем — все это непосредственно сформировывает наше восприятие [ситуации] и то, как мы оцениваем получаемую информацию в каждый конкретный момент», — говорит эксперт.
Высказываются мнения и по поводу того, что все время сокращающиеся по длительности кадры в фильмах демонстрируют как раз то, что концентрация внимания уменьшается. Однако ученый, исследовавший это явление, говорит, что это всего лишь показывает, что кинорежиссеры стали более искусно манипулировать нашим вниманием.
Однако ученый, исследовавший это явление, говорит, что это всего лишь показывает, что кинорежиссеры стали более искусно манипулировать нашим вниманием.
Автор фото, Getty Images
Подпись к фото,У аквариумных рыбок не такая уж короткая память, и они могут обучаться
Интересная ирония
Кое-что еще смущает во всей этой статистике по поводу концентрации внимания.
Оказывается, нет никаких свидетельств того, что у аквариумных рыбок, или каких-либо еще, крайне недлинная устойчивость внимания или память — вопреки тому, что говорится в поп-культуре по этому поводу.
Я поговорил с профессором Фелисити Хантингфорд, которая почти 50 лет изучает поведение рыб и только что прочитала курс лекций на тему «Насколько умны рыбы?»
«Аквариумные рыбки могут обучаться всему тому, чему можно обучить животных или птиц, — говорит профессор. — Они стали модельной системой для изучения процесса обучения и формирования памяти именно потому, что у них есть память и они могут обучаться».
По ее словам, существует в буквальном смысле сотни научных работ, написанных за десятки лет, по поводу обучаемости аквариумных рыбок и их памяти. Я нашел отсылку к исследованию о природе памяти рыб, датируемую 1908 годом.
«То, что тот самый биологический вид, который изучают нейропсихологи и другие группы ученых в качестве модели для понимания формирования памяти, получил такую репутацию — я думаю, в этом заключается любопытная ирония», — говорит профессор Хантингфорд.
Итак, у аквариумной рыбки не короткая устойчивость внимания или память. Как и нет свидетельств тому, что человеческая концентрация внимания сокращается.
Программа More or Less выходит на Всемирной службе Би-би-си по пятницам.
определение переключаемого The Free Dictionary
Резюме: TEHRAN (FNA) — Исследователи описывают первое наблюдение самородного сегнетоэлектрического металла: самосодержащий металл с бистабильными и электрически переключаемыми состояниями спонтанной поляризации — отличительная черта сегнетоэлектричества. Измеряя ширину всего 45 мм, samosPRO COMPACT обеспечивает до 20 безопасных входов и 8 безопасных выходов с использованием 4 переключаемых безопасных входов / выходов, система может быть расширена дополнительными модулями до 116 безопасных входов и 56 безопасных выходов и любого промежуточного числа.Функция переключения черного цвета доступна в качестве обновления как для новых, так и для существующих клиентов Pro8432WT. Новый Street Twin теперь имеет два режима езды — дождь и дорога, а также переключаемый контроль тяги с муфтой помощи крутящего момента. D25 оснащен независимыми поворотными инструментами с регулируемым углом к вспомогательному шпинделю, 3 оси Y и 3 оси Z, ось B для обработки спереди и сзади, до 59 инструментов, возможность переключения для использования с направляющей втулкой или без нее, система ЧПУ нового поколения с сенсорным экраном и различные интерфейсы выбор (USB-память, SD-карта, RS232, Ethernet).Новая программа клеточной терапии Calibr, возглавляемая Тревисом Янгом, доктором философии, директором по белковым наукам в Calibr, предназначена для повышения безопасности, универсальности и эффективности за счет запатентованной модульной «переключаемой» клетки CAR-T, которая использует молекулы-переключатели на основе антител для управления активации и антигенной специфичности клеток CAR-T.
Измеряя ширину всего 45 мм, samosPRO COMPACT обеспечивает до 20 безопасных входов и 8 безопасных выходов с использованием 4 переключаемых безопасных входов / выходов, система может быть расширена дополнительными модулями до 116 безопасных входов и 56 безопасных выходов и любого промежуточного числа.Функция переключения черного цвета доступна в качестве обновления как для новых, так и для существующих клиентов Pro8432WT. Новый Street Twin теперь имеет два режима езды — дождь и дорога, а также переключаемый контроль тяги с муфтой помощи крутящего момента. D25 оснащен независимыми поворотными инструментами с регулируемым углом к вспомогательному шпинделю, 3 оси Y и 3 оси Z, ось B для обработки спереди и сзади, до 59 инструментов, возможность переключения для использования с направляющей втулкой или без нее, система ЧПУ нового поколения с сенсорным экраном и различные интерфейсы выбор (USB-память, SD-карта, RS232, Ethernet).Новая программа клеточной терапии Calibr, возглавляемая Тревисом Янгом, доктором философии, директором по белковым наукам в Calibr, предназначена для повышения безопасности, универсальности и эффективности за счет запатентованной модульной «переключаемой» клетки CAR-T, которая использует молекулы-переключатели на основе антител для управления активации и антигенной специфичности клеток CAR-T. Для тех, кто озабочен защитой от сверхтоков, серия VENGEANCE включает в себя переключаемую конструкцию с одной / несколькими шинами +12 В, что позволяет пользователям мгновенно переключаться с нескольких шин +12 В с OCP на каждом кабеле на одну + 12В рейка; позволяет использовать все возможности блока питания +12 В на любом из разъемов постоянного тока блока питания.Базируясь в Тель-Авиве, Гаузи успешно реализовала на рынке несколько начальных продуктов «умного стекла», в том числе встроенную переключаемую оконную пленку, которая одним нажатием кнопки может переключаться с прозрачной на матовую. Истоки этой технологии начались с профессора химии Университета Королевы Филипа Джессопа. , который потратил более десяти лет на переработку ряда «переключаемых» материалов, которые становятся гидрофильными или гидрофобными за счет простого взаимодействия с диоксидом углерода. Модуль вертикального подъема OnePick включает в себя переключаемые режимы работы, которые позволяют использовать машину в качестве стандартного VLM с неограниченным доступ к полному лотку или с функцией ограничения инвентаря.
Для тех, кто озабочен защитой от сверхтоков, серия VENGEANCE включает в себя переключаемую конструкцию с одной / несколькими шинами +12 В, что позволяет пользователям мгновенно переключаться с нескольких шин +12 В с OCP на каждом кабеле на одну + 12В рейка; позволяет использовать все возможности блока питания +12 В на любом из разъемов постоянного тока блока питания.Базируясь в Тель-Авиве, Гаузи успешно реализовала на рынке несколько начальных продуктов «умного стекла», в том числе встроенную переключаемую оконную пленку, которая одним нажатием кнопки может переключаться с прозрачной на матовую. Истоки этой технологии начались с профессора химии Университета Королевы Филипа Джессопа. , который потратил более десяти лет на переработку ряда «переключаемых» материалов, которые становятся гидрофильными или гидрофобными за счет простого взаимодействия с диоксидом углерода. Модуль вертикального подъема OnePick включает в себя переключаемые режимы работы, которые позволяют использовать машину в качестве стандартного VLM с неограниченным доступ к полному лотку или с функцией ограничения инвентаря. Корал-Спрингс, Флорида, 16 июня 2016 г. — (PR.com) — VISTAMATIC, пионер в области интегрированных решений для затемнения и защитного стекла, рада объявить о запуске Clarity, своего нового решения с переключаемым защитным стеклом, которое обеспечивает конфиденциальность и конфиденциальность. наблюдение для медицинских, коммерческих и жилых приложений.
Корал-Спрингс, Флорида, 16 июня 2016 г. — (PR.com) — VISTAMATIC, пионер в области интегрированных решений для затемнения и защитного стекла, рада объявить о запуске Clarity, своего нового решения с переключаемым защитным стеклом, которое обеспечивает конфиденциальность и конфиденциальность. наблюдение для медицинских, коммерческих и жилых приложений.Переключаемая медленная сотовая проводимость определяет надежность и настраиваемость состояний сети
Abstract
Обработка нейронной информации регулируется быстрыми и локализованными колебаниями состояний мозга.Состояния мозга надежно переключаются между различными пространственно-временными сигнатурами в масштабе сети, даже если они состоят из гетерогенных и переменных ритмов в масштабе клетки. Мы исследовали механизмы этого сетевого управления в модели населения на основе проводимости, которая надежно переключается между активным и осцилляторным средними полями. Надежный контроль свойств среднего поля критически зависит от переключаемой отрицательной собственной проводимости на клеточном уровне. Эта проводимость наделяет цепи общей клеточной положительной обратной связью, которая может включать и выключать популяционные ритмы с клеточным разрешением.Переключатель в значительной степени не зависит от других внутренних свойств нейронов, размера сети и синаптических связей. Следовательно, он совместим с временной изменчивостью и пространственной неоднородностью, вызванной более медленными регуляторными функциями, такими как нейромодуляция, синаптическая пластичность и гомеостаз. Поразительно, что требуемый клеточный механизм доступен во всех типах клеток, которые обладают кальциевыми каналами Т-типа, но недоступен в вычислительных моделях, которые игнорируют медленную кинетику их активации.
Эта проводимость наделяет цепи общей клеточной положительной обратной связью, которая может включать и выключать популяционные ритмы с клеточным разрешением.Переключатель в значительной степени не зависит от других внутренних свойств нейронов, размера сети и синаптических связей. Следовательно, он совместим с временной изменчивостью и пространственной неоднородностью, вызванной более медленными регуляторными функциями, такими как нейромодуляция, синаптическая пластичность и гомеостаз. Поразительно, что требуемый клеточный механизм доступен во всех типах клеток, которые обладают кальциевыми каналами Т-типа, но недоступен в вычислительных моделях, которые игнорируют медленную кинетику их активации.
Информация об авторе
Обработка информации в головном мозге включает электрофизиологические сигналы в различных временных и пространственных масштабах времени, от уровня отдельного нейрона до целых областей мозга. Быстрый и локальный контроль этих сигналов с помощью нейрохимических веществ, называемых нейромодуляторами, необходим в сложных задачах, таких как инициация движения и фокусировка внимания. Нейромодуляторы действуют на клеточном уровне, чтобы управлять сигналами, которые потенциально могут распространяться в гораздо больших масштабах. В данной статье подчеркивается критическая роль клеточного переключателя возбудимости для быстрого и локализованного контроля состояний клетки и сети.Включая и выключая клеточный переключатель, нейромодуляторы могут надежно переключать большие группы населения между различными состояниями сети. Мы подчеркиваем важность управления коммутатором на сотовом уровне и независимо от возможности подключения, чтобы учесть настраиваемые пространственно-временные сигнатуры состояний сети.
Нейромодуляторы действуют на клеточном уровне, чтобы управлять сигналами, которые потенциально могут распространяться в гораздо больших масштабах. В данной статье подчеркивается критическая роль клеточного переключателя возбудимости для быстрого и локализованного контроля состояний клетки и сети.Включая и выключая клеточный переключатель, нейромодуляторы могут надежно переключать большие группы населения между различными состояниями сети. Мы подчеркиваем важность управления коммутатором на сотовом уровне и независимо от возможности подключения, чтобы учесть настраиваемые пространственно-временные сигнатуры состояний сети.
Образец цитирования: Drion G, Dethier J, Franci A, Sepulcher R (2018) Переключаемая медленная сотовая проводимость определяет надежность и настраиваемость состояний сети. PLoS Comput Biol 14 (4): e1006125.https://doi.org/10.1371/journal.pcbi.1006125
Редактор: Уильям В. Литтон, SUNY Downstate Medical Center, США
Поступила: 4 июля 2017 г . ; Принята к печати: 6 апреля 2018 г .; Опубликован: 23 апреля 2018 г.
; Принята к печати: 6 апреля 2018 г .; Опубликован: 23 апреля 2018 г.
Авторские права: © 2018 Drion et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: файлы кода Julia и matlab находятся в свободном доступе по адресу http://www.montefiore.ulg.ac.be/~guilldrion/Files/Drionetal2018-Code.zip и https://osf.io/k86en .
Финансирование: Исследование, приведшее к этим результатам, получило финансирование от Европейского исследовательского совета в рамках Соглашения о гранте Advanced ERC № 670645 и DGAPA-UNAM в рамках гранта PAPIIT RA105816. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Обработка нейронов постоянно формируется колебаниями ритмической активности населения, каждое из которых определяет особые состояния мозга [1–4]. Нейромодуляторы организуют переключение между различными состояниями мозга [5,6], изменяя способ обработки сетей нейронными сигналами [7,8]. Для изменений, связанных с движением, вниманием, восприятием, мотивацией или ожиданием, требуется точный временной и пространственный контроль состояний мозга [9–18].Быстро действующие пути нейротрансмиттеров обеспечивают быструю кинетику, необходимую для быстрых изменений состояний сети и обработки сигналов [17]. Сообщалось, что быстрый контроль состояний сети влияет на пространственное внимание в корковых цепях [6,7], внимание и возбуждение в таламусе [7,18] и на инициацию движения в субталамическом ядре [14]. Вероятно, наиболее изученным примером является таламокортикальная система. Таламус действует как пластичное реле между сенсорными системами, различными подкорковыми областями и корой головного мозга, управляя и модулируя поток нейронных сигналов под модулирующим эффектом корковой обратной связи [19–22].
Таламус действует как пластичное реле между сенсорными системами, различными подкорковыми областями и корой головного мозга, управляя и модулируя поток нейронных сигналов под модулирующим эффектом корковой обратной связи [19–22].
Экспериментально состояния мозга идентифицируются с помощью определенных пространственно-временных характеристик средней электрической активности больших популяций нейронов. Сдвиги ритмической активности происходят при переходе к медленноволновому сну и сонным веретенам [1,7,8,23–27]. Эти сдвиги коррелируют с сильными изменениями в обработке афферентных нейрональных сигналов [8,17,28]. Крайняя ситуация — это когда высокосинхронизированные колебания сна перерастают в абсанс-эпилепсию — поведенческое состояние, которое можно рассматривать как отключение мозга от внешнего мира [29–31].В состоянии бодрствования также наблюдаются переходные переключатели состояния сети, которые коррелируют с модуляциями обработки сенсомоторных сигналов [16].
Каковы механизмы, которые обеспечивают быстрое и надежное переключение среднего поля в гетерогенных популяциях нейронов, которые демонстрируют ритмы в широком диапазоне временных и пространственных масштабов, от отдельных клеток до сетей?
На клеточном уровне ритмы определяются определенным балансом определенных ионных токов. Конкретные синаптические соединения определяют топологию конкретных схем, которые определяют новые и разные ритмы в масштабе схемы.На сетевом уровне топологии схем соединяют большие и разнородные группы нейронов. В совокупности популяции формируют активность среднего поля, которая определяет еще один ритм состояния мозга. На каждом уровне ритмы постоянно меняются под действием нейромодуляторов, которые со временем модулируют клеточную и синаптическую проводимость. Нейромодуляторные системы действуют на клеточном уровне, но их широкие проекции могут одновременно влиять на большие популяции [6,7,17,27]. Глобальные нейромодуляторы контролируют переключение состояний мозга [16,32–34].Вопрос, который мы хотим исследовать в этой статье, заключается в том, как этот глобальный контроль может справиться с неоднородностью и изменчивостью ритмов в клеточном масштабе и получить от этого пользу.
Конкретные синаптические соединения определяют топологию конкретных схем, которые определяют новые и разные ритмы в масштабе схемы.На сетевом уровне топологии схем соединяют большие и разнородные группы нейронов. В совокупности популяции формируют активность среднего поля, которая определяет еще один ритм состояния мозга. На каждом уровне ритмы постоянно меняются под действием нейромодуляторов, которые со временем модулируют клеточную и синаптическую проводимость. Нейромодуляторные системы действуют на клеточном уровне, но их широкие проекции могут одновременно влиять на большие популяции [6,7,17,27]. Глобальные нейромодуляторы контролируют переключение состояний мозга [16,32–34].Вопрос, который мы хотим исследовать в этой статье, заключается в том, как этот глобальный контроль может справиться с неоднородностью и изменчивостью ритмов в клеточном масштабе и получить от этого пользу.
Аналогичный вопрос привлек значительное внимание в последние два десятилетия при изучении нейромодуляции малых ритмических цепей, контролирующих ритмы пилорического и желудочного мельниц краба [35–38]. Эта работа в значительной степени прояснила, как неоднородность и изменчивость в клеточном масштабе не только совместимы с однородностью и стабильностью в масштабе цепи, но фактически являются важным источником устойчивости и настраиваемости в ритмах цепи.Настоящая статья вдохновлена этим направлением работы: мы разработали простую вычислительную модель на основе проводимости, чтобы исследовать, как неоднородность и изменчивость в клеточном и схемном масштабах способствуют надежному контролю и настраиваемости состояний мозга.
Эта работа в значительной степени прояснила, как неоднородность и изменчивость в клеточном масштабе не только совместимы с однородностью и стабильностью в масштабе цепи, но фактически являются важным источником устойчивости и настраиваемости в ритмах цепи.Настоящая статья вдохновлена этим направлением работы: мы разработали простую вычислительную модель на основе проводимости, чтобы исследовать, как неоднородность и изменчивость в клеточном и схемном масштабах способствуют надежному контролю и настраиваемости состояний мозга.
Предыдущие вычислительные модели состояний мозга фокусировались на роли изменений связности в модуляции сетевого ритма [39–41]. Чтобы учесть быстрые колебания, наша модель вместо этого изучает сетевые коммутаторы, которые не требуют изменений в сетевых подключениях.Мы предполагаем, что переключение среднего поля является результатом переключения сотовой связи, которое используется значительной частью населения. Этот механизм в значительной степени не зависит от топологии сети, и всегда предполагается, что подключение к сети будет слабым .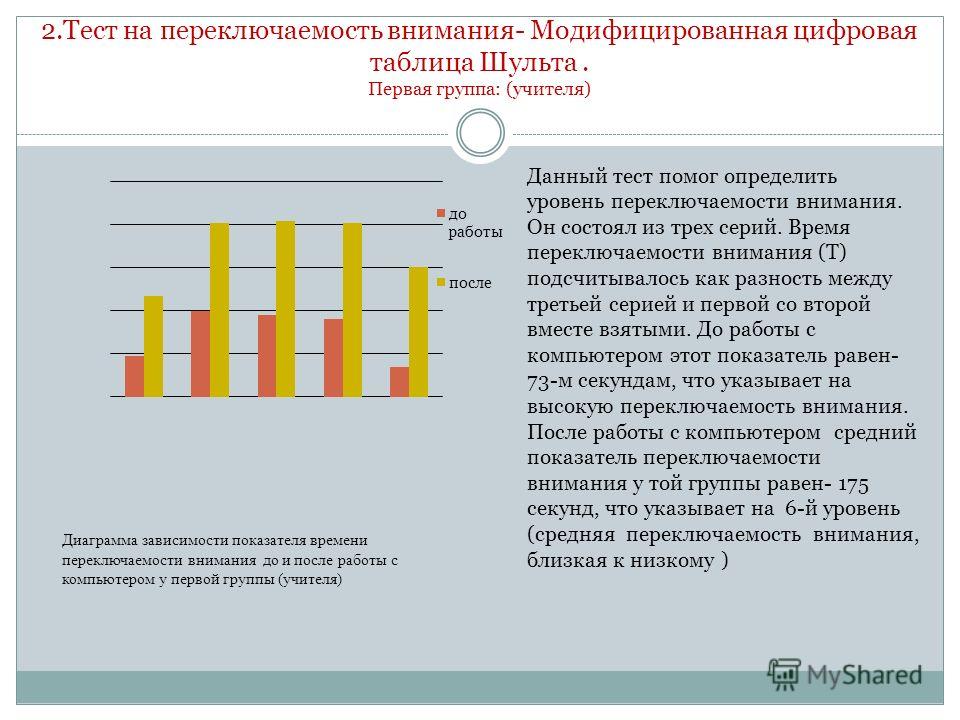 Это то, что допускает ритмическую неоднородность в популяции. Клеточные переключатели управляют только дискретным переходом между двумя различными режимами возбудимости, которые классически называются тоническим возбуждением и взрывом [42]. Слабая связь делает этот дискретный переход совместимым с непрерывной настройкой каждого дискретного состояния.Однородный контроль состояний мозга на сетевом уровне зависит только от общего сотового коммутатора. Он совместим с гетерогенными и вариабельными ритмами в клеточном и круговом масштабах.
Это то, что допускает ритмическую неоднородность в популяции. Клеточные переключатели управляют только дискретным переходом между двумя различными режимами возбудимости, которые классически называются тоническим возбуждением и взрывом [42]. Слабая связь делает этот дискретный переход совместимым с непрерывной настройкой каждого дискретного состояния.Однородный контроль состояний мозга на сетевом уровне зависит только от общего сотового коммутатора. Он совместим с гетерогенными и вариабельными ритмами в клеточном и круговом масштабах.
Наша статья направлена на демонстрацию вычислительной и физиологической значимости этого нового механизма как для сетевых исследований, так и для клеточной физиологии. Что касается сетевых вычислений, мы показываем, что неоднородность и изменчивость в масштабе сотовой связи и канала способствует устойчивости и настраиваемости в масштабе сети.Сотовый коммутатор отделяет управление состояниями сети, которое является быстрым и глобальным, от настройки пространственно-временного ритма, что обеспечивается модуляцией внутренних и синаптических свойств в более медленных временных и более мелких пространственных масштабах. Разделение задач между переключением , и настройкой позволяет точно регулировать как колебательное, так и активное состояния сети. Эта настройка состояний имеет существенное функциональное значение, например, модуляция свойств передачи сети в ее активном состоянии.
Разделение задач между переключением , и настройкой позволяет точно регулировать как колебательное, так и активное состояния сети. Эта настройка состояний имеет существенное функциональное значение, например, модуляция свойств передачи сети в ее активном состоянии.
На физиологическом уровне наши результаты подчеркивают роль на сетевом уровне конкретных ионных механизмов, которые долгое время изучались в нейрофизиологии одиночных клеток, но часто игнорируются в сетевых исследованиях ради числовой или математической управляемости. Сотовый переключатель нашей модели полностью основан на регулируемой медленной отрицательной проводимости. Переключающая роль этой медленной проводимости изучалась в ряде недавних работ авторов. Он регулирует клеточную модуляцию типов возбудимости [43–46] и, в частности, переход между тоническим возбуждением и взрывом [47, 48].Это критически важно для устойчивости и настраиваемости пакетной передачи данных [42]. И это очень важно для устойчивости ритмических контуров, таких как полуцентровые генераторы [49]. В продолжение этого исследования в настоящей статье показано, как медленная отрицательная клеточная проводимость способствует сетевой нейромодуляции, подчеркивая физиологическое значение конкретного клеточного механизма на сетевом уровне.
В продолжение этого исследования в настоящей статье показано, как медленная отрицательная клеточная проводимость способствует сетевой нейромодуляции, подчеркивая физиологическое значение конкретного клеточного механизма на сетевом уровне.
Результаты
Устойчивые и настраиваемые состояния сети, состоящие из разнородных состояний сотовой связи и цепей
Наша вычислительная модель воспроизводит общий пример переключения сети между активным и колебательным состояниями в популяции нейронов.На клеточном уровне мы использовали основанную на проводимости модель, которая включает типичные быстрые и медленные ионные токи разрывающихся ячеек. Каждая клетка может быть надежно модулирована гиперполяризованным входом между двумя различными режимами возбудимости: режим с быстрой деполяризацией, , склонный к всплескам и тоническому возбуждению, и режим , медленный гиперполяризованный, , склонный к взрыву.
На сетевом уровне мы включили соединения AMPA, GABA A и GABA B для моделирования асимметричной связи между субпопуляцией возбуждающих (E) клеток и субпопуляцией тормозных (I) клеток. Эта топология типична для областей мозга, участвующих в государственном регулировании, таких как, например, таламус [7], кора [2] и субталамическое ядро / бледный шар [39]. Наша модель не учитывает внутрипопуляционные связи, что максимизирует неоднородность клеточных ритмов в каждой субпопуляции. Напротив, он предполагает всеобщую связь между двумя популяциями, но со слабыми и случайно распределенными синаптическими весами (подробности см. В методах). Мы исследовали ритмические свойства сети, которые допускают широкую неоднородность как внутренних, так и синаптических параметров максимальной проводимости.Эта параметрическая неоднородность порождает широкий спектр ритмов на клеточном и круговом уровнях.
Эта топология типична для областей мозга, участвующих в государственном регулировании, таких как, например, таламус [7], кора [2] и субталамическое ядро / бледный шар [39]. Наша модель не учитывает внутрипопуляционные связи, что максимизирует неоднородность клеточных ритмов в каждой субпопуляции. Напротив, он предполагает всеобщую связь между двумя популяциями, но со слабыми и случайно распределенными синаптическими весами (подробности см. В методах). Мы исследовали ритмические свойства сети, которые допускают широкую неоднородность как внутренних, так и синаптических параметров максимальной проводимости.Эта параметрическая неоднородность порождает широкий спектр ритмов на клеточном и круговом уровнях.
Переключатели схемы и сети показаны на рис. 1. Они контролируются переходной гиперполяризацией тормозных нейронов, которые, например, имитируют активацию рецепторов GABA_B. На клеточном уровне гиперполяризация вызывает переключение из режима быстрой возбудимости в режим медленной возбудимости. Клеточный переключатель, в свою очередь, вызывает переключение на уровне цепи, потому что медленный режим возбудимости вызывает механизмы отскока между возбуждающей и тормозящей клеткой.Рис. 1A и Рис. 1B иллюстрируют возможности устойчивости и модуляции ритмов в изолированных цепях E-I двух клеток. На рис. 1А показано, что временная гиперполяризация может надежно вызвать переключение с асинхронного всплеска на синхронный всплеск, и что этот переключатель устойчив к изменениям внутренних и синаптических свойств нейронов, вызванных постоянной глобальной нейромодуляцией («NMD») и синаптической пластичностью (« Син. Пласт. »). Благодаря устойчивости клеточного переключателя к изменчивости, ритм цепи надежно поддерживается в широком диапазоне внутренних и синаптических параметров.В результате постоянный нейромодулятор, влияющий как на внутренние, так и на синаптические параметры, может модулировать клеточные и контурные ритмы между последовательными возникновениями временной гиперполяризации (рис.
Клеточный переключатель, в свою очередь, вызывает переключение на уровне цепи, потому что медленный режим возбудимости вызывает механизмы отскока между возбуждающей и тормозящей клеткой.Рис. 1A и Рис. 1B иллюстрируют возможности устойчивости и модуляции ритмов в изолированных цепях E-I двух клеток. На рис. 1А показано, что временная гиперполяризация может надежно вызвать переключение с асинхронного всплеска на синхронный всплеск, и что этот переключатель устойчив к изменениям внутренних и синаптических свойств нейронов, вызванных постоянной глобальной нейромодуляцией («NMD») и синаптической пластичностью (« Син. Пласт. »). Благодаря устойчивости клеточного переключателя к изменчивости, ритм цепи надежно поддерживается в широком диапазоне внутренних и синаптических параметров.В результате постоянный нейромодулятор, влияющий как на внутренние, так и на синаптические параметры, может модулировать клеточные и контурные ритмы между последовательными возникновениями временной гиперполяризации (рис. 1А и 1В).
1А и 1В).
Рис. 1. Надежный сетевой коммутатор, совместимый с глобальной нейромодуляцией, синаптической пластичностью и гомеостазом.
A. Вариации схемы возбуждения двух взаимосвязанных нейронов (один возбуждающий нейрон, синий, и один тормозной нейрон, красный) под контролем внешнего тока (I , приложение , оранжевый), медленные нейромодуляторы (NMD, черный) и синаптическая пластичность (Syn.Пласт., В черном цвете). Черная метка показывает периоды, в течение которых активны медленный нейромодулятор и синаптическая пластичность. Они моделируются как случайные изменения максимальной проводимости ионного канала и рецептора. Возбуждающий нейрон связан с тормозным нейроном через синапсы AMPA, а тормозящий нейрон связан с возбуждающим нейроном через синапсы GABA A и GABA B . Внешний ток, который временно гиперполяризует тормозящий нейрон, переключает ритм цепи.Быстрое переключение устойчиво к ритмической изменчивости, вызванной медленными нейромодуляторами и синаптической пластичностью (сравните ритм, генерируемый быстрым переключением до и после действия NMD и Syn. Plast.). В . Количественная оценка изменчивости ритмов, которые могут быть вызваны действием медленных нейромодуляторов и синаптической пластичности без нарушения быстрого переключения. На рисунке показана разница в периоде пакетной передачи (PER), количестве импульсов в пакетах (SPB), частоте внутри пакета (IBF) и коэффициенте заполнения пакета (DC) ритма, индуцированного быстрым переключением, до и после применения 100 различных тонов медленная нейромодуляция и син.Пласт. (журнал (параметр_after / параметр_before)) C . Спектрограмма потенциалов локального поля (LFP) возбуждающего нейрона (вверху) и популяций тормозных нейронов в сети из 200 клеток (100 возбуждающих клеток полностью связаны со 100 тормозными клетками со случайными синаптическими весами, взятыми в фиксированном диапазоне). Оранжевая кривая внизу показывает период, в течение которого внешний гиперполяризующий ток применяется к тормозящим нейронам. Показано, что гиперполяризация временно переключает ритм среднего поля населения, что показано появлением временной полосы высокой мощности на спектрограмме.
Plast.). В . Количественная оценка изменчивости ритмов, которые могут быть вызваны действием медленных нейромодуляторов и синаптической пластичности без нарушения быстрого переключения. На рисунке показана разница в периоде пакетной передачи (PER), количестве импульсов в пакетах (SPB), частоте внутри пакета (IBF) и коэффициенте заполнения пакета (DC) ритма, индуцированного быстрым переключением, до и после применения 100 различных тонов медленная нейромодуляция и син.Пласт. (журнал (параметр_after / параметр_before)) C . Спектрограмма потенциалов локального поля (LFP) возбуждающего нейрона (вверху) и популяций тормозных нейронов в сети из 200 клеток (100 возбуждающих клеток полностью связаны со 100 тормозными клетками со случайными синаптическими весами, взятыми в фиксированном диапазоне). Оранжевая кривая внизу показывает период, в течение которого внешний гиперполяризующий ток применяется к тормозящим нейронам. Показано, что гиперполяризация временно переключает ритм среднего поля населения, что показано появлением временной полосы высокой мощности на спектрограмме. D. Пример следов активности отдельного нейрона в возбуждающей (синий) и тормозной субпопуляциях (красный) для сетевого переключателя, показанного в C . Оранжевая кривая внизу показывает период, в течение которого внешний гиперполяризующий ток применяется к тормозящим нейронам.
D. Пример следов активности отдельного нейрона в возбуждающей (синий) и тормозной субпопуляциях (красный) для сетевого переключателя, показанного в C . Оранжевая кривая внизу показывает период, в течение которого внешний гиперполяризующий ток применяется к тормозящим нейронам.
https://doi.org/10.1371/journal.pcbi.1006125.g001
На сетевом уровне среднее поле совокупности определяет переключение сети между колебательным состоянием , соответствующим медленному режиму клеточной возбудимости, и активное состояние , соответствующее быстрому режиму клеточной возбудимости.Сетевой коммутатор устойчив к временной изменчивости и пространственной неоднородности населения, поскольку клеточный коммутатор существует в широком диапазоне внутренних и синаптических параметров. Эта надежность делает внешнее управление сетью в значительной степени независимым от размера сети и возможности подключения. Рис. 1C и 1D иллюстрируют это свойство в гетерогенной сети из 200 ячеек. Активность потенциала локального поля (LFP) показывает, что временная гиперполяризация может включать и выключать ритмическую активность среднего поля (определяемую отмеченной полосой частот LFP высокой мощности) всей контролируемой популяции.Рис. 1D также показывает, что этот популяционный контроль совместим с гетерогенными ритмами при разрешении клеток и контуров. Другими словами, надежный контроль состояния сети совместим с ритмической неоднородностью и изменчивостью при клеточном разрешении. Сетевой ритм среднего поля не является результатом синхронизации клеточных или контурных ритмов. Вместо этого ритмическое состояние сети возникает из-за общего сотового коммутатора, который устойчив к изменчивости мелодий при сотовом разрешении.
Активность потенциала локального поля (LFP) показывает, что временная гиперполяризация может включать и выключать ритмическую активность среднего поля (определяемую отмеченной полосой частот LFP высокой мощности) всей контролируемой популяции.Рис. 1D также показывает, что этот популяционный контроль совместим с гетерогенными ритмами при разрешении клеток и контуров. Другими словами, надежный контроль состояния сети совместим с ритмической неоднородностью и изменчивостью при клеточном разрешении. Сетевой ритм среднего поля не является результатом синхронизации клеточных или контурных ритмов. Вместо этого ритмическое состояние сети возникает из-за общего сотового коммутатора, который устойчив к изменчивости мелодий при сотовом разрешении.
Переключение состояния сети гетерогенного населения критически зависит от свойства сотовой связи
Наша вычислительная модель демонстрирует надежное переключение среднего поля на сетевом уровне, несмотря на гетерогенность и изменчивость на клеточном уровне. Такое свойство не предоставляется в вычислительной модели, которая зависит от тысяч неопределенных и переменных параметров. В нашей вычислительной модели он критически полагается на переключаемую медленную отрицательную проводимость на клеточном уровне.
Такое свойство не предоставляется в вычислительной модели, которая зависит от тысяч неопределенных и переменных параметров. В нашей вычислительной модели он критически полагается на переключаемую медленную отрицательную проводимость на клеточном уровне.
Медленная отрицательная проводимость нейрона определяет его медленную возбудимость так же, как быстрая отрицательная проводимость определяет его быструю возбудимость. Точно так же, как активация натриевого канала позволяет быстро переключиться с состояния покоя на спайк, медленная отрицательная проводимость дает возможность медленного перехода от состояния покоя к взрыву. Ионные каналы, которые могут способствовать медленной отрицательной проводимости нейрона, называются медленными регенеративными [45,46,48]. Канал является медленным регенерирующим, если он активирует входящий ток или деактивирует внешний ток во временной шкале, которая медленна по сравнению с быстрой шкалой времени активации натрия.В нашей модели только кальциевые каналы Т-типа восстанавливаются медленно. Их активация n является медленной относительно активации натриевых каналов [50]. Более того, из-за их инактивирующей, низкопороговой природы кальциевые каналы Т-типа снабжают нейрон медленной отрицательной проводимостью, которая переключается на внешним током: он включается при гиперполяризации и выключается при деполяризации. Этот механизм переключения отличается от классического механизма отскока (называемого постингибиторным отскоком, или PIR), связанного с кальциевыми каналами Т-типа и другими временными внутренними токами, такими как токи катионов гиперполяризации-активации [7,49,51,52].Он наделяет клетку медленной возбудимостью. Экспериментальное проявление этой медленной возбудимости, в частности, включает всплеск отскока (RB) (рис. 2A, слева) и взрыв, вызванный гиперполяризацией (HIB) (S1 рис.). Такое поведение широко наблюдалось экспериментально [7,53–58].
Их активация n является медленной относительно активации натриевых каналов [50]. Более того, из-за их инактивирующей, низкопороговой природы кальциевые каналы Т-типа снабжают нейрон медленной отрицательной проводимостью, которая переключается на внешним током: он включается при гиперполяризации и выключается при деполяризации. Этот механизм переключения отличается от классического механизма отскока (называемого постингибиторным отскоком, или PIR), связанного с кальциевыми каналами Т-типа и другими временными внутренними токами, такими как токи катионов гиперполяризации-активации [7,49,51,52].Он наделяет клетку медленной возбудимостью. Экспериментальное проявление этой медленной возбудимости, в частности, включает всплеск отскока (RB) (рис. 2A, слева) и взрыв, вызванный гиперполяризацией (HIB) (S1 рис.). Такое поведение широко наблюдалось экспериментально [7,53–58].
Рис. 2. Клеточный переключатель требует, чтобы кинетика активации кальция Т-типа была медленной; он теряется, когда активация моделируется как мгновенная.
А . Реакция модельного нейрона на применение временной гиперполяризации для двух разных наборов параметров в двух моделях, которые различаются только кинетикой активации кальциевых каналов Т-типа, которая является либо физиологически медленной (слева), либо мгновенной (справа).Для первого набора параметров (средние кривые) высвобождение гиперполяризации вызывает генерацию переходного периода всплеска в обеих моделях, свойство, называемое постингибиторным отскоком (PIR). Это наблюдение показывает, что PIR устойчив к кинетике активации кальциевых каналов Т-типа. Для второго набора параметров (нижние кривые) высвобождение гиперполяризации вызывает генерацию переходного периода всплеска в модели, имеющей медленную кинетику активации кальциевых каналов Т-типа (слева), свойство, называемое рикошетом (RB), тогда как это индуцирует PIR в модели с мгновенной кинетикой активации кальциевых каналов Т-типа (справа).Это наблюдение показывает, что, в отличие от PIR, RB чувствителен к кинетике активации кальциевых каналов Т-типа. В . Эксперименты с фиксацией напряжения в двух моделях одиночных нейронов, которые различаются только кинетикой активации кальциевых каналов Т-типа, которая является либо физиологически медленной (слева), либо мгновенной (справа), все остальные параметры идентичны. Верхние кривые показывают две ступени напряжения, приложенные к нейронам. Эти шаги различаются только начальным удерживающим потенциалом, который составляет либо -60 мВ (пунктирная серая кривая), либо -90 мВ (полностью черная кривая).Нижние кривые показывают ионные токи, зарегистрированные с течением времени во время приложения любого шага напряжения (пунктирные серые кривые — это ток, соответствующий удерживающему потенциалу -60 мВ, сплошные черные кривые — ток, соответствующий удерживающему потенциалу -90 мВ) . Ответы обеих моделей на шаг, начинающийся с -60 мВ, одинаковы (кальциевые каналы Т-типа инактивированы, а все другие параметры идентичны для двух моделей). Реакция обеих моделей на скачок, начинающийся с -90 мВ, сильно различается.
В . Эксперименты с фиксацией напряжения в двух моделях одиночных нейронов, которые различаются только кинетикой активации кальциевых каналов Т-типа, которая является либо физиологически медленной (слева), либо мгновенной (справа), все остальные параметры идентичны. Верхние кривые показывают две ступени напряжения, приложенные к нейронам. Эти шаги различаются только начальным удерживающим потенциалом, который составляет либо -60 мВ (пунктирная серая кривая), либо -90 мВ (полностью черная кривая).Нижние кривые показывают ионные токи, зарегистрированные с течением времени во время приложения любого шага напряжения (пунктирные серые кривые — это ток, соответствующий удерживающему потенциалу -60 мВ, сплошные черные кривые — ток, соответствующий удерживающему потенциалу -90 мВ) . Ответы обеих моделей на шаг, начинающийся с -60 мВ, одинаковы (кальциевые каналы Т-типа инактивированы, а все другие параметры идентичны для двух моделей). Реакция обеих моделей на скачок, начинающийся с -90 мВ, сильно различается. Модель с физиологически медленной кинетикой активации кальциевых каналов Т-типа показывает две фазы увеличения входящего тока: быструю (оранжевым) и медленную (зеленую). Модель с мгновенной кинетикой активации кальциевых каналов Т-типа показывает только одну усиленную быструю фазу увеличения входящего тока (оранжевым цветом). Однако обе текущие кривые достигают одного и того же текущего уровня в установившемся состоянии, показывая, что разница между двумя моделями носит динамический характер. C. Сравнение коммутационных возможностей в 2-клеточных цепях со случайной внутренней и синаптической проводимостью с использованием моделей нейронов с физиологически медленной кинетикой активации кальциевых каналов Т-типа (слева) или мгновенной кинетикой активации кальциевых каналов Т-типа (справа).Верхние кривые показывают примеры нейронной активности до и после приложения гиперполяризующего тока к тормозному нейрону (возбуждающий нейрон изображен синим цветом, а тормозящий нейрон изображен красным).
Модель с физиологически медленной кинетикой активации кальциевых каналов Т-типа показывает две фазы увеличения входящего тока: быструю (оранжевым) и медленную (зеленую). Модель с мгновенной кинетикой активации кальциевых каналов Т-типа показывает только одну усиленную быструю фазу увеличения входящего тока (оранжевым цветом). Однако обе текущие кривые достигают одного и того же текущего уровня в установившемся состоянии, показывая, что разница между двумя моделями носит динамический характер. C. Сравнение коммутационных возможностей в 2-клеточных цепях со случайной внутренней и синаптической проводимостью с использованием моделей нейронов с физиологически медленной кинетикой активации кальциевых каналов Т-типа (слева) или мгновенной кинетикой активации кальциевых каналов Т-типа (справа).Верхние кривые показывают примеры нейронной активности до и после приложения гиперполяризующего тока к тормозному нейрону (возбуждающий нейрон изображен синим цветом, а тормозящий нейрон изображен красным). Нижние гистограммы количественно определяют активность 1000 смоделированных случайных цепей при приложении 5 различных приложенных токов. Клетки либо молчаливы (белые), либо медленно всплывают (черный), либо разрываются асинхронно (серый цвет), либо участвуют в синхронном ритме взрывов (оранжевый).При медленной активации кальциевых каналов Т-типа большинство из 1000 смоделированных случайных цепей переключаются с быстрых ритмов на медленные при гиперполяризации (слева). Ни один из них не переключался при мгновенной активации (справа).
Нижние гистограммы количественно определяют активность 1000 смоделированных случайных цепей при приложении 5 различных приложенных токов. Клетки либо молчаливы (белые), либо медленно всплывают (черный), либо разрываются асинхронно (серый цвет), либо участвуют в синхронном ритме взрывов (оранжевый).При медленной активации кальциевых каналов Т-типа большинство из 1000 смоделированных случайных цепей переключаются с быстрых ритмов на медленные при гиперполяризации (слева). Ни один из них не переключался при мгновенной активации (справа).
https://doi.org/10.1371/journal.pcbi.1006125.g002
В [42] мы подробно изучили, почему медленная отрицательная проводимость критична для клеточной активности, которая позволяет надежно и контролируемо переключаться между быстрым и медленные настраиваемые ритмы и как этот механизм соотносится с моделями разрыва в литературе, такими как всплеск прямоугольной волны или параболический всплеск.Аналогичное наблюдение применимо к двухклеточной возбуждающе-ингибирующей цепи на рис. 1А. Цепной ритм является результатом хорошо известного механизма отскока [7,51], но это происходит только при включении медленной отрицательной проводимости. Чтобы оценить конкретную роль медленной отрицательной проводимости, мы действовали по тому же протоколу, что и в [42]: мы сравнили номинальную модель, в которой кинетика активации кальциевых каналов Т-типа физиологически является медленной , (около десяти время медленнее, чем кинетика активации натриевых каналов) к возмущенной модели, в которой кинетика активации кальциевых каналов Т-типа составляет мгновенных , то есть недифференцированная от кинетики активации натриевых каналов.Мгновенная активация кальциевых каналов — частое предположение при моделировании (см. Обсуждение). И в номинальной, и в возмущенной моделях кальциевые каналы Т-типа обеспечивают инактивирующий внутренний ток, необходимый для механизма отскока (рис. 2А). Однако режим медленной возбудимости теряется в модели возмущений, потому что и натриевые, и кальциевые каналы вносят вклад только в быструю отрицательную проводимость.
1А. Цепной ритм является результатом хорошо известного механизма отскока [7,51], но это происходит только при включении медленной отрицательной проводимости. Чтобы оценить конкретную роль медленной отрицательной проводимости, мы действовали по тому же протоколу, что и в [42]: мы сравнили номинальную модель, в которой кинетика активации кальциевых каналов Т-типа физиологически является медленной , (около десяти время медленнее, чем кинетика активации натриевых каналов) к возмущенной модели, в которой кинетика активации кальциевых каналов Т-типа составляет мгновенных , то есть недифференцированная от кинетики активации натриевых каналов.Мгновенная активация кальциевых каналов — частое предположение при моделировании (см. Обсуждение). И в номинальной, и в возмущенной моделях кальциевые каналы Т-типа обеспечивают инактивирующий внутренний ток, необходимый для механизма отскока (рис. 2А). Однако режим медленной возбудимости теряется в модели возмущений, потому что и натриевые, и кальциевые каналы вносят вклад только в быструю отрицательную проводимость. Это изменение четко прослеживается в эксперименте с ограничением напряжения (рис. 2В). В номинальной модели скачок напряжения от гиперполяризованного потенциала (-90 мВ) деактивирует кальциевые каналы Т-типа, что приводит к двум разным во времени фазам отрицательной проводимости: быстрой (изображена желтым на рис. 2В, слева) и медленной. (показано зеленым на рис. 2В, слева).Специфической сигнатуры медленной отрицательной проводимости в возмущенной модели нет. Вместо этого две отрицательные проводимости складываются в быстрой шкале времени. Выбранное возмущение имеет то преимущество, что оно не влияет на свойства модели в состоянии равновесия: номинальная и возмущенная модели имеют одинаковую кривую ВАХ и одинаковый баланс токов в установившемся состоянии. Рис. 2C иллюстрирует, что медленная кинетика активации кальциевых каналов важна для того, чтобы переключение двухклеточной цепи E-I было устойчивым к изменчивости параметров.Была смоделирована тысяча сетей из 2 ячеек путем случайной генерации 1000 различных наборов параметров для максимальной внутренней и синаптической проводимости (см.
Это изменение четко прослеживается в эксперименте с ограничением напряжения (рис. 2В). В номинальной модели скачок напряжения от гиперполяризованного потенциала (-90 мВ) деактивирует кальциевые каналы Т-типа, что приводит к двум разным во времени фазам отрицательной проводимости: быстрой (изображена желтым на рис. 2В, слева) и медленной. (показано зеленым на рис. 2В, слева).Специфической сигнатуры медленной отрицательной проводимости в возмущенной модели нет. Вместо этого две отрицательные проводимости складываются в быстрой шкале времени. Выбранное возмущение имеет то преимущество, что оно не влияет на свойства модели в состоянии равновесия: номинальная и возмущенная модели имеют одинаковую кривую ВАХ и одинаковый баланс токов в установившемся состоянии. Рис. 2C иллюстрирует, что медленная кинетика активации кальциевых каналов важна для того, чтобы переключение двухклеточной цепи E-I было устойчивым к изменчивости параметров.Была смоделирована тысяча сетей из 2 ячеек путем случайной генерации 1000 различных наборов параметров для максимальной внутренней и синаптической проводимости (см. Методы). На рис. 2С слева показано, что переключение цепи происходит в подавляющем большинстве конфигураций параметров, когда активация кальциевых каналов Т-типа является физиологической (> 90% для приложения I = -2,6 мкА / см 2 ). Это надежность, которая учитывает изменчивость, модуляцию (рис. 1A и 1B) и неоднородность (рис. 1C и 1D) при разрешении ячейки.
Методы). На рис. 2С слева показано, что переключение цепи происходит в подавляющем большинстве конфигураций параметров, когда активация кальциевых каналов Т-типа является физиологической (> 90% для приложения I = -2,6 мкА / см 2 ). Это надежность, которая учитывает изменчивость, модуляцию (рис. 1A и 1B) и неоднородность (рис. 1C и 1D) при разрешении ячейки.
Напротив, устойчивость и свойства настройки клеточных и контурных ритмов полностью теряются, когда активация кальция происходит быстро (рис. 2С, справа). Ритм контура E-I чрезвычайно уязвим для потери медленных регенеративных ионных каналов. Это требует очень точной настройки как ионных (внутренних), так и синаптических (внешних) параметров. Из 1000 случайных наборов параметров, смоделированных в этой конфигурации, не удалось обнаружить ритмическую активность схемы. При отсутствии медленной регенерации переключатель схемы не может быть отделен от ритма схемы: оба существуют только для очень специфических наборов параметров.
Неоднородность при клеточном разрешении способствует устойчивости и настраиваемости состояний сети в больших популяциях
Рис. 3A и 3B иллюстрируют непрерывную настройку ритмов на уровне контура в двух режимах дискретной возбудимости. На рис. 3А показано, что частоту тонического возбуждения и внутрипороговую частоту пакетного режима можно модулировать в широком диапазоне (в несколько раз). Все клетки подтверждают физиологическое свойство, заключающееся в том, что частота внутривзрывного импульса значительно выше, чем частота тонического возбуждения, что является ключевым признаком взрывной передачи сигналов и физиологической сигнатуры взрывов, вызванных гиперполяризацией [7,53–55].На рис. 3В показано, что два свойства медленного ритма (частота всплесков, верх и рабочий цикл всплеска, нижняя часть) также можно модулировать в широком диапазоне (более чем в 5 раз), не влияя на переключение, вызванное гиперполяризацией. Рис. 3C иллюстрирует свойство среднего поля нашей сети: несмотря на неоднородность ритмов в масштабах клетки и цепи, вариабельность ритмов в популяции постепенно уменьшается по мере увеличения размера сети и в конечном итоге исчезает для очень больших популяций нейронов ( предел среднего поля). Существование двух дискретных средних полей является следствием переключения сотовой связи. Неоднородность и изменчивость популяции вносят вклад в настраиваемость двух дискретных средних полей.
Существование двух дискретных средних полей является следствием переключения сотовой связи. Неоднородность и изменчивость популяции вносят вклад в настраиваемость двух дискретных средних полей.
Рис. 3. Устойчивость клеточного переключателя к внутренней и синаптической изменчивости позволяет выполнять большую настройку ритмических свойств.
A : Сравнение частоты тонических всплесков до внешней гиперполяризации и частоты внутри всплесков во время внешней гиперполяризации в 862 случайных двухэлементных цепях (из 1000 случайных двухэлементных смоделированных схем), которые показали переключение, вызванное гиперполяризацией, от тонического всплеска к синхронизированному взрыву (значения показаны для тормозных нейронов, возбуждающие нейроны в основном молчат в период деполяризации).Оба значения даны для каждой тормозной ячейки (черные точки), и линия связывает значения для тех же ячеек. Горизонтальные красные полоски определяют количество ячеек в каждом частотном диапазоне. Частота как тонического возбуждения, так и частоты внутри всплеска демонстрирует большую изменчивость в популяции, но частота внутри всплеска постоянно выше, чем частота тонического возбуждения в данном нейроне. B: Сравнение частоты всплеска (вверху) и рабочего цикла всплеска (внизу) в 914 случайных 2-элементных схемах (из 1000 случайных 2-элементных смоделированных цепей), которые показали синхронизированный ритм всплеска во время гиперполяризации.Вертикальные полосы определяют количество возбуждающих ячеек (синим цветом) и тормозных ячеек (красный цвет) в каждом диапазоне частот всплеска (вверху) или в каждом диапазоне рабочего цикла (внизу). Частота всплесков и рабочий цикл всплесков показывают большую изменчивость в популяции, показывая, что один и тот же переключатель цепи может создавать очень разные ритмы цепи. C : Распределение максимальной частоты мощности LFP во время гиперполяризации в 100 случайных, сильно неоднородных сетях из 10 ячеек (вверху), сети из 40 ячеек (в центре) и сетях из 100 ячеек (внизу).
Частота как тонического возбуждения, так и частоты внутри всплеска демонстрирует большую изменчивость в популяции, но частота внутри всплеска постоянно выше, чем частота тонического возбуждения в данном нейроне. B: Сравнение частоты всплеска (вверху) и рабочего цикла всплеска (внизу) в 914 случайных 2-элементных схемах (из 1000 случайных 2-элементных смоделированных цепей), которые показали синхронизированный ритм всплеска во время гиперполяризации.Вертикальные полосы определяют количество возбуждающих ячеек (синим цветом) и тормозных ячеек (красный цвет) в каждом диапазоне частот всплеска (вверху) или в каждом диапазоне рабочего цикла (внизу). Частота всплесков и рабочий цикл всплесков показывают большую изменчивость в популяции, показывая, что один и тот же переключатель цепи может создавать очень разные ритмы цепи. C : Распределение максимальной частоты мощности LFP во время гиперполяризации в 100 случайных, сильно неоднородных сетях из 10 ячеек (вверху), сети из 40 ячеек (в центре) и сетях из 100 ячеек (внизу). Красные вертикальные полосы показывают наличие сетей в каждом частотном диапазоне. Средняя спектральная мощность поля демонстрирует большую изменчивость в пределах совокупности сетей для небольших сетей, но разброс средней спектральной мощности поля постепенно сокращается по мере увеличения размера совокупности.
Красные вертикальные полосы показывают наличие сетей в каждом частотном диапазоне. Средняя спектральная мощность поля демонстрирует большую изменчивость в пределах совокупности сетей для небольших сетей, но разброс средней спектральной мощности поля постепенно сокращается по мере увеличения размера совокупности.
https://doi.org/10.1371/journal.pcbi.1006125.g003
Пространственно-временной локализованный контроль состояния сети
Из-за своей сотовой природы сетевой коммутатор, описанный в этой статье, позволяет точно контролировать пространственно-временное управление популяцией: его временное разрешение ограничено только кинетикой медленных регенеративных каналов; его пространственное разрешение ограничено только пространственным разрешением рецепторов на нейромодуляторные входы.Такой механизм обеспечивает пространственно-временное управление состоянием сети с множеством разрешений.
Рис. 4 иллюстрирует две простые формы пространственно-временного контроля в нашей модели. На рис. 4А показано, как на состояние сети популяции из 160 клеток влияет количество активных гиперполяризующих нейромодуляторных путей. Для иллюстрации нейромодуляторный вход поровну разделен на 8 путей. Глобальный сетевой ритм индуцируется гиперполяризацией достаточной субпопуляции I-клеток (4 пути из 8 на выбранной иллюстрации).И ритмическая спектральная мощность, и частотное распределение модулируются числом активных проводящих путей. Этот пример показывает, что временные свойства сетевого ритма могут модулироваться пространственно изменяющимися нейромодуляторными входами, даже при отсутствии изменений внутренних или синаптических параметров сети.
На рис. 4А показано, как на состояние сети популяции из 160 клеток влияет количество активных гиперполяризующих нейромодуляторных путей. Для иллюстрации нейромодуляторный вход поровну разделен на 8 путей. Глобальный сетевой ритм индуцируется гиперполяризацией достаточной субпопуляции I-клеток (4 пути из 8 на выбранной иллюстрации).И ритмическая спектральная мощность, и частотное распределение модулируются числом активных проводящих путей. Этот пример показывает, что временные свойства сетевого ритма могут модулироваться пространственно изменяющимися нейромодуляторными входами, даже при отсутствии изменений внутренних или синаптических параметров сети.
Рис. 4. Сотовая природа коммутатора позволяет осуществлять локальный пространственно-временной контроль состояния сети.
А, левый . Набросок пространственного сетевого взаимодействия и кластеризации в модулирующих путях.Возбуждающие клетки обозначены (полными и пустыми) зелеными точками, тормозные клетки (полными и пустыми) красными точками, возбуждающие синапсы — зелеными стрелками, а тормозящие синапсы — красными линиями с перпендикулярными полосами на их концах (возбуждающие и тормозные клетки соединены в всем всем мод). Горизонтальные двойные черные стрелки представляют пространственное измерение, а зеленый, красный и фиолетовый фон — пространственное распространение синапсов AMPA, GABA A и GABA B , исходящих от нейронов, представленных сплошными точками, соответственно (синапсы распределены равномерно в этом случае).Синие стрелки представляют различные модуляторные пути (представлены 3 пути: MOD 1, 2 и 3). Каждый модуляторный путь влияет только на субпопуляцию тормозных клеток, представленную сгруппированными наборами красных точек. А, правый . Спектрограмма потенциалов локального поля (LFP) возбуждающих нейронов (вверху) и тормозных нейронов (внизу) в 160-клеточной сети (80 возбуждающих клеток полностью соединены с 80 тормозными клетками со случайными синаптическими весами, взятыми в фиксированном диапазоне), разделенных в 8 кластеров, каждый из которых модулируется определенным нейромодуляторным путем.Галочка, синие следы внизу обозначают период, в течение которого гиперполяризация активна для каждого нейромодуляторного пути (от 1 до 8).
Горизонтальные двойные черные стрелки представляют пространственное измерение, а зеленый, красный и фиолетовый фон — пространственное распространение синапсов AMPA, GABA A и GABA B , исходящих от нейронов, представленных сплошными точками, соответственно (синапсы распределены равномерно в этом случае).Синие стрелки представляют различные модуляторные пути (представлены 3 пути: MOD 1, 2 и 3). Каждый модуляторный путь влияет только на субпопуляцию тормозных клеток, представленную сгруппированными наборами красных точек. А, правый . Спектрограмма потенциалов локального поля (LFP) возбуждающих нейронов (вверху) и тормозных нейронов (внизу) в 160-клеточной сети (80 возбуждающих клеток полностью соединены с 80 тормозными клетками со случайными синаптическими весами, взятыми в фиксированном диапазоне), разделенных в 8 кластеров, каждый из которых модулируется определенным нейромодуляторным путем.Галочка, синие следы внизу обозначают период, в течение которого гиперполяризация активна для каждого нейромодуляторного пути (от 1 до 8). Пространственная локализация модуляционных входов влияет на временную настройку средней частоты и амплитуды спектральной мощности LFP. B, слева: То же, что и A, слева , за исключением того факта, что здесь синапсы GABA B сгруппированы за счет добавления пространственного гауссова распада в синаптической силе (синапс GABA B сильный для соседних нейронов, но слабый для удаленных нейронов).AMPA и GABA A синапсы по-прежнему распределены равномерно. В, правый . Спектрограмма потенциалов локального поля (LFP) 4 кластеров возбуждающих нейронов (вверху) и популяций тормозных нейронов в 40-клеточной сети (20 возбуждающих клеток полностью соединены с 20 тормозными клетками со случайными синаптическими весами, взятыми в фиксированном диапазоне и пространственном кластеризация в синапсах GABA B ). Обе популяции разделены на 4 кластера, каждый из которых модулируется определенным нейромодуляторным путем.
Пространственная локализация модуляционных входов влияет на временную настройку средней частоты и амплитуды спектральной мощности LFP. B, слева: То же, что и A, слева , за исключением того факта, что здесь синапсы GABA B сгруппированы за счет добавления пространственного гауссова распада в синаптической силе (синапс GABA B сильный для соседних нейронов, но слабый для удаленных нейронов).AMPA и GABA A синапсы по-прежнему распределены равномерно. В, правый . Спектрограмма потенциалов локального поля (LFP) 4 кластеров возбуждающих нейронов (вверху) и популяций тормозных нейронов в 40-клеточной сети (20 возбуждающих клеток полностью соединены с 20 тормозными клетками со случайными синаптическими весами, взятыми в фиксированном диапазоне и пространственном кластеризация в синапсах GABA B ). Обе популяции разделены на 4 кластера, каждый из которых модулируется определенным нейромодуляторным путем. Голубые отметки внизу показывают период, в течение которого гиперполяризация активна для каждого нейромодуляторного пути (от 1 до 4). Пространственная локализация модулирующих входов и возможность подключения GABA B влияет на пространственную настройку состояния сети. Пространственная локализация соединений GABA B позволяет только пространственный контроль состояния сети, хотя соединения AMPA и GABA A являются универсальными.
Голубые отметки внизу показывают период, в течение которого гиперполяризация активна для каждого нейромодуляторного пути (от 1 до 4). Пространственная локализация модулирующих входов и возможность подключения GABA B влияет на пространственную настройку состояния сети. Пространственная локализация соединений GABA B позволяет только пространственный контроль состояния сети, хотя соединения AMPA и GABA A являются универсальными.
https://doi.org/10.1371 / journal.pcbi.1006125.g004
Рис. 4B иллюстрирует пространственную модуляцию состояния сети в полностью подключенном населении. В предложенной возбуждающе-тормозной топологии пространственной кластеризации в GABA B R достаточно только для создания кластеров в сетевом переключателе возбуждающей популяции (рис. 4B, слева). Когда один из нейромодуляторных путей активируется, он влияет только на Е-клетки, которые имеют достаточно сильные связи GABA B с модулированной ингибирующей субпопуляцией. В результате LFP возбуждающих субпопуляций настраиваются индивидуально, а временной ритм модулируется вдоль пространственной оси: каждый пространственно локализованный нейромодуляторный путь может включать и выключать соответствующий ритм (рис. 4B, справа). Каждый ритм имеет определенную характеристику спектральной мощности, а пространственная организация состояния сети контролируется с помощью пространственного разрешения нейромодуляции.
В результате LFP возбуждающих субпопуляций настраиваются индивидуально, а временной ритм модулируется вдоль пространственной оси: каждый пространственно локализованный нейромодуляторный путь может включать и выключать соответствующий ритм (рис. 4B, справа). Каждый ритм имеет определенную характеристику спектральной мощности, а пространственная организация состояния сети контролируется с помощью пространственного разрешения нейромодуляции.
В отличие от возбуждающей популяции, ритмическая активность тормозящей популяции на нашей иллюстрации пространственно однородна.Это связано с тем, что соединения AMPA от возбуждающей к тормозящей популяции не сгруппированы. В результате пространственно-временная организация состояний сети различна в каждой субпопуляции.
Пространственно-временное управление сетью полностью основано на отделении управления переключением от управления настройкой. Переключение всегда происходит в сотовом разрешении. Настройка происходит в любом временном масштабе медленнее, чем клеточный ритм, и в любом пространственном масштабе между клеточным и сетевым уровнем. Поскольку он управляет переключателем, медленная отрицательная проводимость на клеточном уровне снова имеет решающее значение для пространственно-временной управляемости сети.
Поскольку он управляет переключателем, медленная отрицательная проводимость на клеточном уровне снова имеет решающее значение для пространственно-временной управляемости сети.
Управление свойствами передачи сети путем модуляции активного состояния
Наш анализ до сих пор был сосредоточен на настройке медленных ритмов в колебательном состоянии сети. На рис. 5 показано, как модель также учитывает надежную настройку активного состояния, которое имеет решающее значение для модуляции свойств передачи населения.
Рис. 5. Сетевой коммутатор устойчив к модуляции свойств передачи в активном состоянии сети.
A : реакция возбуждающих клеток на импульс возбуждающего тока и приложенный синусоидальный ток для высокого отношения GABAR A / GABAR B (вверху) и низкого отношения GABAR A / GABAR B (внизу) в сети из 40 ячеек. Для каждого соотношения GABAR A / GABAR B верхняя черная кривая показывает ток, приложенный к возбуждающим нейронам с течением времени, синяя кривая показывает активность одного целевого возбуждающего нейрона с течением времени, красная кривая показывает активность тормозящий нейрон с течением времени, а нижняя черная кривая показывает ток, приложенный ко всем тормозным нейронам популяции с течением времени. B : Средняя реакция возбуждающих клеток (в импульсах в секунду) на приложенный синусоидальный ток для высокого (вверху), промежуточного (в центре) и низкого отношения GABAR A / GABAR B (внизу). Синие вертикальные полосы показывают среднюю частоту всплесков за каждый период времени. Изменение соотношения GABAR A / GABAR B сильно влияет на ретрансляционные свойства возбуждающих ячеек в активном состоянии без нарушения сетевого коммутатора, а взрывы в ретрансляционных ячейках сами по себе не запускают сетевой коммутатор, что делает эту схему активации совместимой. с активным состоянием сети.
B : Средняя реакция возбуждающих клеток (в импульсах в секунду) на приложенный синусоидальный ток для высокого (вверху), промежуточного (в центре) и низкого отношения GABAR A / GABAR B (внизу). Синие вертикальные полосы показывают среднюю частоту всплесков за каждый период времени. Изменение соотношения GABAR A / GABAR B сильно влияет на ретрансляционные свойства возбуждающих ячеек в активном состоянии без нарушения сетевого коммутатора, а взрывы в ретрансляционных ячейках сами по себе не запускают сетевой коммутатор, что делает эту схему активации совместимой. с активным состоянием сети.
https://doi.org/10.1371/journal.pcbi.1006125.g005
На рисунке показано, как электронные ячейки обрабатывают импульсы и синусоидальные входные сигналы в двух различных конфигурациях. Единственное различие между двумя конфигурациями — это соотношение силы синаптического соединения GABA A и GABA B . Это согласуется с физиологической регуляцией через синаптическую пластичность [56,57]. Управление переключателем в сети нечувствительно к синаптическому соотношению, но на свойства передачи E-клеток это заметно влияет (рис. 5A).Высокое соотношение усиливает линейный ответ E-клеток: короткий импульс возбуждающего тока вызывает всплеск, а синусоидальный вход увлекает за собой последовательность всплесков с синхронизацией по фазе (рис. 5A, вверху). Напротив, низкое отношение усиливает отклик детектора: короткий импульс возбуждающего тока вызывает всплеск всплесков, а синусоидальный вход вызывает всплеск всплесков, частота которых максимальна в начале фазы нарастания входного сигнала (рис. 5А, внизу). Эта модуляция свойств передачи дополнительно количественно выражена на фиг. 5В, которая иллюстрирует средний ответ нейрональной популяции на синусоидальный вход для трех различных синаптических соотношений.При высоком соотношении пиковый отклик достигается при максимальной амплитуде входного сигнала.
Это согласуется с физиологической регуляцией через синаптическую пластичность [56,57]. Управление переключателем в сети нечувствительно к синаптическому соотношению, но на свойства передачи E-клеток это заметно влияет (рис. 5A).Высокое соотношение усиливает линейный ответ E-клеток: короткий импульс возбуждающего тока вызывает всплеск, а синусоидальный вход увлекает за собой последовательность всплесков с синхронизацией по фазе (рис. 5A, вверху). Напротив, низкое отношение усиливает отклик детектора: короткий импульс возбуждающего тока вызывает всплеск всплесков, а синусоидальный вход вызывает всплеск всплесков, частота которых максимальна в начале фазы нарастания входного сигнала (рис. 5А, внизу). Эта модуляция свойств передачи дополнительно количественно выражена на фиг. 5В, которая иллюстрирует средний ответ нейрональной популяции на синусоидальный вход для трех различных синаптических соотношений.При высоком соотношении пиковый отклик достигается при максимальной амплитуде входного сигнала. Для низкого отношения пиковый отклик достигается, когда входной сигнал пересекает пороговое значение снизу. Такая модуляция режима передачи напоминает физиологические наблюдения в таламокортикальных петлях [7,8].
Для низкого отношения пиковый отклик достигается, когда входной сигнал пересекает пороговое значение снизу. Такая модуляция режима передачи напоминает физиологические наблюдения в таламокортикальных петлях [7,8].
Мы подчеркиваем, что модуляция активного состояния, показанная на рис. 5, снова критически зависит от медленной отрицательной проводимости на клеточном уровне. Соотношение между ГАМК A R и ГАМК B R в первую очередь регулирует поляризацию мембраны.Когда доминируют соединения GABA A R, ингибирующий импульс от I-клеток поддерживает E-клетки, близкие к потенциалу реверсии GABA A R, то есть потенциалу реверсии хлорида (установлен на -70 мВ в нашей модели). При этом потенциале кальциевые каналы Т-типа инактивируются, а медленная отрицательная проводимость отключается: Е-клетки проявляют физиологические признаки режима быстрой возбудимости: спайковую возбудимость и тоническое возбуждение [45,47,48]. Напротив, когда доминируют соединения GABA B R, ингибирующее стремление поддерживает E-клетки, близкие к потенциалу реверсии GABA B R, т. е.е. потенциал обратного калия (установлен на -85 мВ в этой модели). При этом потенциале кальциевые каналы Т-типа деинактивируются, и включается медленная отрицательная проводимость: Е-клетки проявляют физиологические признаки медленной возбудимости: взрывная возбудимость и эндогенный взрыв [45,47,48].
е.е. потенциал обратного калия (установлен на -85 мВ в этой модели). При этом потенциале кальциевые каналы Т-типа деинактивируются, и включается медленная отрицательная проводимость: Е-клетки проявляют физиологические признаки медленной возбудимости: взрывная возбудимость и эндогенный взрыв [45,47,48].
Обсуждение
Разделение управления переключением и настройкой
Пытаясь смоделировать пространственно-временную организацию состояний мозга, мы предложили простую архитектуру нейронной сети, которая использует модуляцию и гетерогенность ритмов при клеточном разрешении для настройки пространственно-временной сигнатуры больших ритмических популяций.В основе нашей модели лежит разделение механизмов переключения и механизмов настройки. Механизм переключателя прост. Он включает в себя единственное свойство клетки, происходит в едином временном масштабе и равномерно распределяется в популяции. Напротив, настраивающих механизмов несколько. Они включают как клеточные, так и синаптические свойства, встречаются во многих временных и пространственных масштабах и могут быть весьма неоднородными в популяции. Переход между двумя дискретными состояниями существует на всех уровнях, от соты к сетям, из-за единообразного сотового коммутатора.Напротив, пространственно-временные сигнатуры каждого дискретного состояния можно непрерывно настраивать для всей совокупности, формируя устойчивые и настраиваемые состояния сети. Главный вклад нашей модели — показать, что сотовый коммутатор важен для формирования свойств сети. Это необходимо для устойчивости сетевого коммутатора и обеспечивает возможность настройки состояний сети с помощью оставшейся внутренней и внешней проводимости.
Переход между двумя дискретными состояниями существует на всех уровнях, от соты к сетям, из-за единообразного сотового коммутатора.Напротив, пространственно-временные сигнатуры каждого дискретного состояния можно непрерывно настраивать для всей совокупности, формируя устойчивые и настраиваемые состояния сети. Главный вклад нашей модели — показать, что сотовый коммутатор важен для формирования свойств сети. Это необходимо для устойчивости сетевого коммутатора и обеспечивает возможность настройки состояний сети с помощью оставшейся внутренней и внешней проводимости.
Особая роль кальциевых каналов Т-типа
Управление сетью, описанное в этой статье, основывается только на двух специфических особенностях: свойстве клетки контролировать присущую медленной отрицательной проводимости, обеспечиваемой кальциевыми каналами Т-типа в нашей модели, и топологии сети, которая взаимно связывает возбуждающую субпопуляцию и тормозящую субпопуляция.Эти два свойства широко распространены среди множества схем, которые демонстрируют быстрый контроль сетевых ритмов [7,8,13–19,30,53–55,58–67]. Каноническим примером является таламус, где как роль ритмов отскока между (возбуждающими) таламокортикальными нейронами и (тормозящими) ретикулярными нейронами, так и важность кальциевых каналов Т-типа давно признаны в управлении колебательными состояниями сети, связанными со сном и вниманием [ 7,8,30,53–55,58–62,65]. Базальные ганглии представляют собой другой пример, где контроль бета-колебаний был связан с ритмами отскока между (возбуждающими) субталамическими нейронами и (тормозящими) нейронами внешнего бледного шара (GPe).Большое количество недавних экспериментальных данных также демонстрирует важность кальциевых каналов Т-типа в модуляции этих ритмов. На клеточном уровне экспериментальные данные показывают, что нейроны как STN, так и GPe обладают ионными токами для переключения возбудимости [53,54,64]. На сетевом уровне колебания были зарегистрированы в сети STN-GPe in vitro и in vivo [13,14,39]. Колебания состояния сети, и, более конкретно, когерентности и силы бета-колебаний, были связаны с инициированием произвольных движений как у животных [68,69], так и у людей [14,68,70–73].
Каноническим примером является таламус, где как роль ритмов отскока между (возбуждающими) таламокортикальными нейронами и (тормозящими) ретикулярными нейронами, так и важность кальциевых каналов Т-типа давно признаны в управлении колебательными состояниями сети, связанными со сном и вниманием [ 7,8,30,53–55,58–62,65]. Базальные ганглии представляют собой другой пример, где контроль бета-колебаний был связан с ритмами отскока между (возбуждающими) субталамическими нейронами и (тормозящими) нейронами внешнего бледного шара (GPe).Большое количество недавних экспериментальных данных также демонстрирует важность кальциевых каналов Т-типа в модуляции этих ритмов. На клеточном уровне экспериментальные данные показывают, что нейроны как STN, так и GPe обладают ионными токами для переключения возбудимости [53,54,64]. На сетевом уровне колебания были зарегистрированы в сети STN-GPe in vitro и in vivo [13,14,39]. Колебания состояния сети, и, более конкретно, когерентности и силы бета-колебаний, были связаны с инициированием произвольных движений как у животных [68,69], так и у людей [14,68,70–73]. Экспериментальные исследования показывают предполагаемое увеличение бета-синхронности перед произвольными движениями [74] и связанную с событием десинхронизацию в бета-диапазоне во время движения [70, 72]. Инициирование произвольных движений также связано с увеличением дофамина и, в частности, с временным повышением активности нигростриатных цепей (фазовое высвобождение дофамина) [75,76]. Это временное повышение дофамина запускает снижение когерентности и мощности активности бета-диапазона [77]. Эти наблюдения согласуются с предсказаниями нашей модели при переходном модулирующем воздействии.Колеблющиеся состояния мозга также описаны в коре головного мозга. Модели слоя V коры, вовлеченного в зрение, включают возбуждающе-тормозную сеть и кальциевые токи Т-типа [78]. В этой области мозга осцилляторная активность в альфа-диапазоне (8–12 Гц) блокирует входящие сигналы, подавляя несущественные для задачи области, таким образом направляя сигналы в соответствующие области [79,80]: альфа-колебания обеспечивают функциональное ингибирование и уменьшают возможности обработки.
Экспериментальные исследования показывают предполагаемое увеличение бета-синхронности перед произвольными движениями [74] и связанную с событием десинхронизацию в бета-диапазоне во время движения [70, 72]. Инициирование произвольных движений также связано с увеличением дофамина и, в частности, с временным повышением активности нигростриатных цепей (фазовое высвобождение дофамина) [75,76]. Это временное повышение дофамина запускает снижение когерентности и мощности активности бета-диапазона [77]. Эти наблюдения согласуются с предсказаниями нашей модели при переходном модулирующем воздействии.Колеблющиеся состояния мозга также описаны в коре головного мозга. Модели слоя V коры, вовлеченного в зрение, включают возбуждающе-тормозную сеть и кальциевые токи Т-типа [78]. В этой области мозга осцилляторная активность в альфа-диапазоне (8–12 Гц) блокирует входящие сигналы, подавляя несущественные для задачи области, таким образом направляя сигналы в соответствующие области [79,80]: альфа-колебания обеспечивают функциональное ингибирование и уменьшают возможности обработки.
Механизм сотовой коммутации, отличный от свойств отскока
На клеточном уровне переключатель возбудимости, смоделированный в этой статье, ответственен за выброс отскока и вызванный гиперполяризацией взрыв, два механизма, которые широко наблюдались в экспериментах [7,53–55,58–60].Мы подчеркиваем, что переключение возбудимости отличается от широко изученного постингибиторного отскока (PIR) [7,49,51,52]. Например, кальциевые каналы Т-типа вносят вклад в механизм переключения через их медленную активацию , которая является внутренним источником медленной отрицательной проводимости , в то время как они вносят вклад в постингибиторный отскок за счет своей инактивации , что является собственный источник положительной проводимости . Другие каналы, такие как каналы HCN, только способствуют отскоку, но не влияют на переключение.Важность кальциевых каналов Т-типа уже давно подчеркивается из-за их вклада в механизмы отскока, как в генераторы центральных паттернов, так и в ритмы мозга млекопитающих [7,49,51,52]. Новизна нашей модели в этом отношении заключается в том, чтобы подчеркнуть важность кальциевых каналов Т-типа для их вклада в работу коммутатора в управлении состоянием сети. В отсутствие переключателя одних лишь механизмов восстановления недостаточно для сетевого управления надежными и настраиваемыми сетевыми ритмами. В отсутствие медленной регенерации ритм отскока в возбуждающе-тормозной сети требует специфического резонанса между PIR и кинетикой синаптических связей [41].В этом случае ритм контура уязвим для изменений внутренних свойств нейронов и синаптических связей. Эта хрупкость сильно ограничивает неоднородность ритмов в популяции.
Новизна нашей модели в этом отношении заключается в том, чтобы подчеркнуть важность кальциевых каналов Т-типа для их вклада в работу коммутатора в управлении состоянием сети. В отсутствие переключателя одних лишь механизмов восстановления недостаточно для сетевого управления надежными и настраиваемыми сетевыми ритмами. В отсутствие медленной регенерации ритм отскока в возбуждающе-тормозной сети требует специфического резонанса между PIR и кинетикой синаптических связей [41].В этом случае ритм контура уязвим для изменений внутренних свойств нейронов и синаптических связей. Эта хрупкость сильно ограничивает неоднородность ритмов в популяции.
Конкретное проявление различия между переключением возбудимости и свойством отскока представлено на рис. 5. На этом рисунке возбуждающие клетки демонстрируют всплески как в активном, так и в колебательном состояниях сети. Однако они участвуют в механизме отскока только в колебательном состоянии.Это изменение свойств отскока не влечет за собой изменения связности. Это происходит только в результате переключения между двумя типами возбудимости.
Это происходит только в результате переключения между двумя типами возбудимости.
Механизм переключения среднего поля, не зависящий от свойств связи
Новизна и важность механизма переключения на сетевом уровне заключается в том, что он в значительной степени не зависит от сетевого подключения. В этом отношении наша статья отличается от более ранних компьютерных исследований, в которых изучались модуляции сетевых ритмов посредством модуляции связности [39–41].Общий механизм в этих моделях состоит в том, что более сильная синхронность в популяции зависит от более сильной связи [81]. В таких моделях активных состояний сети связаны с слабой связностью, и асинхронной ритмической активностью, тогда как колебательных состояний связаны с сильной связностью и синхронной ритмической активностью. Вместо этого в настоящей статье переключение между активным и колебательным состояниями происходит без изменения связности.Связность всегда слабая, что позволяет воспроизводить гетерогенные ритмы как в активном, так и в колебательном состоянии. Более сильная связь снижает неоднородность ритмов в популяции. Вместо этого общий сотовый коммутатор позволяет синхронизировать события даже при наличии неоднородности.
Более сильная связь снижает неоднородность ритмов в популяции. Вместо этого общий сотовый коммутатор позволяет синхронизировать события даже при наличии неоднородности.
Сетевые вычислительные модели часто не имеют сотового коммутатора
В большинстве описанных в литературе сетевых вычислительных моделей отсутствует сотовый коммутатор, изучаемый в настоящей статье. Это очевидно для всех моделей, которые фокусируются на синаптической связности и только моделируют скорость или пиковые свойства на клеточном уровне.Но это также относится ко многим моделям, которые учитывают взрывные свойства на клеточном уровне, но не имеют переключателя возбудимости. Даже модели, которые включают кальциевые каналы Т-типа, часто моделируют активацию как мгновенную [82–88]. Эти модели могут моделировать всплески или свойства отскока для определенных значений параметров, но отсутствие медленной отрицательной проводимости делает их хрупкими и не настраиваемыми [42]. Тот факт, что в большинстве моделей не учитывается медленная кинетика активации кальциевых каналов, является дополнительным доказательством вычислительных моделей, которые учитывают механизмы отскока, но не учитывают клеточный переключатель возбудимости. Это показывает, что различию между двумя механизмами не уделялось особого внимания. Это не означает, что моделирование переключения требует больше биофизических деталей, чем моделирование отскока. В недавней статье [89] показано, что для моделирования переключателя достаточно простой модели «интегрировать и запустить» при условии, что она содержит отдельные быстрые и медленные пороги для учета двух различных типов возбудимости. Это также объясняет, почему существующие модели интеграции и включения, которые имеют только один порог, не могут моделировать переключатель возбудимости даже при наличии переменных адаптации.
Это показывает, что различию между двумя механизмами не уделялось особого внимания. Это не означает, что моделирование переключения требует больше биофизических деталей, чем моделирование отскока. В недавней статье [89] показано, что для моделирования переключателя достаточно простой модели «интегрировать и запустить» при условии, что она содержит отдельные быстрые и медленные пороги для учета двух различных типов возбудимости. Это также объясняет, почему существующие модели интеграции и включения, которые имеют только один порог, не могут моделировать переключатель возбудимости даже при наличии переменных адаптации.
Методы
Все моделирование проводилось с использованием языка программирования Julia. Анализ проводился либо в Julia, либо в Matlab. Файлы кода Julia и Matlab находятся в свободном доступе по адресу http://www.montefiore.ulg.ac.be/~guilldrion/Files/Drionetal2018-Code.zip и https://osf.io/k86en .
Однокамерные модели Ходжкина-Хаксли использовались для всех моделей нейронов, следуя уравнению, где I ion соответствует ионным токам, а I app — приложенному извне току. Модель состоит из тока утечки, переходного тока натрия, тока калия выпрямителя с задержкой, кальциевого тока Т-типа, тока калия, активируемого кальцием, и тока катионов, активированных гиперполяризацией, где m представляет переменные активации и h представляет переменные инактивации. Динамика переменных активации и инактивации, зависящих от напряжения, моделируется с помощью уравнения, где и, за исключением τ h , Na ( В ), которое задается как τ hNa ( В, ) = 0.67 / (1 + exp ((V + 62.9) / — 10.0))) * (1.5 + 1 / (1 + exp (V + 34.9) /3.6))). Значения В половина , В наклон , A , B , D и E для каждой переменной приведены в таблице 1, и соответствующие кривые график на рис. 6. Кальций-зависимая активация кальций-активированного калиевого тока моделируется следующим образом: m K , Ca ([ Ca ]) = ([ Ca ] / ( [ Ca ] + K D )) 2 .
Модель состоит из тока утечки, переходного тока натрия, тока калия выпрямителя с задержкой, кальциевого тока Т-типа, тока калия, активируемого кальцием, и тока катионов, активированных гиперполяризацией, где m представляет переменные активации и h представляет переменные инактивации. Динамика переменных активации и инактивации, зависящих от напряжения, моделируется с помощью уравнения, где и, за исключением τ h , Na ( В ), которое задается как τ hNa ( В, ) = 0.67 / (1 + exp ((V + 62.9) / — 10.0))) * (1.5 + 1 / (1 + exp (V + 34.9) /3.6))). Значения В половина , В наклон , A , B , D и E для каждой переменной приведены в таблице 1, и соответствующие кривые график на рис. 6. Кальций-зависимая активация кальций-активированного калиевого тока моделируется следующим образом: m K , Ca ([ Ca ]) = ([ Ca ] / ( [ Ca ] + K D )) 2 .
Рис. 6. Кривые стробирования стационарного канала (слева) и кривые постоянной времени (справа) для различных ионных каналов, присутствующих в модельном нейроне.
Сплошные линии представляют кривые активации. Пунктирные линии представляют кривые инактивации. Постоянные времени показаны в логарифмической шкале.
https://doi.org/10.1371/journal.pcbi.1006125.g006
При моделировании использовались следующие параметры: C м = 1 мкФ / см 2 , V Na = 50 мВ , В K = −85 мВ , В Ca = 120 мВ , В утечка = −59 мВ , V H = −20 мВ .Все максимальные значения проводимости были выбраны случайным образом относительно равномерного распределения в следующих диапазонах (в мСм / см 2 ):,,,,,. Динамика кальция следовала уравнению, в котором k 1 и k 2 также выбирались случайным образом относительно равномерного распределения ( k 1 ϵ [0,075,0,125] и k 2 ϵ [0,0075,0,0125]). В случае мгновенной активации кальциевых каналов Т-типа кальциевый ток Т-типа моделировался как.
Динамика кальция следовала уравнению, в котором k 1 и k 2 также выбирались случайным образом относительно равномерного распределения ( k 1 ϵ [0,075,0,125] и k 2 ϵ [0,0075,0,0125]). В случае мгновенной активации кальциевых каналов Т-типа кальциевый ток Т-типа моделировался как.
Модели нейронов были связаны через соединения AMPA, GABA A и GABA B с использованием следующих уравнений:, и, где AMPA , GABAA и GABAB — переменные, изменение которых зависит от пресинаптического потенциала V до в соответствии с уравнениями« T m ( V до ) = 1 / (1 + exp (- ( V до — 2) / 5)).Синаптические веса были взяты случайным образом с учетом равномерного распределения вокруг центрального значения (). Потенциал реверсии рецептора AMPA был установлен на 0 мВ , GABA Потенциал реверсирования рецептора был установлен на потенциал реверсии хлорида ( В Cl = -70 мВ ), а потенциал реверсирования рецептора GABA B был установлен на обратный потенциал калия ( В K = -85 мВ ). Активация рецептора GABA B считалась в 50 раз медленнее, чем активация рецептора GABA A и AMPA.
Активация рецептора GABA B считалась в 50 раз медленнее, чем активация рецептора GABA A и AMPA.
Динамика потенциала локального поля (LFP) является результатом коллективной синаптической активности популяции нейронов и моделируется нормированной суммой постсинаптических токов. LFP фильтруются на частоте 100 Гц через фильтр Баттерворта четвертого порядка, чтобы отразить использование макроэлектродов при обнаружении LFP. Анализ спектрограммы или частотно-временные графики являются результатом логарифмического представления спектрограммы кратковременного преобразования Фурье LFP. Для кратковременного преобразования Фурье мы рассматриваем частоту дискретизации F s = 1 кГц.
Пространственная кластеризация соединений GABA B была введена путем добавления гауссовского затухания синаптической силы от нейрона i к нейрону j: где g syn — максимальная синаптическая сила, c ij — пространственная постоянная, контролирующая распространение связности (установлено на 0,8), а i , j — позиции нейрон в популяциях E и I. g SD нормализован по пресинаптической популяции, чтобы получить одинаковую общую силу связи для каждого нейрона в постсинаптической популяции.
g SD нормализован по пресинаптической популяции, чтобы получить одинаковую общую силу связи для каждого нейрона в постсинаптической популяции.
Дополнительная информация
S1 Рис. Ответ модельного нейрона с физиологически медленной кинетикой активации кальциевых каналов Т-типа на применение временной гиперполяризации для четырех различных наборов параметров.
Для первого набора параметров (верхний левый график) высвобождение гиперполяризации вызывает генерацию переходного периода всплеска в обеих моделях, свойство, называемое постингибиторным отскоком (PIR). Для второго набора параметров (верхний правый график) высвобождение гиперполяризации вызывает генерацию переходного периода всплеска в модели, имеющей медленную кинетику активации кальциевых каналов Т-типа (слева), свойство, называемое рикошетом (RB).Для третьего и четырех наборов параметров (нижние кривые) гиперполяризация вызывает переключение с медленного тонического всплеска на всплеск, при этом частота внутри всплеска намного выше, чем частота тонического возбуждения, свойство, называемое всплеском, вызванным гиперполяризацией (HIB). Оба двух разных набора параметров генерируют одинаковую частоту медленного тонического возбуждения, вызванное гиперполяризацией переключение на взрыв, но сама частота всплеска сильно различается между ними, показывая, что модуляция ритмической активности не зависит от механизма переключения.
Оба двух разных набора параметров генерируют одинаковую частоту медленного тонического возбуждения, вызванное гиперполяризацией переключение на взрыв, но сама частота всплеска сильно различается между ними, показывая, что модуляция ритмической активности не зависит от механизма переключения.
https://doi.org/10.1371/journal.pcbi.1006125.s001
(EPS)
Благодарности
Авторы выражают благодарность профессору Еве Мардер за полезные комментарии к рукописи.
Ссылки
- 1. Zagha E, McCormick DA (2014) Нейронный контроль состояния мозга. Curr Opin Neurobiol. 29: 178–86. pmid: 25310628
- 2. Бузаки Г. (2006) Ритмы мозга. Нью-Йорк: Издательство Оксфордского университета.
- 3.Ван XJ (2010) Нейрофизиологические и вычислительные принципы корковых ритмов в познании. Physiol Rev. 90: 1195–268. pmid: 20664082
- 4.
Uhlhaas PJ, Singer W (2006) Нервная синхрония при расстройствах мозга: актуальность для когнитивных дисфункций и патофизиологии. Нейрон. 52: 155–68. pmid: 17015233
- 5. Lee SH, Dan Y (2012) Нейромодуляция состояний мозга. Нейрон. 76: 209–22. pmid: 23040816
- 6. McCormick DA, Nusbaum MP (2014) Редакционный обзор: нейромодуляция: настройка свойств нейронов, сетей и поведения.Curr Opin Neurobiol. 29: iv – vii. pmid: 25457725
- 7. Маккормик Д.А., Бал Т. (1997) Сон и возбуждение: таламокортикальные механизмы. Annu Rev Neurosci. 20: 185–215. pmid: 12
- 8. Шерман С.М. (2001) Тонизирующая и импульсная стрельба: двойные режимы таламокортикального реле. Trends Neurosci. 24: 122–6. pmid: 11164943
- 9. Фрис П. (2015) Ритмы познания: общение через согласованность. Нейрон. 88: 220–35. pmid: 26447583
- 10. Song K, Meng M, Chen L, Zhou K, Luo H (2014) Поведенческие колебания внимания: ритмические α-импульсы, опосредованные через θ-диапазон.J Neurosci. 34: 4837–44. pmid: 24695703
- 11.
Кэннон Дж. , Маккарти М.М., Ли С., Ли Дж., Бёргерс С., Уиттингтон М.А., Копелл Н. (2014) Нейросистемы: ритмы мозга и когнитивная обработка. Eur J Neurosci. 39: 705–19. pmid: 24329933
- 12. Lakatos P, Musacchia G, O’Connel MN, Falchier AY, Javitt DC, Schroeder CE (2013) Механизм спектрально-временного фильтра слухового избирательного внимания. Нейрон. 77: 750–61. pmid: 23439126
- 13. Herrojo Ruiz M1, Brücke C, Никулин В.В., Schneider GH, Kühn AA (2014) Колебания амплитуды бета-диапазона во внутреннем бледном глобусе человека поддерживают кодирование границ последовательности во время начального обучения сенсомоторной последовательности.Нейроизображение. 85 Pt 2: 779–93.
- 14. Kühn AA, Williams D, Kupsch A, Limousin P, Hariz M, Schneider GH, Yarrow K, Brown P (2004) Связанная с событием бета-десинхронизация в субталамическом ядре человека коррелирует с двигательными функциями. Мозг 127: 735–46. pmid: 14960502
- 15.
Тимофеев И. , Баженов М. (2005) Механизмы и биологическая роль таламокортикальных колебаний. В: Колумб Ф, редактор, Trends in Chronobiology Research, Nova Science Publishers, Inc., глава 1.С. 1–47.
- 16. McGinley MJ, Vinck M, Reimer J, Batista-Brito R, Zagha E, Cadwell CR, Tolias AS, Cardin JA, McCormick DA (2015) Состояние бодрствования: быстрые изменения модулируют нейронные и поведенческие реакции. Нейрон. 87: 1143–61. pmid: 26402600
- 17. McCormick DA, McGinley MJ, Salkoff DB (2015) Активность, зависящая от состояния мозга в коре и таламусе. Curr Opin Neurobiol. 31: 133–40. pmid: 25460069
- 18. Choi S, Yu E, Lee S, Llinas RR (2015) Измененная таламокортикальная ритмика и связность у мышей, лишенных CaV3.1 Са2 + каналы Т-типа в бессознательном состоянии. Proc Natl Acad Sci U S A 112: 7839–44. pmid: 26056284
- 19.
Шерман С.М., Гиллери Р.В. (1996) Функциональная организация таламокортикальных реле. J Neurophysiol 76: 1367–95. pmid: 88
- 20. Briggs F, Usrey WM (2008) Новые взгляды на кортикоталамическую функцию. Curr Opin Neurobiol. 18: 403–7. pmid: 18805486
- 21. Шерман С.М. (2012) Таламокортикальные взаимодействия. Curr Opin Neurobiol. 22: 575–9.pmid: 22498715
- 22. Константинополь К.М., Бруно Р.М. (2015) Глубокие корковые слои активируются непосредственно таламусом. Наука. 340: 1591–4.
- 23. Golomb D, Wang XJ, Rinzel J (1994) Синхронизирующие свойства колебаний веретена в модели ретикулярного ядра таламуса. J Neurophysiol 72: 1109–26. pmid: 7807198
- 24. Ван XJ, Golomb D, Rinzel J (1995) Возникающие колебания веретена и прерывистая вспышка в таламической модели: специфические нейронные механизмы.Proc Natl Acad Sci USA 92: 5577–81. pmid: 7777551
- 25.
Golomb D, Wang XJ, Rinzel J (1996) Распространение веретенообразных волн в модели таламического среза. J Neurophysiol 75: 750–69. pmid: 8714650
- 26. Sohal VS, Huguenard JR (2002) Взаимное ингибирование контролирует колебательное состояние в таламических сетях. Нейрокомпьютеры 44–46: 653–9.
- 27. Fröhlich F, Bazhenov M, Timofeev I, Steriade M, Sejnowski TJ (2006) Медленные переходы между состояниями устойчивых нейронных колебаний путем зависимой от активности модуляции собственной возбудимости.J Neurosci 26: 6153–62. pmid: 16763023
- 28. McCormick DA, Feeser HR (1990) Функциональные последствия импульсного возбуждения и одиночной спайковой активности в латеральных коленчатых релейных нейронах. Неврология 39: 103–13. pmid: 2089273
- 29. Steriade M (2006) Группировка ритмов мозга в кортикоталамических системах. Неврология 137: 1087–106. pmid: 16343791
- 30. Каин С.М., Снатч Т.П. (2010) Управляемые напряжением кальциевые каналы при эпилепсии. Эпилепсия 51:11.
- 31.
Ситникова Е. (2010) Таламо-корковые механизмы сонных веретен и спайк-волновые разряды в модели абсансной эпилепсии на крысах (обзор). Эпилепсия Res 89: 17–26. pmid: 19828296
- 32. Полак П.О., Фридман Дж., Гольшани П. (2013) Клеточные механизмы модуляции усиления в зависимости от состояния мозга в зрительной коре. Nat Neurosci. 16: 1331–9. pmid: 23872595
- 33. Reimer J, Froudarakis E, Cadwell CR, Yatsenko D, Denfield GH, Tolias AS (2014) Колебания зрачка отслеживают быстрое переключение корковых состояний во время спокойного бодрствования.Нейрон. 84: 355–62. pmid: 25374359
- 34. McGinley MJ, David SV, McCormick DA (2015) Сигнатура потенциала корковой мембраны оптимальных состояний для обнаружения сенсорного сигнала. Нейрон. 87: 179–92. pmid: 26074005
- 35. Мардер Э., Гериц М.Л., Отопалик А.Г. (2015) Устойчивые ритмы схемы в небольших схемах возникают из переменных компонентов и механизмов схемы. Curr Opin Neurobiol. 31: 156–63. pmid: 25460072
- 36.
Marder E, Goaillard JM (2006) Изменчивость, компенсация и гомеостаз нейронной и сетевой функции. Nat Rev Neurosci. 7: 563–74. pmid: 167
- 37. Мардер Э., Бухер Д. (2007) Понимание динамики контура с использованием стоматогастральной нервной системы омаров и крабов. Annu Rev Physiol. 69: 291–316. pmid: 17009928
- 38. Marder E, O’Leary T, Shruti S (2014) Нейромодуляция цепей с переменными параметрами: отдельные нейроны и небольшие цепи раскрывают принципы зависимой от состояния и надежной нейромодуляции. Annu Rev Neurosci. 37: 329–46. pmid: 25032499
- 39.Bevan MD, Magill PJ, Terman D, Bolam JP, Wilson CJ (2002) Двигайтесь в ритм: колебания в сети субталамического ядра и внешнего бледного шара. Trends Neurosci. 25: 525–31. pmid: 12220881
- 40. Destexhe A, Contreras D, Steriade M (1998) Механизмы, лежащие в основе синхронизирующего действия кортикоталамической обратной связи через ингибирование таламических ретрансляционных клеток. J Neurophysiol. 79: 999–1016. pmid: 9463458
- 41.
Destexhe A (1998) Спайк-волновые колебания, основанные на свойствах рецепторов GABAB. J Neurosci 18: 9099–111. pmid: 9787013
- 42. Franci A, Drion G, Sepulcher R (2018). Надежный и настраиваемый всплеск требует медленной положительной обратной связи. J Neurophysiol. 119: 1222–1234 pmid: 29357476
- 43. Drion G, Franci A, Seutin V, Sepulcher R (2012) Новый фазовый портрет возбудимости нейронов. PLoS One. 7: e41806. pmid: 227
- 44. Фрэнси А., Дрион Дж., Сепулчер Р. (2012) Организационный центр в плоской модели нейрональной возбудимости.SIAM J Appl Dyn Syst. 11: 1698–1722.
- 45. Franci A, Drion G, Seutin V, Sepulcher R (2013) Уравнение баланса определяет переключатель возбудимости нейронов. PLoS Comput Biol. 9: e1003040. pmid: 23717194
- 46. Drion G, O’Leary T, Marder E (2015) Вырождение ионных каналов обеспечивает надежную и регулируемую частоту возбуждения нейронов. Proc Natl Acad Sci U S A. 112: E5361–70. pmid: 26354124
- 47.
Фрэнси А., Дрион Дж. , Сепульчер Р. (2014) Моделирование модуляции всплеска нейронов: подход теории сингулярностей.SIAM J Appl Dyn Syst. 13: 798–829.
- 48. Drion G, Franci A, Dethier J, Sepulcher R (2015) Динамические входные проводимости формируют выбросы нейронов. eNeuro. 2 (1).
- 49. Dethier J, Drion G, Franci A, Sepulcher R (2015) Положительная обратная связь на клеточном уровне способствует устойчивости и модуляции на уровне схемы. J Neurophysiol. 114: 2472–84. pmid: 26311181
- 50. Хилле Б. (1984) Ионные каналы возбудимых мембран. Сандерленд, Массачусетс: Sinauer Associates Inc, 426 стр.
- 51. Perkel DH, Mulloney B (1974) Производство моторных паттернов в реципрокно ингибирующих нейронах, демонстрирующих постингибиторный отскок. Наука. 185: 181–3. pmid: 4834220
- 52.
Angstadt JD, Grassmann JL, Theriault KM, Levasseur SM (2005) Механизмы постингибиторного отскока и его модуляция серотонином в возбуждающих двигательных нейронах плавания медицинской пиявки. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 191: 715–32. pmid: 15838650
- 53.Beurrier C, Congar P, Bioulac B, Hammond C (1999) Нейроны субталамического ядра переключаются с одиночной активности на импульсный режим. J Neurosci. 1999, 15 января; 19: 599–609. pmid: 9880580
- 54. Hallworth NE, Wilson CJ, Bevan MD (2003) Чувствительные к апамину калиевые каналы с небольшой проводимостью, активируемые кальцием, благодаря их избирательному соединению с потенциалозависимыми кальциевыми каналами, являются критическими детерминантами точности, темпа и характера генерации потенциала действия у крыс. нейроны субталамического ядра in vitro.J Neurosci. 23: 7525–42. pmid: 12930791
- 55. Маккормик Д.А., Принц Д.А. (1986) Ацетилхолин вызывает взрывную реакцию в ретикулярных нейронах таламуса, активируя калиевую проводимость. Природа. 319: 402–5. pmid: 2418361
- 56.
Griffen TC, Maffei A (2014) ГАМКергические синапсы: их пластичность и роль в сенсорной коре. Front Cell Neurosci. 8:91. pmid: 24723851
- 57. Барберис А., Баччи А. (2015) Редакция: Пластичность ГАМКергических синапсов.Front Cell Neurosci. 9: 262. pmid: 26217186
- 58. Contreras D, Curro Dossi R, Steriade M (1993) Электрофизиологические свойства ретикулярных таламических нейронов кошек in vivo. J Physiol. 470: 273–94. pmid: 8308730
- 59. Astori S, Wimmer RD, Prosser HM, Corti C, Corsi M, Liaudet N, Volterra A, Franken P, Adelman JP, Lüthi A (2011) Кальциевый канал Ca (V) 3.3 является основным кардиостимулятором веретена сна в таламусе. Proc Natl Acad Sci U S A. 108: 13823–8. pmid: 21808016
- 60.Huguenard JR, Prince DA (1992) Новый ток Т-типа лежит в основе длительной Са2 + -зависимой вспышки в габаэргических нейронах ретикулярного ядра таламуса крысы. J Neurosci. 12: 3804–17. pmid: 1403085
- 61.
Cheong E, Shin HS (2013) Са2 + каналы Т-типа в нормальных и аномальных функциях мозга. Physiol Rev. 93: 961–92. pmid: 23899559
- 62. Crunelli V, Cope DW, Hughes SW (2006) Таламические каналы Ca2 + Т-типа и медленный сон. Клеточный кальций. 40: 175–90.pmid: 16777223
- 63. Park YG, Park HY, Lee CJ, Choi S, Jo S, Choi H, Kim YH, Shin HS, Llinas RR, Kim D (2010) Ca (V) 3.1 — кардиостимулятор тремора нижней оливы. Proc Natl Acad Sci U S. 107: 10731–6. pmid: 20498062
- 64. Tai CH, Yang YC, Pan MK, Huang CS, Kuo CC (2011) Модуляция субталамических каналов Ca (2+) T-типа устраняет двигательный дефицит в модели болезни Паркинсона на крысах. J Clin Invest. 121: 3289–305. pmid: 21737877
- 65.Choi S, Yu E, Lee S, Llinas RR (2015) Измененная таламокортикальная ритмичность и связность у мышей, лишенных Ca2 + -каналов CaV3.1 T-типа в бессознательном состоянии. Proc Natl Acad Sci U S A. 112: 7839–44. pmid: 26056284
- 66.
Miwa H, Kondo T (2011) Кальциевый канал T-типа как новая терапевтическая мишень для тремора. Мозжечок. 2011 сен; 10: 563–9. pmid: 21479969
- 67. Пак Ю.Г., Ким Дж., Ким Д. (2013) Потенциальная роль каналов Ca2 + Т-типа в координации движений.Передние нервные цепи. 7: 172. pmid: 241
- 68. Энгель А.К., Фрис П. (2010) Колебания в бета-диапазоне — сигнал о статус-кво? Curr Opin Neurobiol 20: 156–65. pmid: 20359884
- 69. Leventhal DK, Gage GJ, Schmidt R, Pettibone JR, Case AC, Berke JD (2012) Бета-колебания базальных ганглиев сопровождают использование реплик. Нейрон 73: 523–36. pmid: 22325204
- 70. Леви Р., Эшби П., Хатчисон В. Д., Ланг А. Э., Лозано А. М., Достровский Д. О. (2002) Зависимость колебаний субталамического ядра от движения и дофамина при болезни Паркинсона.Мозг 125: 1196–209. pmid: 12023310
- 71.
Brown P, Williams D (2005) Потенциальная активность локального поля базальных ганглиев: характер и функциональное значение для человека. Clin Neurophysiol 116: 2510–9. pmid: 16029963
- 72. Кюн А.А., Дойл Л., Погосян А., Ярроу К., Купш А., Шнайдер Г. Х., Хариз М. И., Троттенберг Т., Браун П. (2006) Модуляция бета-колебаний в субталамической области во время воображения движения при болезни Паркинсона. Мозг 129: 695–706. pmid: 16364953
- 73.Амирновин Р., Уильямс З.М., Косгроув Г.Р., Эскандар Е.Н. (2004) Движения под визуальным контролем подавляют субталамические колебания у пациентов с болезнью Паркинсона. J Neurosci 24: 11302–6. pmid: 15601936
- 74. Дженкинсон Н., Браун П. (2011) Новое понимание взаимосвязи между дофамином, бета-колебаниями и двигательной функцией. Trends Neurosci 34: 611–8. pmid: 22018805
- 75. Намбу А. (2008) Семь проблем на базальных ганглиях. Curr Opin Neurobiol 18: 595–604.pmid: 143
- 76. Jin X, Costa RM (2010) Сигналы пуска / остановки возникают в нигростриатальных цепях во время обучения последовательности. Природа 466: 457–62. pmid: 20651684
- 77. Бриттен Дж. С., Браун П. (2013) Колебания и базальные ганглии: моторный контроль и за его пределами. Нейроизображение.
- 78. Jones SR, Pinto DJ, Kaper TJ, Kopell N (2000) Альфа-частотные ритмы десинхронизируются на больших корковых расстояниях: исследование моделирования. J Comput Neurosci. 9: 271–91.pmid: 11139043
- 79. Jones SR, Kerr CE, Wan Q, Pritchett DL, Hämäläinen M, Moore CI (2010) Приведенное пространственное внимание управляет функционально релевантной модуляцией мю-ритма в первичной соматосенсорной коре. J Neurosci 30: 13760–5. pmid: 20943916
- 80. Дженсен О., Мазахери А. (2010) Формирование функциональной архитектуры за счет колебательной альфа-активности: стробирование путем торможения. Фронт Hum Neurosci 4: 186. pmid: 21119777
- 81. Ferrari FA, Viana RL, Lopes SR, Stoop R (2015) Фазовая синхронизация связанных разрывных нейронов и обобщенная модель Курамото.Нейронная сеть. 66: 107–18. pmid: 25828961
- 82. Ван XJ, Rinzel J (1992) Чередующиеся и синхронные ритмы в реципрокно ингибирующих модельных нейронах. Neural Comput. 4: 84–97.
- 83. Pospischil M, Toledo-Rodriguez M, Monier C, Piwkowska Z, Bal T, Frégnac Y, Markram H, Destexhe A (2008) Минимальные модели типа Ходжкина-Хаксли для различных классов корковых и таламических нейронов. Biol Cybern. 99: 427–41. pmid: 129
- 84. Кубота С., Рубин Дж. (2011) NMDA-индуцированная вспышка в модельном нейроне субталамического ядра.J нейрофизиол. 106: 527–537. pmid: 21562199
- 85. Рубин Дж, Терман Д. (2004) Высокочастотная стимуляция субталамического ядра устраняет патологическую таламическую ритмичность в вычислительной модели. J Comput Neurosci. 16: 211–35. pmid: 15114047
- 86. Rush ME, Rinzel J (1994) Анализ взрыва в модели таламического нейрона. Biol Cybern. 71: 281–91. pmid: 7948220
- 87. Smith GD, Cox CL, Sherman SM, Rinzel J (2000) Фурье-анализ таламокортикальных ретрансляционных нейронов, управляемых синусоидой, и минимальная модель интеграции и запуска или взрыва.J Neurophysiol. 83: 588–610. pmid: 10634897
- 88. Terman D, Rubin J, Yew AC, Wilson CJ (2002) Модели активности в модели для субталамопаллидной сети базальных ганглиев. J Neurosci. 22: 2963–76. pmid: 11923461
- 89. Ван Поттельберг Т., Дрион Дж., Сепулчер Р. (2018) Надежная модуляция моделей интеграции и возгорания. Neural Comput., 30: 987–1011. pmid: 29381445
 Нейрон. 52: 155–68. pmid: 17015233
Нейрон. 52: 155–68. pmid: 17015233 , Маккарти М.М., Ли С., Ли Дж., Бёргерс С., Уиттингтон М.А., Копелл Н. (2014) Нейросистемы: ритмы мозга и когнитивная обработка. Eur J Neurosci. 39: 705–19. pmid: 24329933
, Маккарти М.М., Ли С., Ли Дж., Бёргерс С., Уиттингтон М.А., Копелл Н. (2014) Нейросистемы: ритмы мозга и когнитивная обработка. Eur J Neurosci. 39: 705–19. pmid: 24329933 , Баженов М. (2005) Механизмы и биологическая роль таламокортикальных колебаний. В: Колумб Ф, редактор, Trends in Chronobiology Research, Nova Science Publishers, Inc., глава 1.С. 1–47.
, Баженов М. (2005) Механизмы и биологическая роль таламокортикальных колебаний. В: Колумб Ф, редактор, Trends in Chronobiology Research, Nova Science Publishers, Inc., глава 1.С. 1–47. pmid: 88
pmid: 88 pmid: 8714650
pmid: 8714650 Эпилепсия Res 89: 17–26. pmid: 19828296
Эпилепсия Res 89: 17–26. pmid: 19828296 Nat Rev Neurosci. 7: 563–74. pmid: 167
Nat Rev Neurosci. 7: 563–74. pmid: 167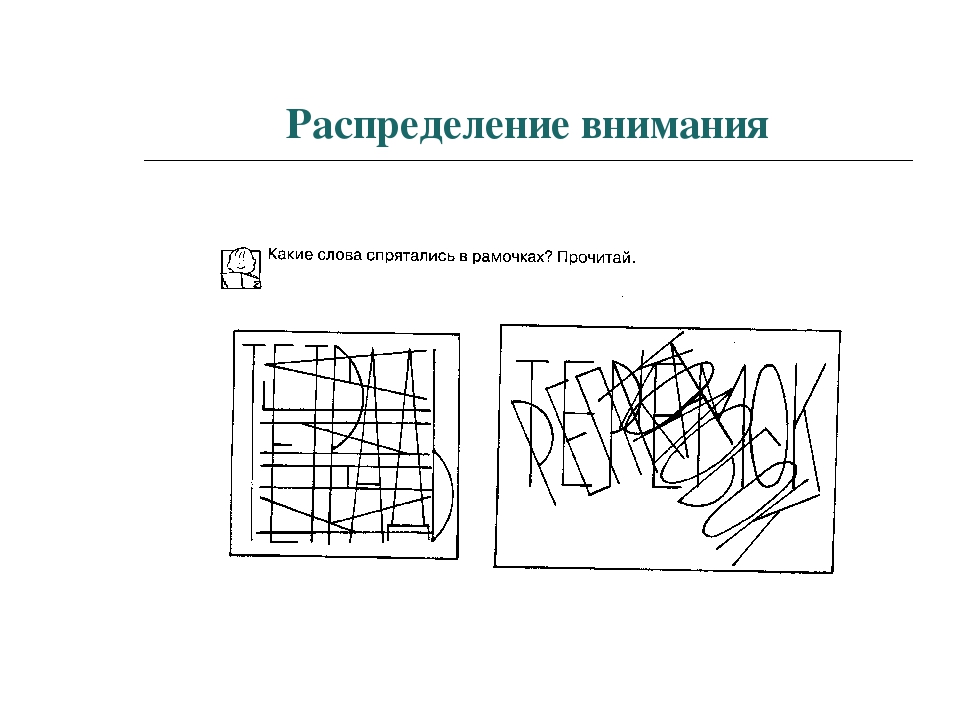 J Neurosci 18: 9099–111. pmid: 9787013
J Neurosci 18: 9099–111. pmid: 9787013 , Сепульчер Р. (2014) Моделирование модуляции всплеска нейронов: подход теории сингулярностей.SIAM J Appl Dyn Syst. 13: 798–829.
, Сепульчер Р. (2014) Моделирование модуляции всплеска нейронов: подход теории сингулярностей.SIAM J Appl Dyn Syst. 13: 798–829.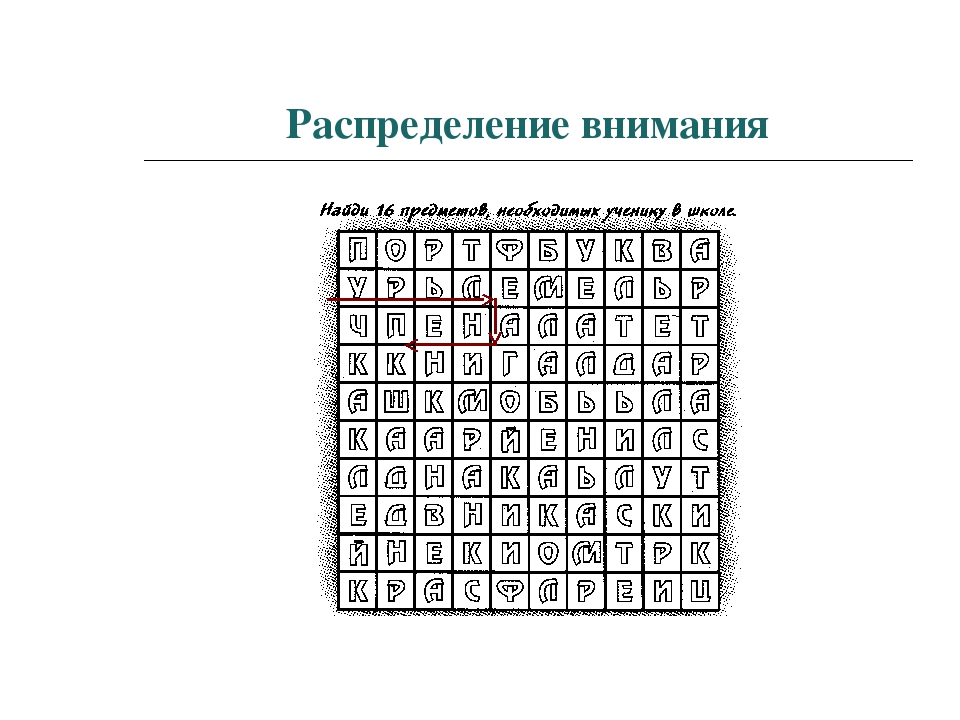 J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 191: 715–32. pmid: 15838650
J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 191: 715–32. pmid: 15838650 Front Cell Neurosci. 8:91. pmid: 24723851
Front Cell Neurosci. 8:91. pmid: 24723851 93: 961–92. pmid: 23899559
93: 961–92. pmid: 23899559 Мозжечок. 2011 сен; 10: 563–9. pmid: 21479969
Мозжечок. 2011 сен; 10: 563–9. pmid: 21479969 pmid: 16029963
pmid: 16029963Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или уточнить у системного администратора.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или уточнить у системного администратора.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Переключаемые органо-неорганические нанокомпозитные материалы, реагирующие на раздражители — Университет Йонсей
TY — JOUR
T1 — Отключаемые на стимулы переключаемые органо-неорганические нанокомпозитные материалы
AU — Lee, Wonsik
Lee 9000 — Kim, Dong
AU — Kim, Dong , SeokheeAU — Park, Jihyeon
AU — Oh, Seoyeah
AU — Kim, Geonho
AU — Lim, Joonyoung
AU — Kim, Jiwon
N1 — Информация о финансировании: Это исследование было поддержано Министерством науки, ИКТ и планирования будущего (MSIP) Кореи в рамках «Творческой программы ICT Consilience» (IITP-2018-2017-0-01015) под руководством Института продвижения информационных и коммуникационных технологий ( IITP), а также Программой фундаментальных научных исследований Национального исследовательского фонда Кореи (NRF-2016-009709).Это исследование также было поддержано научным сообществом TJ Park Фонда POSCO TJ Park Foundation. Работа поддержана Инициативой перспективных исследований Университета Йонсей 2017 г. (2017-22-0076).
PY — 2018/12
Y1 — 2018/12
N2 — Органико-неорганические нанокомпозитные материалы привлекли большое внимание, поскольку они покрывают широкий спектр свойств за счет сочетания контрастирующих свойств и / или создания новых свойств. Функциональными свойствами композитов можно дополнительно управлять с помощью таких стимулов, как свет, pH, магнитное / электрическое поле, механическая сила и тепло.Когда стимулы могут управлять широким спектром свойств материалов, это может быть применено к устройству с системой включения-выключения: переключателю. Переключаемый материал должен демонстрировать свойство, которое заметно отличается при стимуляции (т.е. состояниях ВКЛ и ВЫКЛ) с обратимым механизмом. Активно проводились исследования переключаемых материалов, чтобы максимально увеличить разницу между состояниями ВКЛ и ВЫКЛ путем изменения структуры и состава нанокомпозитов, а также стимулов для контроля времени отклика.Переключаемые материалы находят свое применение в различных областях, таких как катализаторы, сенсоры, фотодетекторы, устройства памяти и системы доставки лекарств. В этом обзоре мы обсуждаем последние достижения в разработке / синтезе органо-неорганических наноразмерных материалов, в которых их свойства можно контролировать в ответ на конкретный стимул. Кроме того, мы представляем различные стимулы (более того, мульти-стимулы), применение переключаемых нанокомпозитов в широком диапазоне областей, а также проблемы с перспективой.
AB — Органико-неорганические нанокомпозитные материалы привлекли большое внимание, поскольку они охватывают широкий спектр свойств за счет сочетания контрастирующих свойств и / или создания новых свойств. Функциональными свойствами композитов можно дополнительно управлять с помощью таких стимулов, как свет, pH, магнитное / электрическое поле, механическая сила и тепло. Когда стимулы могут управлять широким спектром свойств материалов, это может быть применено к устройству с системой включения-выключения: переключателю. Переключаемый материал должен демонстрировать свойство, которое заметно отличается при стимуляции (т.е.е. Состояния ВКЛ и ВЫКЛ) с реверсивным механизмом. Активно проводились исследования переключаемых материалов, чтобы максимально увеличить разницу между состояниями ВКЛ и ВЫКЛ путем изменения структуры и состава нанокомпозитов, а также стимулов для контроля времени отклика. Переключаемые материалы находят свое применение в различных областях, таких как катализаторы, сенсоры, фотодетекторы, устройства памяти и системы доставки лекарств. В этом обзоре мы обсуждаем последние достижения в разработке / синтезе органо-неорганических наноразмерных материалов, в которых их свойства можно контролировать в ответ на конкретный стимул.Кроме того, мы представляем различные стимулы (более того, мульти-стимулы), применение переключаемых нанокомпозитов в широком диапазоне областей, а также проблемы с перспективой.
UR — http://www.scopus.com/inward/record.url?scp=85056699406&partnerID=8YFLogxK
UR — http://www.scopus.com/inward/citedby.url?scp=85056699406&partnerID=8Y
U2 — 10.1016 / j.nantod.2018.10.006
DO — 10.1016 / j.nantod.2018.10.006
M3 — См. Статью
AN — SCOPUS: 85056699406
VL — 23
SP — 97
SP — 97EP — 123
JO — Nano Today
JF — Nano Today
SN — 1748-0132
ER —
Переключаемая электронная проводимость в интеркалированных металлоорганических каркасах
ВВЕДЕНИЕ
Физические свойства Несколько типов металлоорганических каркасов (MOF), которые включают органический линкер и ионы / кластеры металлов, были изучены с использованием настраиваемых и четко определенных пористых образцов, чтобы изучить их применимость в адсорбции и разделении газов ( 1 ).В настоящее время их электрические свойства ( 2 — 4 ), такие как их проводимость ( 5 — 10 ), магнетизм ( 11 , 12 ), фотонные характеристики ( 13 , 14 ) и свойства аккумуляторов энергии ( 15 — 18 ), также исследуются. До недавнего времени сообщалось о различных типах электропроводящих MOF: (i) MOF на основе тетратиафульвалена с одномерными (1D) π-стэкинг-каналами взаимодействия ( 5 , 6 ), (ii) на основе Ni-дитиолена MOF с использованием полупроводников p-типа по окислительно-восстановительному механизму ( 13 ), (iii) MOF на основе дигидрокситерефталата с гексагональной упаковкой с одномерной подвижностью заряда с использованием цепочек металл-кислород (или металл-сера) ( 7 — 9 ), (iv) MOF на основе бензентрикарбоксилата, пропитанные тетрацианохинодиметаном, демонстрирующим настраиваемую электрическую проводимость ( 10 ), и (v) высокопроводящие 2D MOF путем наложения листов с расширенным π-сопряжением ( 19 — 21 ).Эти электропроводящие MOF являются потенциальными кандидатами для будущих моделей реконфигурируемой электроники и датчиков. Они также могут быть полезны в качестве альтернативного подхода в органических электронных устройствах для удовлетворения требований безопасности по термочувствительному переключению при высокой температуре.
Мы недавно сообщили о синтезе материалов электродов с интеркалированным MOF (iMOF) посредством электрохимически обратимой реакции интеркаляции Li с переносом заряда ( 15 — 17 ).Как показано на рис. 1, 2,6-нафталиндикарбоксилат дилития [2,6-Naph (COOLi) 2 ], типичный iMOF, состоит из ароматического дикарбоксилата лития и образует органо-неорганическую слоистую структуру на основе π- уложенные слои насадки нафталина и тетраэдрические LiO 4 единиц соответственно. Он также демонстрирует связанное с двухэлектронным переносом обратимое интеркалирование ионов Li, соответствующее обратимой удельной емкости ок. 220 мА · час g −1 , при постоянном потенциале 0.8 В (по сравнению с Li / Li + ), без растворения в органическом электролите. Этот рабочий потенциал позволяет как поддерживать высокое напряжение, так и подавлять отложение дендритов лития, одновременно создавая внутреннее короткое замыкание для проблем безопасности ( 15 ). Кроме того, 2,6-Naph (COOLi) 2 также проявляет другое интересное свойство, заключающееся в том, что он претерпевает очень небольшое изменение в объеме своей элементарной ячейки (0,33%), сохраняя при этом свой каркас во время интеркаляции Li.Кроме того, порошок этого материала, состоящий из белых частиц микрометрового размера, обладает изоляционными свойствами. Однако этот материал электрохимически полностью обратим при использовании для изготовления электродов; это верно даже при низком содержании углерода ( 17 ). Хотя существует ряд отчетов об ароматических дикарбоксилатных солях, которые похожи на электродные материалы ( 22 — 24 ), эти материалы считаются имеющими плохую электронную проводимость, а также основные механизмы электронной проводимости, ответственные за электрохимическую обратимость. еще предстоит выяснить.Чтобы выяснить причину электрохимической обратимости электродов iMOF, мы провели экспериментальные и теоретические исследования электрических свойств этих электродов. Наши результаты являются прямым доказательством переключающейся электронной проводимости 2,6-Naph (COOLi) 2 при сохранении его ионной проводимости и термостабильности, и предполагают, что эти материалы будут очень подходящими для производства нового класса термочувствительных органических соединений. коммутационные устройства.
Инжир.1 Схематическое изображение окислительно-восстановительного механизма кристаллического 2,6-Naph (COOLi) 2 .Оранжевый интеркалированный Li; зеленый, первозданный Ли; красный, О; синий, C; белый, H.
РЕЗУЛЬТАТЫ
Мы экспериментально охарактеризовали электрохимическое поведение электродов 2,6-Naph (COOLi) 2 (рис. S1A). Из кривых разряда, соответствующих реакции интеркаляции лития, было обнаружено, что первый потенциал разряда этих электродов составляет примерно На 100 мВ ниже, чем во втором и последующих циклах.Как показано на рис. S1B, кривые дифференциальной емкости ( dQ / dV ) показали уменьшение поляризации при повторном циклировании, причем изменение потенциала между первым и вторым циклами разряда было наибольшим. Вольт-амперное сопротивление ( I — В ) после второго цикла было вдвое меньше первого (рис. S1B, вставка). Это наводит на мысль, что уменьшение внутреннего сопротивления в значительной степени связано с интеркаляцией Li, особенно во время первого цикла, и что степень интеркаляции Li достаточно высока, чтобы изменить материал из изоляционного материала в проводящий.
Подробные процессы интеркаляции / деинтеркаляции лития 2,6-Naph (COOLi) электродов 2 были проанализированы с использованием метода гальваностатического периодического титрования (GITT) для исследования причины внутреннего сопротивления (рис. S2A). Потенциал холостого хода (OCP) для начального цикла и после 10 циклов был почти одинаковым, как показано на рис. S3. Что было замечательно, так это различие в поведении поляризации Δ E i , когда ток подавался на OCP (подробное объяснение см. На рис.S2B). Поляризация в исходном состоянии (рис. S3A) была вдвое больше, чем после 10-го цикла (рис. S3B). Точно так же сопротивление падения IR в исходном состоянии (от 200 до 300 Ом) (рис. S3A, вставка), основным компонентом которого было электронное сопротивление с точки зрения постоянной времени, было в два-три раза больше, чем после 10-й цикл (около 100 Ом) (рис. S3B, вставка). Таким образом, можно сделать вывод, что изменение внутреннего сопротивления способствовало электронной проводимости, которая в значительной степени была связана с состоянием интеркаляции Li.
Затем мы выполнили моделирование с использованием расчетов из первых принципов, основанных на теории функционала плотности, чтобы понять механизм электронной зонной проводимости до и после интеркаляции Li. Ранее мы определили две модели для положений интеркалированного Li с помощью измерения дифракции рентгеновских лучей (XRD) и уточнения по Ритвельду с использованием расчетов из первых принципов ( 15 ): В модели 1 интеркалированные ионы Li расположены вблизи тетраэдрические сетчатые слои LiO 4 , тогда как в модели 2 основная структура расположена в π-уложенных слоях нафталина (рис.S4). В исходном состоянии (фиг. 2A) запрещенная зона относительно велика и имеет энергию ( E g ) 3,27 эВ. Согласно моделям 1 и 2, соответствующим интеркаляции Li (рис.2, B и C, соответственно), уровень Ферми перемещается вверх в зону проводимости после интеркаляции Li из-за увеличения электронной плотности из-за органической единицы (рис. . S5). Следовательно, рассчитанные значения E g для модели 1 ( E g = 1,51 эВ) и модели 2 ( E g = 0.99 эВ) ниже, что указывает на то, что интеркаляция Li превращает 2,6-наф (COOLi) 2 из изоляционного материала в полупроводниковый; это особенно характерно для модели 2. Хотя MOF 2D канального типа с высокой проводимостью демонстрирует малую ширину запрещенной зоны ( E g = 0,13–0,23 эВ) ( 21 ), расчетное значение E g Значение для модели 2 относительно ниже, чем значения E g ранее описанных 1D канальных MOF, таких как кубические массивы Zn 4 O (CO 2 ) 6 каркасов типа ( E г = 1.От 3 до 6,8 эВ) ( 25 , 26 ) или цилиндрические каналы с шестигранной насадкой из M 3 O 3 (CO 2 ) 3 каркаса типа ( E г = 1,47 до 2,60 эВ) ( 9 ). CBM исходной модели и CBM модели 1 линейны (красные линии на рис. 2, A и B). С другой стороны, CBM в модели 2 изогнут и имеет дисперсию (рис. 2C). Это говорит о том, что электроны в зоне проводимости в модели 2 делокализованы и что электронная подвижность увеличивается.Рассчитанные 2D и 3D распределения электронной плотности CBM показали следующее: исходная структура локализована внутримолекулярно вдоль направления поперечного сечения слоев упаковки нафталина с π-стопкой (рис. 2, D и G). Напротив, в случае модели 1 интеркалированная Li структура делокализована внутримолекулярно между C1 и C1 ‘(рис. 2, E и H). Согласно модели 2, он делокализован межмолекулярно между C1 и C2 (или C1 ‘и C2’), помимо того, что он внутримолекулярно делокализован (рис.2, Е и I). Это показывает, что в модели 2 интеркаляция лития обеспечивает путь прыжковой проводимости электронов через двумерные π-стопки нафталиновой упаковки органических слоев.
Рис. 2 Прогнозный расчет электронной зонной проводимости.( A, – C ) Расчетные электронные зонные структуры, полученные из функционала HSE06 для исходного образца (A) и образца с интеркалированным литием, соответствующих модели 1 (B) и модели 2 (C). Максимум валентной зоны и минимум зоны проводимости (CBM) представлены в зонных структурах синими и красными линиями соответственно.( D — F ) Трехмерное распределение электронной плотности (полученное с помощью функционала PBEsol) CBM кристаллической структуры 2,6-Naph (COOLi) 2 для исходного образца (D) и интеркалированного Li образец, соответствующий модели 1 (E) и модели 2 (F). Оранжевый, интеркалированный Li; зеленый, первозданный Ли; красный, О; синий, C; белый, H. ( G — I ) 2D-распределение электронной плотности CBM через π-стопочный слой нафталина через атомы углерода C1, C1 ‘, C2 и C2’, соответствующие приведенному выше рисунку для исходного образца (G) и литий-интеркалированный образец, полученный с использованием модели 1 (H) и модели 2 (I).
Для дальнейшего исследования электронной проводимости между соседними молекулами, параметров прыжковой проводимости, включая интеграл переноса ( V ), коэффициент диффузии ( D ) и дрейфовую подвижность прыжков (μ h ) для первичной структуры. и литий-интеркалированный, соответствующий модели 2, были определены на основе теории Маркуса (подробное объяснение см. в разделе «Материалы и методы») ( 27 ). Чтобы иметь возможность описать прыжковый транспорт до и после интеркаляции Li кристалла 2,6-Naph (COOLi) 2 , мы рассмотрели следующие три прыжковых пути, по которым заряд может передаваться только между соседними молекулами, как показано на рис. .3 (A и B): (i) π-сопряженные молекулы, расположенные в слоистой структуре базальной упаковки типа «елочка» (P), (ii) упаковка по базовому краю (T) и (iii) упаковка «ребро-край» через тетраэдрический LiO . 4 слой (L). Интегралы переноса всех трех путей перескока, соответствующих состоянию интеркаляции Li, были больше, чем таковые до интеркаляции Li, как показано на фиг. 3C. Напротив, коэффициент диффузии (рис. 3D) и дрейфовая подвижность прыжков (рис. 3E) для пути T заметно увеличились на три порядка после интеркаляции Li.Кроме того, значения для пути P показали высокие значения, тогда как значения для пути L уменьшились. Эти результаты предполагают, что анизотропная прыжковая проводимость электронов возникает в π-стопочном слое упаковки нафталина вдоль плоского направления b — c , но не в органически-неорганическом стопочном слое вдоль вертикального направления b — c ; это вызывает анизотропный 2D электронный транспорт внутри органического слоя кристалла 2,6-Naph (COOLi) 2 .Более того, эти результаты согласуются с рассчитанным распределением электронной плотности зонной структуры, как показано на рис. 2. Расчеты прыжковой проводимости, выполненные в этой работе, могут быть использованы в качестве инструмента молекулярного дизайна для поиска материалов iMOF с высокой электронной проводимостью.
Рис. 3 Прогнозный расчет прыжковой проводимости электронов.( A и B ) Схемы путей перескока электронов между соседними молекулами для исходного образца (A) и образцов с интеркалированным литием, соответствующих модели 2 (B).Оранжевый, интеркалированный Li; зеленый, первозданный Ли; красный, О; синий, C; белый, H. ( C — E ) Сравнение предсказанного интеграла электронного переноса (C), коэффициента диффузии (D) и дрейфовой подвижности прыжка (E) до и после интеркаляции Li.
Для сбора прямых доказательств электронной проводимости в 2,6-Naph (COOLi) 2 был приготовлен образец, интеркалированный литием, с использованием нафталинида лития, который является известным восстанавливающим агентом лития ( 28 , 29 ), на синтезированном образце проведены измерения электронной проводимости.С восстановительным агентом лития и образцом композитного электрода, не содержащего углерода (см. Фиг. 4A), образец, интеркалированный литием, может быть приготовлен без использования каких-либо электрохимических методов. Образец изменился с белого в исходном состоянии до черного в состоянии с интеркалированным литием (фиг. 4B). Кроме того, дифрактограммы, соответствующие исходному состоянию и состоянию интеркаляции лития, были разными и подтвердили, что соответствующие кристаллические структуры образцов были аналогичны тем, о которых сообщалось ранее (рис.S6) ( 15 ). Это подтвердило, что катализируемое нафталином восстановительное литиирование можно использовать для индуцирования литий-интеркалированного состояния в 2,6-Naph (COOLi) 2 , который в этом состоянии имеет черный цвет. Хотя расслоение происходит, когда нафталин вводится в промежуточный слой, когда другие слоистые материалы, такие как графит ( 29 ) и халькогениды ( 30 ), вступают в реакцию с нафталинидом лития, реакция 2,6-нафталина (COOLi) № 2 и нафталинид лития не вызывали расслоения кристаллической структуры, которое приводило к образованию аморфного нафталина.
Рис. 4 Экспериментальное подтверждение переключаемой электронной проводимости.( A ) Схема химического восстановительного литиирования 2,6-Naph (COOLi) 2 с использованием нафталинида лития. ( B ) Фотографии 2,6-Naph (COOLi) 2 электрода без проводящего углерода до (слева) и после (справа) погружения в раствор нафталинида лития / тетрагидрофурана (THF). ( C — E ) Сравнение кривых IV (C), графиков Боде (D) и графиков Найквиста (E) для 2,6-Naph (COOLi) 2 образцов до (синий) и после (красный) погружают в тот же раствор.В (E) вставка представляет собой эквивалентную схему для смешанного электронного и ионно-проводящего материала. Пересечения высоких (100 кГц) и низких (1 Гц) частот с действительной осью представляют собой параллельные смешанные электронные и ионные сопротивления ( R e R i / R e + R i ) и единственное электронное сопротивление ( R e ) соответственно. ( F ) Графики Аррениуса для каждой проводимости, полученные из кривых I-V и EIS при различных температурах (рис.S10), σ e (красный) и σ i (зеленый), которые рассчитаны с использованием модели эквивалентной схемы, показанной на вставке (E) для смешанного электронного и ионно-проводящего материала, и измерения поляризации, σ I — V (синий). Все значения проводимости были рассчитаны с использованием теории перколяции (см. Материалы и методы).
Кривые I-V электродов образца, интеркалированного литием, с использованием закрытой ячейки с параллельными электродами (рис.S7) были линейными, как показано на фиг. 4C, что указывает на то, что образец с интеркалированным литием показал благоприятный отклик по току при приложенном напряжении смещения 0,5 В; Напротив, в исходном состоянии образец показал незначительный отклик по току. Чтобы исследовать подробные электрические характеристики, мы провели спектроскопию электрохимического импеданса (EIS) с использованием той же ячейки. На графиках Боде, показанных на фиг. 4D, образец, интеркалированный литием, показал ок. 10 5 Ом. Это значение на четыре порядка ниже, чем у исходного образца (ок.10 9 Ом), так как частота становится ниже. На графиках Найквиста, показанных на рис. 4E, в исходном образце наблюдали блокирующее поведение, соответствующее только ионной проводимости без электронной проводимости, а в образце с интеркалированным литием наблюдали вдавленный полукруг, что означает смешанную электронную и ионную проводимость (рис. S8) ( 31 — 33 ). Поскольку низкочастотная составляющая сопротивления образца с интеркалированным литием соответствует электронному сопротивлению ( R e ), наблюдаемая токовая характеристика образца с интеркалированным литием, показанная на рис.4C связано с уменьшением R e , предполагая, что переключаемый путь электронной проводимости образован интеркаляцией Li 2,6-Naph (COOLi) 2 . В состояниях с белым и черным цветами (рис. 4B) 2,6-Naph (COOLi) 2 проявлял изолирующие и проводящие характеристики соответственно. Результаты оптической эмиссионной спектроскопии с индуктивно связанной плазмой (ICP-OES) показали, что количество интеркалированного Li можно контролировать с помощью времени обработки нафталинидом лития; кроме того, электронная проводимость улучшалась в зависимости от количества интеркалированного Li (рис.S9).
Как кривые I-V , так и графики Найквиста для образца, интеркалированного литием, показали более низкое сопротивление при повышении температуры (рис. S10). Графики Найквиста для образца с интеркалированным литием показали вдавленный полукруг при всех температурах. Объемная электронная проводимость (σ e ) и ионная проводимость (σ i ) были рассчитаны отдельно от графиков Найквиста наивысшего и наименьшего значений с использованием модели эквивалентной схемы для смешанных электронных и ионных проводников (рис.S8B). Полученные значения σ e и σ i составили от 10 −6 от до 10 −5 См см −1 и 10 −5 от до 10 −4 См см −1 в пределах температуры диапазон от 25 ° до 60 ° C соответственно. Значение σ e согласуется с рассчитанным по кривым I-V (σ I — V ). Полученное σ e сопоставимо с проводящими MOF, такими как гексагонально упакованные цилиндрические каналы M 3 O 3 (CO 2 ) 3 -тип каркасов (от 10 −16 до 10 — 6 См −1 ) ( 9 ), каркасы на основе тетратиафульвалена (10 −6 -10 −4 См −1 ) ( 6 ), каркасы на основе тетрацианохинодиметана (10 −5 от до 10 −1 См см −1 ) ( 34 ) и каркасы на основе дитиолена (10 −11 до 10 −5 См см −1 ) ( 13 ).Кроме того, по сравнению с σ e (от 10 −8 до 10 −4 См см −1 ) и σ i (от 10 −5 до 10 −3 См −1 ) из нетронутых электродных материалов на основе оксидов для батарей, которые известны как смешанные электронные и ионные проводники ( 33 , 35 , 36 ), предлагаемый материал имеет сопоставимые электронные и ионные проводники. Как σ e , так и σ и демонстрируют поведение Аррениуса (рис.4F). Энергии активации ( E a ) для σ e и σ i , рассчитанные из графиков Аррениуса, составляют примерно 18,3 кДж моль -1 (или около 190 мэВ) и около 3,5 кДж моль -1 (или примерно 36 мэВ) соответственно. Хотя E a для σ e немного больше, чем в предыдущих отчетах о высокоподвижных проводящих материалах MOF ( 10 , 20 ), это значение на порядок меньше, чем у нетронутые электродные материалы ( 36 , 37 ). E a для σ i был ниже, чем у ионных проводников хорошо известных твердых электролитов ( 38 — 40 ). Низкий E a обеспечивает наименьшие колебания ионной проводимости и, следовательно, способствует практическому применению в широком диапазоне рабочих температур.
Когда образец использовался в качестве электродного материала, ожидалось, что исчезновение его электронной проводимости во время процесса повышенной температуры примерно при 200 ° C будет действовать как выключатель для безопасности батареи, потому что его изоляционная природа повысит безопасность во время теплового разгона.Чтобы проверить это явление, мы измерили дифрактограммы образца с интеркалированным литием, полученного во время нагрева (рис. 5А). Результаты показали, что кристаллическая структура, демонстрирующая электронную проводимость, остается неизменной примерно до 180 ° C. После этого кристаллическая структура изменилась на исходную (синий узор на рис. S6), демонстрируя изолирующие характеристики через область двухфазного сосуществования в диапазоне температур от 180 ° до 250 ° C, где исходная структура с изолирующими свойствами сохранялась до тех пор, пока температура 400 ° C.Кроме того, прямое свидетельство исчезновения электронной проводимости выше 200 ° C было предоставлено EIS и кривой I-V интеркалированного литием образца после нагревания (рис. 5, B — D). В нагреваемом образце при 100 ° C, который поддерживает кристаллическое состояние с интеркалированным литием, хотя небольшое увеличение сопротивления было подтверждено на низкой частоте из графиков Боде (рис. 5B), токовый отклик наблюдали по кривой IV ( Рис. 5C). На графике Найквиста в низкочастотном диапазоне появился новый полукруг (рис.5D). Это указывает на наличие сопротивления границ зерен за счет роста Li-содержащей пленки, которая образовалась во время химически восстановительного литирования на границе зерен (рис. S9). Эквивалентную схему этого графика Найквиста можно пояснить на рис. S8C ( 32 ). В образце, нагретом до 100 ° C, почти не происходит изменения объемного электронного сопротивления при создании сопротивления границ зерен. Напротив, в нагреваемом образце при 200 ° C, который возвращается в исходное кристаллическое состояние, из-за поведения импеданса было подтверждено увеличение электронного сопротивления, и токовый отклик кривой I-V резко исчез.
Рис. 5 Включение / выключение характеристик электронного транспорта с учетом нагрева.( A ) Температурная зависимость рентгеновских дифрактограмм in situ на порошке 2,6-наф (COOLi), интеркалированного литием. 2 для температур до 400 ° C. ( B — D ) Изменение графиков Боде (B), кривых IV (C) и графиков Найквиста (D) для исходных, интеркалированных литий образцов и образцов, интеркалированных литием, полученных термообработкой при температурах 100 ° и 200 ° C.
Чтобы исследовать Li, который исчез из основной структуры во время термообработки, мы сравнили рентгенограммы исходного нелегированного образца и образца, легированного литием после нагрева, как показано на рис. S11. В образце с интеркалированным литием после нагрева в области больших углов появилось несколько новых пиков, соответствующих LiO, Li 2 O, Li 2 O H 2 O и Li 2 CO 3 . Эти пики появлялись при температурах выше 200 ° C, соответствующих началу выделения Li (рис.S12). Это Li-содержащие компоненты, которые переходят в стойкую фазу. Таким образом, результаты XRD предполагают, что Li, высвобождающийся из образца, легированного литием во время нагрева, включается в Li-содержащие компоненты и становится компонентом сопротивления границ зерен. Кроме того, в образце, интеркалированном литием, после нагрева, хотя рефлекс (011), соответствующий неорганической тетраэдрической сетке LiO 4 ( 16 ), не изменился, рефлекс (102), соответствующий π-стопке нафталиновой упаковки слой смещен на нижний угол.Этот результат означает увеличение расстояния π-стопки нафталина и, таким образом, ингибирование прыжковой проводимости основной структуры. Таким образом, термообработка образца, легированного литием, включает следующие механизмы блокировки трехэлектронной проводимости: (i) исчезновение основной кристаллической структуры, ответственной за прыжковую проводимость, сопровождающую высвобождение Li, (ii) ингибирование прыжковой проводимости, сопровождающее распространение расстояния между пакетами нафталина в основной кристаллической структуре и (iii) формирование Li-содержащего изолирующего слоя с высвобожденным Li и нагревание на границе зерен.Этот электронный механизм блокировки проводимости будет полезной функцией отключения во время теплового разгона. Хотя существует множество косвенных подходов к безопасности аккумуляторных батарей с использованием электролита ( 41 , 42 ), сепараторов ( 43 ) и периферийных материалов ( 44 , 45 ), предлагаемый материал рассматривается как прямой подход. к безопасности электрода, хранящего электрическую энергию.
МАТЕРИАЛЫ И МЕТОДЫ
Синтез образца
Ранее сообщалось о синтезе 2,6-Naph (COOLi) 2 ( 15 ).2,6-Наф (COOLi) 2 синтезировали из моногидрата гидроксида лития (4,85 г, 115,6 ммоль) и 2,6-нафталиндикарбоновой кислоты (10,0 г, 46,3 ммоль) нагреванием с обратным холодильником в 300 мл метанола. Решение было изначально ясным; однако белый осадок образовывался в течение 30 мин. Суспензию перемешивали при кипячении с обратным холодильником в течение 12 часов. Жидкость в суспензии выпаривали, и полученное твердое вещество промывали метанолом и сушили в условиях окружающей среды.Полученный продукт имел форму игольчатых белых кристаллов [выход 96,8% на основе 2,6-Naph (COOLi) 2 ].
Химическая интеркаляция лития
Раствор нафталинида лития (0,1 моль дм -3 ) был приготовлен с использованием ранее описанной процедуры ( 28 , 29 ). Нафталин растворяли в безводном ТГФ, чтобы приготовить 0,1 М раствор, и эквимолярное количество лития добавляли к 0,1 М раствору нафталина при перемешивании при комнатной температуре в перчаточном боксе, заполненном аргоном.После добавления лития раствор стал темно-зеленым. Количество Li-элемента в исходном и восстановленном образцах было подтверждено методом ICP-OES (CIROS 120EOP, Rigaku и SPECTRO).
Порошок in situ XRD
Рентгенограмма интеркалированного литием 2,6-Naph (COOLi) 2 образца, не содержащего проводящего углерода, была получена с использованием камеры Дебая-Шеррера на канале связи BL5S2, Центр синхротронного излучения в Аичи, Префектура Айти, Япония (предложение № 201601031). Данные регистрировались детектором PILATUS 100K (DECTRIS Ltd.) с разрешением 1.00011 Å (калибровка с использованием стандартного порошка CeO 2 ) для значений 2θ от 10 ° до 24 °; размер шага 0,01 °. Образец нагревали с помощью высокотемпературного газодувки N 2 со скоростью 5 ° C мин. -1 до температур до 400 ° C, и рентгенограмма регистрировалась каждые 10 ° C после времени ожидания. 2 мин для каждой температуры. Перед измерениями in situ XRD был приготовлен образец 2,6-Naph (COOLi) 2 с интеркалированным литием путем пропитывания электрода 2,6-Naph (COOLi) 2 от проводящего углерода в нафталине лития / THF. решение (0.1 моль дм −3 ) в перчаточном ящике, наполненном аргоном. Электроды 2,6-Naph (COOLi) 2 , не содержащие углерода в качестве проводящего агента, также были приготовлены путем диспергирования 98% (масс.) 2,6-Naph (COOLi) 2 и 2% масс. Карбоксиметилцеллюлозы (CMC). ) в воде, а затем путем нанесения суспензии на медную фольгу; скорость нагружения электрода составляла ок. 3 мг / см −2 . После изготовления электрода образец снимали с электрода, помещали в капиллярную трубку и герметично закрывали для предотвращения окисления влажным воздухом во время измерений XRD, которые проводились в перчаточном боксе, заполненном аргоном.
Электрохимические измерения
Измерения гальваностатического заряда / разряда и GITT проводились с использованием двухэлектродной системы ячеек Li / 2,6-Naph (COOLi) 2, собранных в перчаточном боксе, заполненном аргоном. Электроды из 2,6-наф (COOLi) 2 были приготовлены путем нанесения покрытия из дисперсии из 81 мас.% 2,6-наф (COOLi) 2 , 14 мас.% Углеродной сажи в качестве проводящего агента, 2 мас.% КМЦ в виде связующее и 3 мас.% бутадиенстирольного каучука (также используемого в качестве связующего) в воде на медной фольге.Скорость нагружения электродов составляла ок. 3 мг / см −2 . В качестве электролита использовали 1 М LiPF 6 , растворенный в растворе этиленкарбоната, диметилкарбоната и этилметилкарбоната (объемное соотношение 30:40:30). В качестве разделителя использовалась микропористая полипропиленовая пленка. Во время измерений гальваностатического заряда / разряда для определения удельной емкости и электрохимической обратимости электродов 2,6-Naph (COOLi) 2 при 25 ° C, элементы монетного типа циклически менялись между 0.5 и 2,0 В (по сравнению с Li / Li + ) со скоростью, соответствующей полной зарядке до теоретической емкости 2,6-Naph (COOLi) 2 за 10 часов.
Электрические измерения
Все электронные транспортные характеристики были выполнены с использованием закрытой ячейки с параллельными электродами с четырехэлектродной системой (12962A, Solartron). Сопротивление ячейки без образца на семь-восемь порядков меньше по сравнению с ячейкой с образцом, поэтому его можно игнорировать. Характеристики I-V и EIS были измерены при 25 ° C с помощью системы Solartron MultiStat (1470E, Solartron) с использованием электродов из 2,6-Naph (COOLi) 2 , не содержащих проводящий углерод; измерения проводились как до, так и после погружения электродов в раствор нафталинида лития / ТГФ.После удаления погруженного раствора электрод промывали ТГФ. Перед измерением электроды просушивали. Затем были приготовлены закрытые ячейки с параллельными гранулами с электродами в сухом состоянии (рис. S7). Электрическое соединение было выполнено с помощью куска медной фольги. Кривые I-V были измерены при скорости сканирования 100 мВ с -1 в диапазоне поляризации от -0,5 до 0,5 В с использованием двухэлектродной системы, собранной в перчаточном боксе, наполненном аргоном. Измерения EIS той же ячейки проводились на OCP.Частота варьировалась от 100 кГц до 1 Гц с амплитудой возмущения 0,5 В (от пика до пика). Измерения проводились при температуре от –30 ° до 60 ° C. Перед измерением образцов, подвергнутых термообработке, образец с интеркалированным литием был отожжен при температурах 100 ° и 200 ° C в течение 5 мин в атмосфере Ar.
Теория перколяции в электронных свойствах
Эффективная электронная проводимость (σ eff ) композитов проводник-изолятор определяется следующим образом (1) где R x , L и S — сопротивление, толщина составного электрода и геометрическая площадь электрода соответственно.Фактическая проводимость может быть оценена из теории перколяции ( 46 — 48 ). Согласно теории перколяции, эффективная электронная проводимость (σ eff ) композитов проводник-изолятор может быть выражена следующим образом (2) где σ 0 — фактическая проводимость, p — массовая доля проводящего материала. , p c — критическая массовая доля на пороге перколяции, а n — критический показатель, который зависит от топологической размерности перколяционной системы.Расчетное значение p для этого композитного электрода в данном исследовании составляет прибл. 0,466. Значение p c композитов проводник-изолятор составляет 0,32 ( 49 , 50 ). В случае модели случайных пустот, в которой изолирующие проницаемые сферы случайным образом погружены в проводящий континуум, показатель степени n оказывается равным 2,38 ( 49 ).
Кинетическая интерпретация индивидуальных проводимостей 2,6-Naph (COOLi), интеркалированного литием
2 композитных электродовДля определения кинетических параметров композитных электродов энергии активации каждой проводимости были оценены с использованием уравнения Аррениуса (3 ) где A , E a , R и T — частотный коэффициент, энергия активации, газовая постоянная и абсолютная температура, соответственно.
Расчет электронной зонной структуры и плотности состояний
Расчеты из первых принципов для структурной оптимизации и анализа электронных свойств были выполнены с помощью пакета моделирования Vienna ab initio ( 51 ) с использованием метода волн с проектором. Точки отсечки для кинетической энергии и расстояния между точками к были установлены на 500 эВ и приблизительно 0,1 Å -1 , соответственно. При оптимизации кристаллических структур гауссово размытие шириной 0.В качестве интеграла по зоне Бриллюэна было принято 2 эВ. Для обменно-корреляционной энергии в рамках теории функционала плотности был применен функционал PBEsol ( 52 ), который пересматривает функционал приближения обобщенного градиента Пердью-Берке-Эрнцерхофа для улучшения равновесных свойств твердых тел и поверхностей. Для расчета электронной структуры использовались метод тетраэдров с поправками Блохля и функционал HSE06 ( 53 , 54 ) для интеграла по зоне Бриллюэна и обменно-корреляционной энергии соответственно.
Расчет прыжковой проводимости электронов с использованием теории Маркуса
Теория Маркуса широко использовалась для расчета прыжковой подвижности в органических полупроводниках ( 27 , 55 , 56 ). Скорость перескока электронов между соседними молекулами ( W, ) может быть определена на основе теории Маркуса, как показано (4), где V — интеграл переноса между начальным и конечным состояниями (в электрон-вольтах), λ — энергия реорганизации (которая определяется как изменение энергии, связанное с релаксацией геометрии во время переноса заряда) — это постоянная Планка, а k B — постоянная Больцмана.Коэффициент диффузии ( D ) вычисляется из скоростей прыжков (см 2 с -1 ) как (5) где n — размерность, i представляет собой конкретный путь прыжка, r i — это расстояние прыжка между двумя молекулами, а P i — прыжковая подвижность, которая рассчитывается по формуле (6)
Затем можно оценить дрейфовую подвижность прыжка (μ h ). из соотношения Эйнштейна (см 2 В −1 с −1 ) (7) где e — заряд электрона.
Интеграл передачи ( В, ) был рассчитан с использованием метода поправки на энергию участка ( 57 ) и подхода DiPro ( 58 ). Энергия реорганизации (λ) была получена из суммы следующих двух разностей энергий ( 56 ) (8) (9) (10) где — энергия заряженной молекулы в равновесной нейтральной (заряженной) геометрии, и составляет нейтральной молекулы в равновесной нейтральной (заряженной) геометрии. Расчеты интеграла переноса и энергии реорганизации были выполнены с использованием пакета Gaussian 09 ( 59 ) с функционалом B3LYP и базисом 6-311 ++ G * для C 12 H 6 O 4 Li 2 (Li 2 ) молекул, извлеченных из экспериментальной кристаллической структуры.
Благодарности: Мы благодарим S. Kosaka за измерения ICP-OES, а также H. Nakano и M. Ohashi за термообработку образцов. Измерения XRD проводились на канале BL5S2, Центр синхротронного излучения Айти, префектура Аити, Япония (предложение № 201601031). Финансирование: Эта работа была поддержана Toyota Central R&D Laboratories Inc. Вклад авторов: Н. Огихара задумал, спроектировал, провел эксперименты и подготовил рукопись.Ю.К. выполнили измерения XRD. Н. Охба выполнил расчеты зонной структуры и прыжковой проводимости. Конкурирующие интересы: Авторы заявляют, что у них нет конкурирующих интересов. Доступность данных и материалов: Все данные, необходимые для оценки выводов в статье, представлены в статье и / или дополнительных материалах. Дополнительные данные, относящиеся к этой статье, могут быть запрошены у авторов.
От макро- к наномасштабу: влияние конечных размеров на переключаемость металлоорганических каркасов
Основные моменты
Переключаемые металлоорганические каркасы (MOF) завоевывают передовые области применения в хранении энергии, зондировании, разделении газов, катализе и биомедицине.Они извлекают выгоду из уникальных адсорбционных характеристик и чувствительности, что приводит к аномально высокой селективности разделения и полезной емкости хранения.
Наноструктурирование обеспечивает эффективное средство адаптации отклика переключаемых MOF. Характерное давление переключения зависит от критических размеров кристалла.
Понимание эффектов критического размера, влияющих на кооперативные преобразования, реагирующие на стимулы, находится в зачаточном состоянии.Термодинамические и кинетические факторы определяют возможность переключения в зависимости от размера. Общие модели, объясняющие связанные с размером изменения чувствительности, требуют дальнейшей методологической разработки. Развитие вычислительных и аналитических инструментов имеет решающее значение для достижения более глубокого понимания поверхностных, межфазных и конечных эффектов.
Переключаемые металлоорганические каркасы (MOF) выделяются для потенциальных применений в хранении, разделении, зондировании и катализе. Понимание механизмов переключения MOF значительно улучшилось за последние два десятилетия.Наноструктурирование необходимо для интеграции таких материалов в тонкие пленки, иерархические композиты и мембраны, а также для биологических приложений. Однако уменьшение размеров ниже критических вызывает резкие изменения в динамическом поведении и реакции на внешние раздражители. Мы обсуждаем наиболее важные экспериментальные результаты и выводим общие рекомендации и гипотезы, касающиеся влияния размера кристалла на возможность переключения. Понимание термодинамики наноструктур и ее последствий для настройки динамических пористых систем требует междисциплинарного подхода, передовых методов определения физических характеристик и новых стратегий моделирования для охвата более широкого диапазона масштабов времени и длины.
Ключевые слова
металлоорганических каркасов
наноструктурирование
переключаемость
термодинамика
Рекомендуемые статьиЦитирующие статьи (0)
© 2021 Автор (ы).