Функция Month (Visual Basic для приложений)
- Чтение занимает 2 мин
В этой статье
Возвращает значение Variant (Integer), определяющее целое число в интервале между 1 и 12 включительно и представляющее месяц года.
Синтаксис
Месяц(дата)
Аргументом обязательной даты является любое вариант,числовая экспрессия, строковая экспрессия илилюбое сочетание, которое может представлять дату. Если аргумент date содержит Null, возвращается значение Null.
Примечание
Если параметр свойства Calendar григорианский, возвращенный integer представляет григорианский день недели для аргумента даты.
Пример
В этом примере функция Month используется для получения значения месяца из заданной даты. В среде разработки буквальная дата отображается в кратком формате даты с помощью параметров локального кода.
Dim MyDate, MyMonth MyDate = #February 12, 1969# ' Assign a date. MyMonth = Month(MyDate) ' MyMonth contains 2.
См. также
Поддержка и обратная связь
Есть вопросы или отзывы, касающиеся Office VBA или этой статьи? Руководство по другим способам получения поддержки и отправки отзывов см. в статье Поддержка Office VBA и обратная связь.
Определение онкопротеина р16ink4a
Иммуноцитохимическое исследование, позволяющее выявить экспрессию белка р16 (p16INK4a) на поверхности эпителиальных клеток. Белок р16 является биологическим маркером начала канцерогенеза – повышение его экспрессии наблюдается при предраковых изменениях и может рассматриваться как непрямой маркер активной онкогенной экспрессии ВПЧ высокого онкологического риска (выявляется при переходе ВПЧ из эписомальной формы в интегрированную). Определение экспрессии белка р16 позволяет с высокой точностью обнаружить риск развития рака шейки матки, своевременно поставить диагноз и назначить лечение. Определение уровня экспрессии позволяет выявить степень нарушения пролиферации, сопутствующей опухолевому росту, способность опухоли к инвазии и метастазированию.
Синонимы русские
Белок p16INK4a; иммуноцитохимическое исследование.
Синонимы
Оncoprotein p16INK4ain; immunocytochemical study.
Метод исследования
Иммуноцитохимический метод + жидкостная цитология.
Какой биоматериал можно использовать для исследования?
Аспират из полости матки.
Общая информация об исследовании
Одной из самых современных методик ранней диагностики предраковых состояний является иммуноцитохимическое исследование — определение онкопротеина р16ink4a для оценки потенциала дисплазии эпителия.
Было доказано, что экспрессия онкопротеина р16ink4a связана с низкой, умеренной и тяжелой дисплазией (по цитологической классификации ВОЗ (CIN I, CIN II и CIN III) и цитологической классификации Бетесда (ASCUS, LSIL и HSIL), причем экспрессия онкопротеина р16ink4a не встречалась в плоском эпителии без признаков дисплазии. В норме кодируемый геном-супрессором белок p16INK4a блокирует вызываемую фактором роста стимуляцию деления клетки за счет угнетения циклин-зависимой киназы, что приводит к нарушению фосфорилирования белка ретинобластомы (БРБ).
В то же время, хотя отмечается четкая корреляция обнаружения ДНК вируса папилломы человека с дисплазиями, имеется также большое количество ПЦР-позитивных по HPV случаев (с эписомальной локализацией HPV), в которых при последующем гистологическом исследовании отсутствует предрак и рак. Это подтверждается негативной иммуноцитохимической реакцией на р16ink4a в диспластических клетках.
В данном исследовании применяется современная технология приготовления цитопрепаратов — жидкостная цитология. Важная особенность метода, повышающая качество исследования: исследуемый материал берется в специальный стабилизирующий раствор, который обеспечивает его сохранность без разрушения и потери клеток. При этом весь клеточный материал сохраняет без изменения свои морфологические и иммуноцитохимические свойства. В последние пять лет в разных странах проводилось много исследований, в которых сравнивалась эффективность традиционной техники и жидкостной цитологии с использованием для подтверждения диагнозов гистологической экспертизы как «золотого стандарта» и оценки цитопрепаратов согласно классификации TBS (The Bethesda System). Метод жидкостной цитологии признан надежным лабораторным тестом, который снижает количество ложноотрицательных результатов, неудовлетворительных для анализа препаратов и время, необходимое цитологу для оценки препарата.
При этом весь клеточный материал сохраняет без изменения свои морфологические и иммуноцитохимические свойства. В последние пять лет в разных странах проводилось много исследований, в которых сравнивалась эффективность традиционной техники и жидкостной цитологии с использованием для подтверждения диагнозов гистологической экспертизы как «золотого стандарта» и оценки цитопрепаратов согласно классификации TBS (The Bethesda System). Метод жидкостной цитологии признан надежным лабораторным тестом, который снижает количество ложноотрицательных результатов, неудовлетворительных для анализа препаратов и время, необходимое цитологу для оценки препарата.
Материал для цитологического исследования получают с поверхности слизистой оболочки. Слизь, присутствующая во взятом материале, мешает перенести на мазок клетки, также материал невозможно равномерно перемешать. При переносе материала на стекло традиционным способом клетки области шейки матки могут не попасть в препарат, подсушивание и потеря прилипших к инструменту клеток значительно снижает диагностическую информативность микропрепаратов.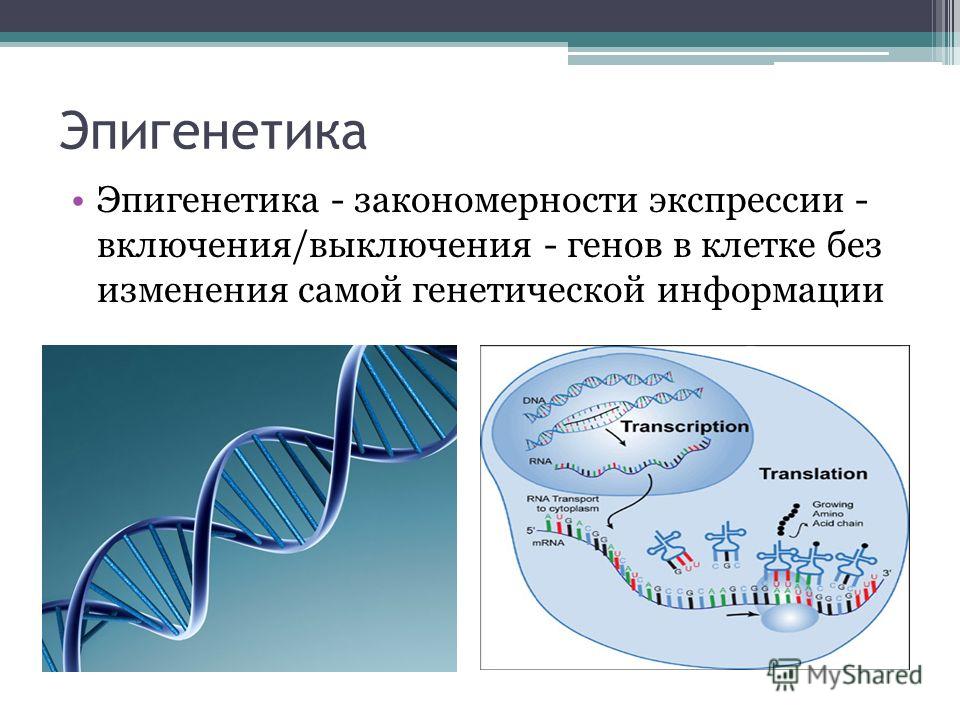
Данное обследование может быть применено в диагностике поражения шейки матки, где это является наиболее частым и востребованным методом исследования. Оно может позволить:
- достоверно оценивать потенциал дисплазии в отношении развития рака шейки матки и, соответственно, выбирать более консервативную или более агрессивную тактику лечения;
- уточнять заключения цитолога — дополнительному исследованию на онкопротеина р16ink4a подлежат все случаи атипической цитологии (кроме инвазивного рака шейки матки), неопределенные цитологические заключения (атипические клетки плоского эпителия неясного значения-ASCUS) и все изменения железистого эпителия;
- разрешать спорные вопросы при выявлении гинекологом высокоаномальной кольпоскопической картины, не сопровождающейся изменениями в цитологическом мазке;
- во многих случаях обоснованно отказаться от биопсии — у большинства пациенток пожилого возраста с цитологической картиной дисплазии на фоне атрофического цервицита обоснованно избирать выжидательную тактику, отказавшись не только от лечения, но и от биопсий, и увеличить интервалы между диспансерными осмотрами; у небольшого числа р16ink4a-позитивных пациенток этой возрастной группы, напротив, обоснованно избрать более активную тактику;
- определять индивидуальную тактику в отношении пациенток, инфицированных штаммами вируса папилломы человека высокого онкогенного потенциала, если у них нет цитологических и кольпоскопических изменений; у женщин с позитивной ПЦР по HPV выделить группу пациенток, у которых уже инициирован канцерогенез в эпителии шейке матки, и направить на них необходимое лечебное воздействие; соответственно, в более многочисленной группе р16ink4a-негативных женщин необходимо продолжить мониторинг;
- усовершенствовать мониторинг пациенток, прошедших органосохраняющее лечение по поводу «тяжелой дисплазии» или «рака in situ», добавив определение онкопротеина р16ink4a к рутинному цитологическому исследованию, это позволит у пациенток наибольшего риска повысить чувствительность цитологического исследования и тестировать персистирующую папиллома-вирусную инфекцию до появления морфологических изменений.


Для чего используется исследование?
- Обследование женщин независимо от наличия или отсутствия патологии шейки матки;
- обследование женщин и мужчин при подозрении на рак шейки матки, рак толстой кишки, рак молочной железы, рак мочевого пузыря, рак предстательной железы;
- наличие патологии шейки матки;
- контроль эффективности лечения патологии шейки матки
Когда назначается исследование?
- При наличии клинико-анамнестических признаков (эрозия, лабораторно подтвержденное инфицирование ВПЧ), позволяющих заподозрить онкологический процесс в шеечной части матки, мочевого пузыря, прямой кишке;
- все случаи атипической цитологии (кроме инвазивного рака шейки матки), неопределенные цитологические заключения (атипические клетки плоского эпителия неясного значения) и все изменения железистого эпителия;
- при выявлении высокоаномальной кольпоскопической картины, не сопровождающейся изменениями в цитологическом мазке;
- при наблюдении за пациентками, прошедшими органосохраняющее лечение по поводу «тяжелой дисплазии» или «рака in situ» в целях повышения чувствительности цитологического исследования.

Что означают результаты?
Референсные значения: отсутствие экспрессии белка р16 в клетках эпителия (негативная реакция).
Заключение включает в себя два подраздела: цитологическое описание и иммуноцитохимическое исследование на р16ink4a.
Цитологическое заключение интерпретируется в зависимости от выявления онкомаркера р16ink4a. В особенности это относится к дисплазиям шейки матки.
Белок р16(INK4a) осуществляет контроль разобщения комплекса E2F-Rb, препятствуя пролиферации клетки. В норме по механизму обратной связи синтез р16 сдерживается, таким образом, концентрация данного белка в нормальной клетке чрезвычайно мала, что проявляется негативной иммуноцитохимической реакцией.
Достоверно показано, что экспрессия р16 связана с низкой, умеренной и тяжелой дисплазией, причем экспрессия р16 практически не встречается в плоском эпителии без признаков дисплазии (0-2% случаев).
Экспрессия белка р16 нарастает с увеличением степени дисплазии и выявляется:
- в 16% случаев при CIN I – плоскоклеточном интраэпителиальном поражении низкой степени тяжести – воспалительные изменения, характерные для инфекции ВПЧ, слабая дисплазия эпителия;
- в 64% при CIN II — плоскоклеточное интраэпителиальное поражение высокой степени тяжести – умеренная дисплазия эпителия;
- в 90% случаев при CIN III — тяжелая дисплазия эпителия и внутриэпителиальный рак шейки матки;
- в 100% при плоскоклеточном раке шейки матки.

Кто назначает исследование?
Гинеколог, онколог.
Также рекомендуется
[12-134] Определение предиктора изменений эндометрия PTEN, Ki67
[09-106] Human Papillomavirus высокого канцерогенного риска (16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59 типы), ДНК генотипирование [реал-тайм ПЦР]
Литература
- Аляутдина О.С., Синицына О.В. ЗНАЧЕНИЕ ТЕСТА НА ОНКОБЕЛОК Р16INK4A В АЛГОРИТМЕ ДИАГНОСТИКИ РАКА ШЕЙКИ МАТКИ // Международный журнал прикладных и фундаментальных исследований. – 2016. – № 11-1. – С. 58-60.
- Ашрафян Л.А., Харченко Н.В., Огрызкова В.Л., Антонова И.Б. Принципы лечения премикроинвазивного рака шейки матки.// Практическая онкология. — 2002. — Том 3. — С. 173-178;17.
- Бохман Я.В. Руководство по онкогинекологии./Я.В. Бохман. – С.-Пб. «Фолиант». 2002.
- Н.В. Данилова, Ю.Ю. Андреева, с соавт. Маркеры апоптоза и белки-регуляторы клеточного цикла при фоновых и предраковых изменениях железистого эпителия шейки матки. Онкология. Журнал им. П.А. Герцена, 1, 2013.
- Протасова А.Э. Клиническая оценка и фармако-экономический анализ диагностики и лечения основных злокачественных опухолей женских гениталий в амбулаторных условиях.Автореф. дис. д-ра мед. наук. СПб., 2011.
- Сагайдак В.Н., Комарова Л.Е. Рак шейки матки и цитологический скрининг. – М. 1994.
- Международный журнал прикладных и фундаментальных исследований. – 2016. – № 11 (часть 1) – С. 58-60.
- Сергеева Н.С., Маршутина Н.В. Новые серологические и уринологическиеопухолевые маркеры в уточняющей диагностике и мониторинге онкологических больных // Материалы VII Российской онкологической конференции. — М., 2003.
- Хансон К.П., Имянитов Е.Н. Современные представления о канцерогенезе рака шейки матки.// Практическая онкология — 2002. — Том 3. — С. 145-155;18.
- Экспрессия р53 в поверхностных уротелиальных карциномах мочевого пузыря – независимый фактор прогноза. М.В. Ковылина, Е. А. Прилепская, О.А. Цыбуля, Н.В. Тупикина, И.В. Рева.ОНКОУРОЛОГИЯ 2’2016. ТОМ 12.
- Bibbo M., Klump W.J., DeCecco J., Kovatish A.J. Procedure for immunocytochemical detection of p16INK4A antigen in thin-layer, liquid-based specimens.// ActaCytol. — 2002. — V.46 — P. 25-29;15.
- Jenison S.A., Yu X.P., Valentine J.M., Galloway D.A. Characterization of human antibody-reactive epitopes encoded by human papillomavirus types 16 and 18. // J Virol. 1991 — V.65(3) — P. 1208-1218;1.
- Klaes R., Froedrich T., Spitkovsky D. et al. Overexpression of p16(INK4A) as a specific marker for dysplastic and neoplastic epithelial cells of the cervix uteri.// Int J Cancer. 2001 — V.92(2) — P.276-84;4.
- Meyer J.L., Hanlon D.W., Andersen B.T. et al. Evaluation of p16INK4a expression in ThinPrep cervical specimens with the CINtec p16INK4a assay: correlation with biopsy follow-up results.// Cancer. — 2007. — V. 111(2). — P. 83-92;20.
- Nieh S., Chen S. F., Chu T.Y. et al. Expression of p16 INK4A in Papanicolaou smears containing atypical squamous cells of undetermined significance from the uterine cervix. // Gynecol Oncol. — 2003. — V.91(1) — P.201-8;5.
 Онкология. Журнал им. П.А. Герцена, 1, 2013.
Онкология. Журнал им. П.А. Герцена, 1, 2013. А. Прилепская, О.А. Цыбуля, Н.В. Тупикина, И.В. Рева.ОНКОУРОЛОГИЯ 2’2016. ТОМ 12.
А. Прилепская, О.А. Цыбуля, Н.В. Тупикина, И.В. Рева.ОНКОУРОЛОГИЯ 2’2016. ТОМ 12. F., Chu T.Y. et al. Expression of p16 INK4A in Papanicolaou smears containing atypical squamous cells of undetermined significance from the uterine cervix. // Gynecol Oncol. — 2003. — V.91(1) — P.201-8;5.
F., Chu T.Y. et al. Expression of p16 INK4A in Papanicolaou smears containing atypical squamous cells of undetermined significance from the uterine cervix. // Gynecol Oncol. — 2003. — V.91(1) — P.201-8;5.СРАВНЕНИЕ МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКОЙ СТРУКТУРЫ ОПУХОЛЕВЫХ КЛЕТОК ДО ЛЕЧЕНИЯ И ПОСЛЕ КОНСТАТАЦИИ РЕЦИДИВА МНОЖЕСТВЕННОЙ МИЕЛОМЫ (КРАТКИЙ ОБЗОР И ОПИСАНИЕ КЛИНИЧЕСКОГО СЛУЧАЯ) | Сергеева
Введение
Множественная миелома (ММ) относится к злокачественным заболеваниям системы крови с клональной пролиферацией плазматических клеток. Патогенез ММ представляет собой многоступенчатый процесс трансформации генетического материала клетки, который изучен лишь частично [1]. Молекулярно-цитогенетическое исследование клеток костного мозга у больных ММ является рутинным методом обследования, по результатам которого проводится стратификация больных на группы риска и стадирование ММ. При цитогенетическом исследовании определяются: транслокации с вовлечением локуса генов IGH 14-й хромосомы, наличие трисомий по целому ряду нечетных хромосом, делеция короткого плеча 17-й хромосомы, моносомия или делеция длинного плеча 13-й хромосомы (del13q) и амплификация локуса 1q21 (amp1q21) [2]. Наряду с этим для тестирования точечных мутаций в генах, ассоциированных с развитием ММ, широко используется метод секвенирования по Сэнгеру, позволяющий получать надежные результаты при анализе последовательности ДНК на предмет единичных нуклеотидных замен, микроделеций и микроинсерций коротких участков длиной от одного нуклеотида до нескольких десятков оснований. Однако этот метод пригоден только для анализа мутаций, носящих клональный характер или присутствующих в субклонах, составляющих не менее 10 % от общей опухолевой массы. Исследования с использованием современных методов секвенирования показали высокую гетерогенность различных генетических нарушений при ММ [3, 4].
Наряду с этим для тестирования точечных мутаций в генах, ассоциированных с развитием ММ, широко используется метод секвенирования по Сэнгеру, позволяющий получать надежные результаты при анализе последовательности ДНК на предмет единичных нуклеотидных замен, микроделеций и микроинсерций коротких участков длиной от одного нуклеотида до нескольких десятков оснований. Однако этот метод пригоден только для анализа мутаций, носящих клональный характер или присутствующих в субклонах, составляющих не менее 10 % от общей опухолевой массы. Исследования с использованием современных методов секвенирования показали высокую гетерогенность различных генетических нарушений при ММ [3, 4].
По данным, полученным на большой выборке больных с использованием секвенирования полного эк- зома (whole exome sequencing, WES), наиболее часто встречающимися нарушениями у больных ММ являются точечные мутации в генах сигнального MAPK- пути (N-RAS, K-RAS, BRAF) и онкосупрессора ТР53, которые суммарно встречаются примерно у половины больных [5].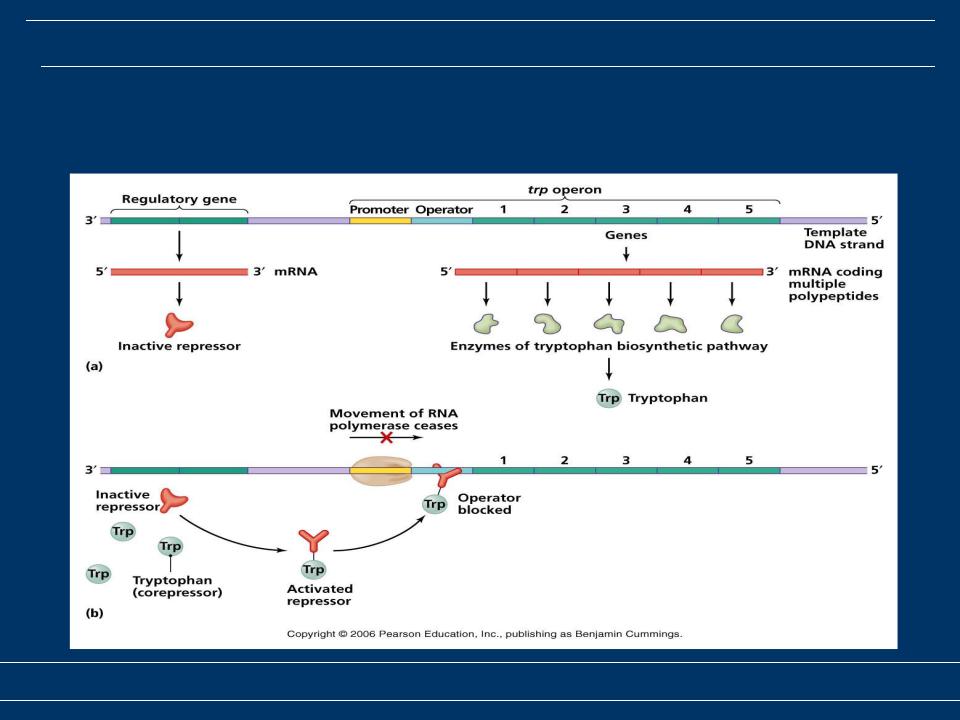
В патогенезе ММ нарушается регуляция транскрипции целого ряда генов, таких как C-MYC, MMSET (NSD2), CCND1, CCND2, CKS1B, Notch3, IL-6, MDM2. В результате появляется понятие «профиля» экспрессии генов, различные варианты которого могут иметь прогностическое значение, определяя характер течения заболевания и его резистентность к терапии [6].
Гены семейства RAS. Гены семейства RAS — наиболее часто мутирующие гены при ММ. В исследовании W. Chng и соавт. [7] частота встречаемости мутаций генов семейства RAS составила 23 % (102/561), из них у 74 (17 %) были выявлены мутации в гене N-RAS, у 28 (6 %) — в гене К-RAS. Большинство мутаций было детектировано в кодоне 61-го гена N-RAS (64 из 74). По данным литературы [8], активирующие миссенсмутации в генах N-RAS и К-RAS изменяют свойства соответствующих белков и превращают протоонкогены в онкогены. Функциональная нагрузка таких мутаций заключается в том, что белки RAS теряют ГТФазную активность, вследствие чего нарушается их нормальная регуляция цитокинами в передаче митоген-активирующего сигнала к ядру, что приводит к повышенной пролиферации клеток [8]. Стадия, предшествующая ММ, моноклональная гаммапатия неясного генеза (МГНГ), на молекулярном уровне отличается от ММ отсутствием мутаций в генах семейства RAS [9]. Имеющиеся в литературе сведения о роли активирующих мутаций в генах K-RAS и N-RAS в патогенезе ММ весьма противоречивы. По одним данным, они считаются фактором неблагоприятного прогноза, ассоциируются с прогрессией заболевания и меньшей общей выживаемостью, по другим — могут влиять только на резистентность к терапии определенными препаратами (например, бортезомибом) [7, 10, 11].
Стадия, предшествующая ММ, моноклональная гаммапатия неясного генеза (МГНГ), на молекулярном уровне отличается от ММ отсутствием мутаций в генах семейства RAS [9]. Имеющиеся в литературе сведения о роли активирующих мутаций в генах K-RAS и N-RAS в патогенезе ММ весьма противоречивы. По одним данным, они считаются фактором неблагоприятного прогноза, ассоциируются с прогрессией заболевания и меньшей общей выживаемостью, по другим — могут влиять только на резистентность к терапии определенными препаратами (например, бортезомибом) [7, 10, 11].
BRAF. Киназа BRAF передает сигнал от RAS белков к нижестоящим эффекторным белкам сигнального пути МАРК. Частота мутаций в гене BRjAF при ММ варьирует от 4 до 14,9 % [12]. Чаще всего встречается активирующая мутация в 600-м кодоне с заменой валина на глютаминовую кислоту (V600E). Показано [13], что эта мутация ассоциируется с клинически более агрессивным течением ММ, более низкими показателями общей выживаемости, а также с высокой частотой (>50 %) выявления экстрамедуллярных очагов.
TP53. Опухолевый супрессор p53 является транскрипционным фактором. Этот белок связывается с промоторами различных генов и может как активировать, так и ингибировать их транскрипцию. Белок p53 вовлечен в репликацию и репарацию ДНК. Отсутствие гена ТР53 ведет к развитию синдрома Ли — Фраумени, проявляющегося предрасположенностью к отдельным новообразованиям и возникновению первично-множественных опухолей [14]. Известен спектр нарушений, ведущих к дисфункции белка p53. Сюда относятся мутации гена TP53, метилирование его регуляторных областей, нарушение транспорта белка p53 в ядро, амплификация ингибитора p53 белка mdm2, связывание с вирусным белком E6, которое ведет к деградации p53 [14]. Кроме того, при делеции короткого плеча хромосомы 17 (p17del) одна копия гена утрачивается полностью. Инактивация белка p53 считается универсальным изменением в опухолевой клетке и встречается в 50—60 % новообразований. В основном генные нарушения встречаются в гетерозиготном состоянии, представлены миссенс-мутациями, приводящими к аминокислотным заменам, и затрагивают чаще всего ДНК-связывающий домен. В результате подобных мутаций происходит изменение вторичной структуры белка и появление новых свойств, таких как способность активировать экспрессию некоторых генов, к которым относятся с-MYC, CCND1, MRD1. Усиление экспрессии MRD1 связано с появлением множественной лекарственной устойчивости у клеток опухоли [15].
В результате подобных мутаций происходит изменение вторичной структуры белка и появление новых свойств, таких как способность активировать экспрессию некоторых генов, к которым относятся с-MYC, CCND1, MRD1. Усиление экспрессии MRD1 связано с появлением множественной лекарственной устойчивости у клеток опухоли [15].
с-MYC. Протоонкоген с-MYC задействован во многих путях передачи сигналов от рецепторных комплексов на поверхности мембраны клетки. Его экспрессия находится под контролем целого набора транскрипционных регуляторов [16]. Белок c-myc вовлечен в большое число внутриклеточных взаимодействий, его повышенная экспрессия ассоциирована с начальными стадиями неопластической трансформации клеток и усилением их пролиферации [17]. В спектр функций c-myc входит регуляция внутриклеточных систем энергообеспечения — гликолиза, метаболизма глютамина и биогенеза митохондрий, которые приводят к аккумуляции энергии для репликации ДНК и деления клетки [18]. Повышенная экспрессия этого белка ассоциируется с геномной нестабильностью, вызываемой стимуляцией митохондрий, которая приводит к наработке активных форм кислорода [19], и в частности может быть связана с появлением транслокаций [20].
MMSET (NSD2). Белок MMSET (multiple myeloma SET domain) является гистонметилтрансферазой и принимает участие в реорганизации хроматина. Его повышенная экспрессия наблюдается на начальных стадиях патогенеза ММ. Транслокация с участием гена MMSET t(4;14) является фактором неблагоприятного прогноза. Искусственная репрессия белка MMSET приводит к пониженному темпу пролиферации, индукции апоптоза и клеточной агдезии [21].
CCND1 и CCND2. Высококонсервативные белки из семейства D-циклинов, CCND1 и CCND2, представляют собой регуляторные субъединицы киназ, необходимые для перехода клетки из G1 в S-фазу клеточного цикла. Эти белки взаимодействуют с опухолевым супрессором Rb, и их экспрессия положительно регулируется Rb. Результатом ранних событий в патогенезе множественной миеломы, таких как IGH-транслокации, появление гипердиплоидности и трисомии, является гиперэкспрессия этих циклинов [22].
CKS1B. Увеличение экспрессии гена субъединицы 1В циклинкиназы является фактором неблагоприятного прогноза при ММ. Функция CKS1B заключается в активации циклина через его фосфорилирование [23]. Ген расположен на длинном плече 1-й хромосомы, его амплификация регулярно встречается среди генетических нарушений множественной миеломы [24].
Функция CKS1B заключается в активации циклина через его фосфорилирование [23]. Ген расположен на длинном плече 1-й хромосомы, его амплификация регулярно встречается среди генетических нарушений множественной миеломы [24].
Notch3. Белки семейства Notch вовлечены в регуляцию гемопоэза. Отсутствие белков этого семейства ведет к нарушению поздних стадий дифференцировки В-клеток [25]. Нарушение регуляторных механизмов работы рецепторов или их лигандов, входящих в сигнальный путь Notch, исследовано в различных типах злокачественных солидных и гематологических опухолей, в том числе и при ММ. Ингибирование Notch индуцирует апоптоз в клетках опухоли, уменьшает устойчивость к лекарственным препаратам, изменяет характеристики миграции/рециркуляции плазматических клеток и их обновление [26]. Notch может участвовать в прогрессии ММ, увеличивая уровень интерлейкина-6 (IL-6), одного из ключевых факторов, стимулирующих клеточную пролиферацию [27].
Интерлейкин-6. Представляет собой цитокин плей- отропного действия. Он индуцирует дифференцировку и рост разных типов клеток, в том числе дифференцировку нормальных B-клеток в плазматические клетки, производящие антитела. Пролиферативная активность клеток множественной миеломы зависит от IL-6, осуществляющего ее аутокринную регуляцию [28]. Экспрессия IL-6 ассоциирована с агрессивным течением заболевания, высоким пролиферативным индексом и устойчивостью к лекарственным препаратам, запускающим апоптоз [29]. Повышение активности IL-6 связано с остеодеструкцией — повышенной активностью и пролиферацией остеокластов и понижением количества остебластов при ММ [30]. Высокий уровень экспрессии IL-6 является фактором плохого прогноза при ММ.
Представляет собой цитокин плей- отропного действия. Он индуцирует дифференцировку и рост разных типов клеток, в том числе дифференцировку нормальных B-клеток в плазматические клетки, производящие антитела. Пролиферативная активность клеток множественной миеломы зависит от IL-6, осуществляющего ее аутокринную регуляцию [28]. Экспрессия IL-6 ассоциирована с агрессивным течением заболевания, высоким пролиферативным индексом и устойчивостью к лекарственным препаратам, запускающим апоптоз [29]. Повышение активности IL-6 связано с остеодеструкцией — повышенной активностью и пролиферацией остеокластов и понижением количества остебластов при ММ [30]. Высокий уровень экспрессии IL-6 является фактором плохого прогноза при ММ.
MDM2 (murine double minute 2) — белок ингибитор TP53. Изначально ген этого белка был идентифицирован в экстрахромосомной ДНК, образующейся в раковых клетках. Его взаимодействие с TP53 ведет к тому, что комплекс MDM2/TP53 направляется из ядра в цитоплазму, где подвергается убиквитинированию. В условиях стресса сайты связывания MDM2 и TP53 фосфорилируются, и комплекс, в котором TP53 не активен, не образуется. В результате белок TP53 переходит из латентной формы в активированную форму, запускает процессы остановки клеточного цикла и апоптоза. Гиперэкспрессия MDM2 — типичный механизм разоружения раковой клетки, лишающий ее защитных функций опухолевого супрессора TP53 [31].
В условиях стресса сайты связывания MDM2 и TP53 фосфорилируются, и комплекс, в котором TP53 не активен, не образуется. В результате белок TP53 переходит из латентной формы в активированную форму, запускает процессы остановки клеточного цикла и апоптоза. Гиперэкспрессия MDM2 — типичный механизм разоружения раковой клетки, лишающий ее защитных функций опухолевого супрессора TP53 [31].
Цель исследования — проанализировать молекулярно-генетический статус опухоли у больного ММ с коротким периодом ремиссии в дебюте и рецидиве заболевания и сопоставить с клиническим течением заболевания.
Материалы и методы
Пункцию костного мозга для молекулярно-генетического скрининга (fluorescence in situ hybridization (FISH), измерение экспрессии и мутационный статус генов) проводили дважды — в дебюте и рецидиве ММ.
Для оценки значимости изменения величин экспрессии генов регуляторных белков у больного ММ в дебюте и рецидиве заболевания сравнивали их с соответствующими показателями для клеток CD138+, полученных от 10 доноров (5 женщин, 5 мужчин в возрасте от 25 до 41 года).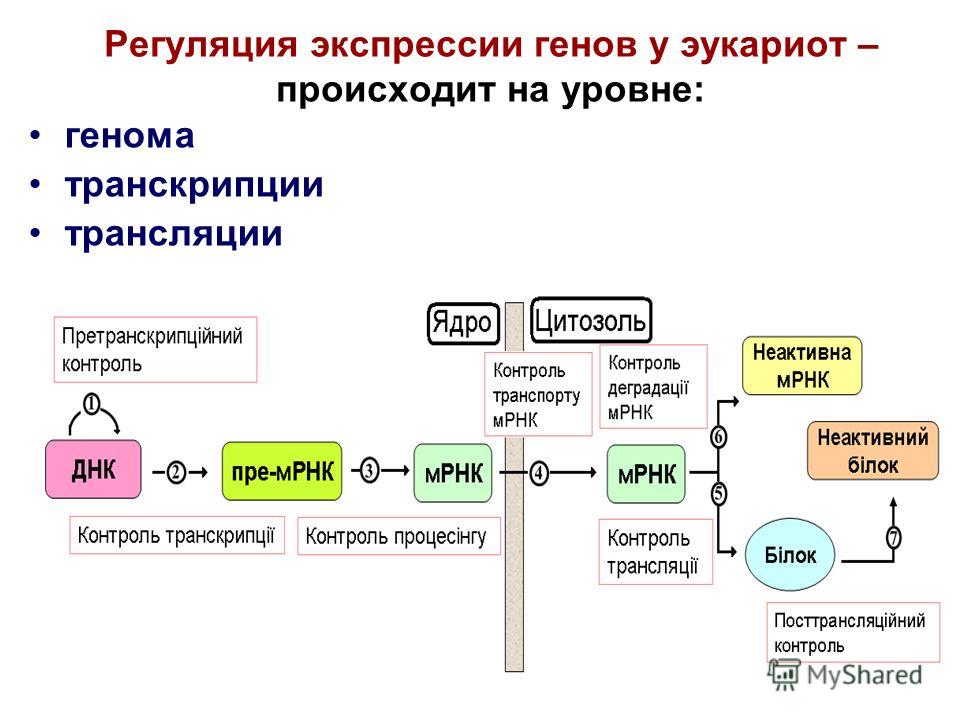 Если значение экспрессии гена у больного вписывалось в диапазон значений, характерных для доноров, считали, что оно находится в пределах нормы.
Если значение экспрессии гена у больного вписывалось в диапазон значений, характерных для доноров, считали, что оно находится в пределах нормы.
Выделение мононуклеаров из костного мозга и CD138-клеток. Используя градиент плотности фиколла (1,077 г/ см3), из костномозговой взвеси выделяли фракцию мононуклеаров. В дальнейшем проводилась высокоактивная магнитная сепарация согласно протоколу Miltenyi Biotec (Miltenyi Biotec GmbH, Germany, http://www.miltenyibiotec.com) с использованием магнитного сепаратора OctoMacs, антител anti-CD138, конъюгированных с 50-нм частицами оксида железа и антител anti-CD138, конъюгированных с фикоэритрином. Чистота выделения мононуклеаров, обогащенных С0138+-клетками, в дебюте (установка диагноза 29.05.2013) и прогрессии (констатация рецидива 01.08.2014) составила 80 и 70 % соответственно. В дальнейшем полученные мононуклеары и С0138+-клетки использовали для проведения FISH-исследования, измерения уровня экспрессии генов и определения мутационного статуса генов.
Флуоресцентная гибридизация in oitu (FISH). Молекулярно-цитогенетическое исследование клеток костного мозга выполняли в лаборатории кариологии ФГБУ «НМИЦ гематологии» Минздрава России. В дебюте заболевания FISH-исследование проводилось на моно- нуклеарах костного мозга для выявления первичных (t(14q32)/IGH, множественные трисомии) и вторичных (del 17p13/TP53, del13q14/-13, del1p32, amp1q21 и t(8q24)/ cMYC) хромосомных аномалий. В прогрессии заболевания на сепарированных С0138+-клетках проводился повторный FISH-анализ с ДНК-зондами для выявления del17p13/TP53, del13q14/-13, del1p32, amp1q21, t(8q24)/cMYC и множественных трисомий. В работе использовали различные центромерные и локус-спе- цифичные ДНК-зонды: XL IGHplus, XL P53, XL cMYC BA, XL 1p32/1q21, XL 5p15/9q22/15q22 Hyperdiploidy Amplification Probe (MetaSystems, Germany) и D13S25 (Cytocell, UK). Исследование проводили согласно протоколам производителей. Для каждого зонда анализировали по 200 интерфазных ядер с четкими сигналами. Результаты FISH-анализа описывали в соответствии с международной номенклатурой (International System for Cytogenetic Nomenclature, ISCN, 2013) [32].
Анализ экспрессии генов. Тотальную фракцию РНК выделяли из плазматических клеток костного мозга с поверхностным маркером CD138+ при помощи лизиса в гуанидин-изотиоцианатном буфере с последующей очисткой стандартным фенол-хлороформным методом. Полученную РНК хранили при температуре —70 °С в EtOH. Для анализа экспрессии генов использовали метод секвенирования РНК нового поколения (RNA-seq). Анализ проводили на приборе Illumina HiSeq (Illumina, США) с использованием наборов для подготовки библиотеки транскриптома тотальной РНК (Whole-transcriptome analysis with total RNA sequencing). Выравнивание полученных после секвенирования последовательностей на референсный геном и сборка транскриптома проводилось с помощью программного обеспечения STAR [33]. Пересчет уровня экспрессии генов был выполнен в программе DESeq2. Числовые значения экспрессии (normalized gene counts, ngc) представляют собой абсолютные значения, полученные при сборке и выравнивании транскрипта гена на последовательность референс- ного гена с последующим нормированием на глубину секвенирования [34]. Для определения значимости различий между экспрессией в опухолевых клетках в дебюте и рецидиве заболевания полученные значения приведены с меньшим и большим диапазоном значений экспрессии в плазматических клетках 10 доноров (табл. 1).
Таблица 1. Изменения уровня экспрессии генов в дебюте ММ и при констатации рецидива у больного в сравнении со значениями экспрессии этих генов у доноров
Table 1. Gene expression fold changes are at the time of diagnosis and progression in comparison with a range of gene expression values for a group of donors
Гены Genes | Доноры (n = 10) Donors (n = 10) | Больной Patient | ||
|---|---|---|---|---|
минимальное значение, ngc min value, ngc | максимальное значение, ngc max value, ngc | дебют заболевания, ngc disease debut, ngc | рецидив заболевания, ngc disease relapse, ngc | |
с-myc | 188 | 493 | 1374 | 2074 |
CCND1 | 5 | 981 | 117 | 12 |
CCND2 | 332 | 4592 | 4383 | 613 |
MMSET(NSD2) | 611 | 1608 | 338 | 1195 |
Notch3 | 945 | 2276 | 1763 | 5805 |
CKS1B | 4 | 40 | 17 | 29 |
IL6 | 2 | 41 | 33 | 953 |
TP53 | 166 | 479 | 237 | 210 |
MDM2 | 384 | 910 | 1821 | 4458 |
RAF1 | 706 | 1395 | 791 | 1710 |
STAT4 | 22 | 143 | 46 | 145 |
mTOR | 456 | 723 | 625 | 1585 |
Клональные IGH/L Сlonal IGH/L | ||||
IGHG2 | 35146 | 280485 | 628255 | 67532 |
IGHM | 46047 | 279812 | 2118 | 298 |
IGHV3-33 | 811 | 11080 | 13954 | 1246 |
IGLC2 | 25465 | 126208 | 242255 | 22381 |
IGLV1-44 | 1460 | 6682 | 21176 | 1491 |
Неклональные IGH Non-clonal IGH | ||||
IGHV3-23 | 8970 | 48891 | 940 | 14 |
IGHV2-5 | 3204 | 15381 | 1613 | 36 |
Примечание. ngc — normalized gene counts, единица измерения экспрессии гена, выраженная в количестве прочтений гена, нормированного на общее число прочтений в образце.
Note. ngc — normalized gene counts (units of given values).
Анализ соматических мутаций. Соматические мутации в геномной ДНК плазматических клеток CD138+ костного мозга анализировали методом секвенирования по Сэнгеру. В данной работе исследовались кодирующие области генов N-RAS, K-RAS, TP53, и экзон 15 гена BRAF. Геномную ДНК выделяли методом фенол-хлороформной экстракции. Клетки лизировали в 1 мл STE (Sodium Chloride-Tris-EDTA) буфера, содержащего 1 % додецлсульфат натрия (SDS) и протеиназу К (200 мкг/мл), в течение ночи при 37 °С или двух часов при 60 °С с последующей фенольной экстракцией.
Для постановки полимеразной цепной реакции (ПЦР) использовали смесь PCR Master Mix (2X) Thermo Scientific™, 0,01—0,02 мкг геномной ДНК и 10 пкмоль каждого из праймеров в усредненных условиях (94 °С — 1 мин., 60—62 °С — 1 мин., 72 °С — 1—2 мин., 30 циклов). Все праймерные системы и зонды являются оригинальными и разработаны в ходе проведения данного исследования. Для гена BRAF праймеры были синтезированы для 15-го экзона: прямой праймер BRAF15D: gatctcttacctaaactcttca; обратный праймер BRAF15R: ccttcaatgactttctagtaact, для гена NRAS и KRAS праймеры были подобраны таким образом, чтобы ам- плифицировать 2, 3 и 4-й экзоны обоих генов: RAS1D: atgtggctcgccaattaacc и RAS1R tgggtaaagatgatccgacaa; RAS1: cccttaccctccacaccccc; RAS2 ctcatttccccataaagattcag; NRAS3x: ttcaagcagtctgccctcct; NRAS4: aactgatgcaaactcttgcaca; KRASD1: gatacacgtctgcagtcaac; KRASR1: tcctgcaccagtaatatgcat; KRASD2: ccagactgtgtttctcccttg; KRASR2: ttactccactgctctaatccc; KRAS3: gacaaaagttgtggacaggtt; KRAS4:ggacactggattaagaagcaa. Все кодирующие экзоны TP53, кроме 11-го, были включены в анализ и амплифицировались с использованием следующих праймеров: TP53D1: gccgagctgtctcagacact; TP53R1: gaggaatcccaaagttccaaacaa; TP53D2: acg ccaactctctctagctc; TP53R2: ggccactgacaaccaccctta; TP53D3: ggcctcccctgcttgccaca; TP53R3: caaccacccttgtcctttct; TP53D4: ggcttctcctccacctacct; TP53R4: gcaggctaggctaagctatga; TP53D5: catgttgcttttgtaccgtca; TP53R5: cagctgcctttgaccatgaa.
Продукты ПЦР разделяли при помощи электрофореза в 6 % полиакриламидном геле (ПААГ) и визуализировали в УФ-свете после окрашивания бромистым этидием. Для секвенирования ПЦР-продукты реакции очищали на колонках Wizard® SV Gel and PCR Clean-Up System Promega.
Праймеры синтезировали в ООО «Синтол». Секве- нирование проводили с помощью набора реактивов ABI PRISM®BigDyeTM Terminator v.3.1 с последующим анализом продуктов реакции на автоматическом секвенаторе ДНК ABI PRISM 3500 в ЦКП «Геном». Полученные с секвенатора электрофореграммы анализировали в программе Vector NTI.
Результаты
Клиническое течение заболевания
Больной И. Ю. И., 1957 года рождения, наблюдался в ФГБУ «НМИЦ гематологии» Минздрава России с мая 2013 г. с диагнозом ММ IIA ст. по Durie-Salmon, I ст. по ISS, I ст. по R-ISS. Диагноз был установлен в соответствии с критериями, разработанными Международной рабочей группой по изучению ММ (The International Myeloma Working Group — IMWG) [35]. На момент диагностики заболевания был выполнен первый молекулярно-генетический скрининг (FISH, определение экспрессии и мутационный статус генов). При проведении индукции ремиссии было выполнено пять курсов по программе PAD (бортезомиб + адриабластин + дексаметазон). После второго курса была достигнута частичная ремиссия (ЧР), а после пяти — очень хорошая частичная ремиссия (ОХЧР).
На фоне ОХЧР были выполнены мобилизация и сбор аутологичных стволовых клеток крови по схеме 4 г/м2 и гранулоцитарный колониестимулирующий фактор 5 мкг/кг. За две процедуры лейкафереза было заготовлено 13,0 X 106/кг CD34+-клеток. В перерыве между сбором аутологичных стволовых клеток крови и выполнением трансплантации аутологичных гемопоэтических стволовых клеток (ауто-ТГСК) с целью сдерживания достигнутого противоопухолевого ответа проведено четыре бортезомиб-содержащих курса (VCD: борте- зомиб + дексаметазон). По результатам обследования непосредственно перед ауто-ТГСК была констатирована полная строгая ремиссия заболевания (результаты иммунохимического исследования, данные костного мозга). Однако сохранялись остеодеструктивные очаги, замещенные содержимым жировой плотности. При обследовании на +100-й день после ауто-ТГСК была констатирована прогрессия заболевания (В-симптомы, по данным иммунохимического исследования сыворотки крови и мочи помимо исходного парапротеина G-лямбда, впервые за весь период наблюдения была выявлена секреция парапротеина М-лямбда 5,3 г/л). Реин- дукционные бортезомиб-содержащие курсы оказались неэ фф ективными. Больной был переведен на вторую линию терапию иммуномодулирующими препаратами, на фоне которой удалось достичь лишь частичного противоопухолевого ответа. Спустя четыре месяца от момента констатации рецидива заболевания больной умер от тяжелых инфекционных осложнений.
Молекулярно-цитогенетическое исследование (FISH). При молекулярно-цитогенетическом исследовании (FISH) в дебюте заболевания был выявлен лишь гипердиплоидный тип ММ (трисомии 5, 9, 15) в 50 % ядер. При повторном цитогенетическом исследовании на фоне прогрессии заболевания сохранялся гипердиплоидный тип ММ, однако появилась amp1q21 в 170 интерфазных ядрах из 200 исследованных, то есть в 85 % ядер. На рисунке 1 представлены результаты FISH-исследования мо- нонуклеаров костного мозга в дебюте заболевания, а на рисунке 2 — результаты FISH CD138+-клеток костного мозга в рецидиве ММ.
Рисунок 1. Результаты FISH-исследования мононуклеаров костного мозга в дебюте заболевания: А — гипердиплоидия (трисомии 5, 9, 15): три зеленых сигнала от локуса 5р 1 2, три голубых сигнала от локуса 9q22/NR4A3 и три красных сигнала от локуса 15q22/SMAD6; Б — амплификация локуса 1q21 не выявлена: два зеленых сигнала от локуса 1p32/CDKN2C и два красных сигнала от локуса 1q21/CKS1B
Figure 1. Results of FISH-analysis mononuclear cells bone marrow at diagnosis: А — hyperdiploidy (trisomies 5, 9, 15): three green signals of 5р12 locus, three blue signals of 9q22/NR4A3 locus и three red signals of 15q22/SMAD6 locus; Б — Amplification of locus 1q21 was not found: two green signals of 1p32/CDKN2C locus and two red signals of 1q21/CKS1B locus
Рисунок 2. Результаты FISH-исследования CD138+-клеток костного мозга в рецидиве заболевания: А — гипердиплоидия (трисомии 5, 9, 15): три зеленых сигнала от локуса 5р 1 2, три голубых сигнала от локуса 9q22/NR4A3 и три красных сигнала от локуса 15q22/SMAD6; Б — амплификация локуса 1q21: два зеленых сигнала от локуса 1p32/CDKN2C и четыре красных сигнала от локусс 1q21/CKS1B (два из них дополнительные)
Figure 2. Results of FISH-analysis CD138-positive cells bone marrow at the time of relapse: А — hyperdiploidy (trisomies 5, 9, 15): three green signals of 5р12 locus, three blue signals of 9q22/NR4A3 locus и three red signals of 15q22/SMAD6 locus; Б — Amplification of locus 1q21: two green signals of 1p32/CDKN2C locus and four red signals of 1q21/CKS1B locus (two of them are additional)
Анализ экспрессии генов с-MYC, MMSET, CCND1, CCND2, CKS1B, NOTCh3, IL-6, BCL2, TP53, MDM2. Анализ экспрессии гена с-MYC показал, что в рецидиве заболевания в плазматических клетках костного мозга больного незначительно повысилось количество мРНК с-MYC (в 1,5 раза по сравнению с моментом диагностики заболевания). Экспрессия с-MYC на момент диагностики была в 4,4 раза выше, чем среднее значение, полученное в результате измерений для 10 доноров.
Значительноеувеличениеуровня экспрессии при развитии рецидива заболевания было выявлено для гена Notch3. Экспрессия Notch3 увеличилась в 3,2 раза, причем в дебюте заболевания она не превышала верхние показатели, полученные для доноров, но была выше усредненного значения. Наиболее заметное повышение экспрессии наблюдалось для гена IL-6 в рецидиве заболевания, его экспрессия увеличилась в 29 раз по сравнению с дебютом заболевания. Уже до начала лечения значение экспрессии гена IL-6 было выше в три раза, чем среднее значение у доноров. Высокий уровень экспрессии гена MDM2 — репрессора р53 — был зарегистрирован в дебюте и при рецидиве заболевания, причем при рецидиве его количество возросло в 2,5 раза. Во время диагностики ММ его уровень был выше верхних показателей у доноров в два раза. Повышенное значение экспрессии в рецидиве по сравнению с дебютом заболевания можно также отметить у генов MMSET и CKS1B (в 3,5 и 1,8 раза соответственно). Значительное снижение экспрессии во время прогрессии ММ было детектировано у генов CCND1 и CCND2. Однако ее значения находятся в диапазоне колебаний уровня экспрессии у доноров, и, вероятно, эти изменения не несут функциональной нагрузки. Экспрессия гена TP53 у больного не выходила за рамки значений, полученных для доноров (166 и 479 ngc соответственно) и существенно не изменилась в рецидиве по сравнению с дебютом заболевания.
Отмечено снижение на порядок при рецидиве экспрессии генов клональных иммуноглобулинов, характеризующих опухолевые клетки (IGHV3-33/IGHG2, IGLV1-44/IGLC2′), что соответствует выявленному при иммунохимическом исследовании уменьшению моноклональной секреции парапротеина G-лямбда с 23,8 до 5 ,4 г/л. В то же время экспрессия неклональных иммуноглобулинов (в таблице 1 для примера приведены IGHV3-23 и IGHV2-5) снизилась почти на два порядка, что свидетельствует о наличии у больного с рецидивом заболевания глубокого иммунодефицита. Полученные результаты анализа экспрессии генов представлены на рисунке 3 и в таблице 1.
Рисунок 3. Изменение экспрессии генов в дебюте ММ и при констатации рецидива у больного в сравнении со значениями экспрессии этих генов у доноров. По оси абсцисс расположены гены, по оси ординат — значения их экспрессии, выраженной в количестве прочтений гена, нормированное на общее число прочтений (normalized gene counts, ngc). Уровень экспрессии генов проанализирован в дебюте (синие окружности) и рецидиве (красные окружности) заболевания. Диапазон значений экспрессии данных генов у здоровых людей представлен в виде прямоугольников с перцентилями. Перцентили отображают разброс значений, где в верхнюю и в нижнюю перцентили попадает по 25 % краевых значений. Пересекающая прямоугольник линия — это медиана значений экспрессии для данного гена
Figure 3. Change in gene expression at the diagnosis of MM and at the time of relapse in comparison with the range expression of those genes in the group of donors. Axis X: genes, axis Y: gene expression values in normalized gene counts, ngc. Gene expression level (gene normalized counts) is analyzed at the diagnosis (blue circles) and at the time of relapse (red circles) of the patient with MM. Range of health people gene expression values are represented in the boxes with percentiles. The percentiles show the value below (lower percentiles) or above (upper percentiles) which 25 % of the observations may be found. The line which is crossing the box determines the median value of the expression of the gene
Мутационный статус генов N-RAS, K-RAS, BRAF и ТР53
Секвенирование экзонов 2, 3 и 4 генов N-RAS и K-RAS выявило у больного активирующую соматическую миссенс-мутацию с.182А> C в кодоне 61-го гена N-RAS (p.Q61P). В материале, взятом у больного во время рецидива, диагностированная ранее мутация в той же позиции сохранилась (рис. 4). Соотношение нормального и мутантного пиков на электрофореграмме, близкое к 1, свидетельствует о клональном характере данного нарушения.
Рисунок 4. Активирующая мутация гена N-RAS в кодоне 61. Фрагменты электрофореграмм 3-го экзона гена N-RAS, содержащие миссенс-мутацию (замена аденина на цитозин в 61-м кодоне): А — дебют заболевания; Б — рецидив заболевания
Figure 4. Activating mutation of N-RAS gene in 61st codon. Fragment of the sequence elecfrophoregram of N-RAS gene 3rd exon containing missence mutation (replacement of adenine by cytosine in 61 codon: A — disease debut; Б — disease relapse
При параллельном секвенировании ДНК и РНК гена N-RAS в клетках CD138+ опухоли в дебюте и рецидиве заболевания, электрофореграммы практически не отличались друг от друга (соотношение нормального и мутантного пиков 1:1), что свидетельствует об отсутствии различий в экспрессии мутантного аллеля и аллеля «дикого типа».
Секвенирование экзона 15 гена BRAF, в котором обычно локализуются активирующие соматические мутации, в том числе наиболее распространенная p.V600E, показало отсутствие каких-либо отклонений от референсной последовательности как в первичном, так и в рецидивном материале.
Была проанализирована также первичная структура всех функционально важных областей гена TP53 в материале опухоли как при диагностике ММ, так и при рецидиве заболевания. Соматических мутаций найдено не было. Выявлен только полиморфный вариант (p.P72R, rs1042522 в базе данных NCBI/SNP) в гетерозиготном состоянии, не имеющий функционального значения [36].
Обсуждение
В работе был сопоставлен молекулярно-генетический статус опухолевых клеток у больного ММ в дебюте и при рецидиве заболевания. Исходно противоопухолевый ответ на индукционную терапию бортезомиб-содержащими курсами расценивался как полная иммунохимическая ремиссия, однако при рецидиве заболевания была констатирована резистентность к терапии бортезомибом. Рецидив заболевания сопровождался не только сохранением прежнего клонального парапротеина IgG-λ, но и появлением не выявлявшегося ранее парапротеина IgM-λ. Имму- нохимический рецидив характеризовался появлением симптомокомплекса CRAB: включающего гиперкаль- циемию (Calcium), почечную недостаточность (Renal failure), анемию (Anemia) и остеолитическое поражение костей (Bone lesions). Больной умер в результате инфекционных осложнений через четыре месяца от момента диагностики прогрессии заболевания.
При молекулярно-цитогенетическом исследовании в дебюте и при рецидиве заболевания была выявлена трисомия хромосом 5, 9 и 15. Гипердиплоидия при ММ ассоциируется с благоприятным прогнозом и лучшей выживаемостью больных [37].
На фоне прогрессии ММ у больного возникла новая цитогенетическая аберрация — амплификация локуса хромосомы 1 (amp1q21). Анализ экспрессии гена CKS1B, расположенного в этом локусе, показал ее возрастание при рецидиве примерно в два раза по сравнению с дебютом заболевания, что согласуется с увеличением его копийности. Данная хромосомная аномалия считается фактором плохого прогноза. Отчасти это может быть связано с увеличением числа копий гена CKS1B и усилением его экспрессии, которое может влиять на общую и без- рецидивную выживаемость, а также резистентность к бортезомиб-содержащим курсам [38]. Показано [39] также, что amp1q21 значительно реже встречается при МГНГ, чем при ММ, и может способствовать прогрессии заболевания, однако авторы этой работы пришли к выводу о том, что уровень экспрессии гена CKS1B на этот процесс не влияет, и он обусловлен, скорее всего, генетической нестабильностью длинного плеча хромосомы 1 как таковой.
Анализ транскриптомных данных показал, что при рецидиве в сравнении с дебютом заболевания повышена экспрессия генов с-MYC, Notch3, MDM2 и IL-6. Однако, если экспрессия первых трех генов повысилась всего в 1,5—3 раза, количество транскрипта гена IL-6 возросло почти в 30 раз. Не исключено, что именно резкое усиление экспрессии этого цитоки- на послужило пусковым механизмом прогрессии ММ у больного, поскольку IL-6 стимулирует клеточную пролиферацию, активируя различные сигнальные пути (MAPK, JAK-STAT, PI3K) [1, 40].
При рецидиве внутри каждого из этих путей наблюдалось повышение экспрессии по крайней мере одного гена регуляторного белка (RAF1, STAT4 и mTOR соответственно). В свою очередь, индукция IL-6 может, по крайней мере отчасти, объясняться усилением экспрессии гена Notch3 (в 3,3 раза по сравнению с дебютом заболевания). Notch3 активирует экспрессию IL-6 в клетках костномозговых ниш (клетках стромы) и плазматических клетках костного мозга. Показано, что IL6 является одним из ключевых факторов, влияющих на жизнеспособность и неконтролируемое деление миеломных клеток [30].
Экспрессия гена транскрипционного фактора с-MYC повысилась при рецидиве заболевания незначительно, всего в 1,5 раза, однако, как и в дебюте заболевания, была на существенно более высоком уровне, чем у доноров. Механизм повышенной экспрессии в данном случае остается не ясен, так как при FISH-исследовании ни транслокации с вовлечением локуса гена с-MYC, ни трисомия 8 не были выявлены. Возможно, имеет место эпигенетическая регуляция гена с-MYC. Клетки, конститутивно экспрессирующие высокий уровень с-MYC, обладают пониженной зависимостью от ростовых факторов и большей скоростью пролиферации. Таким образом, повышенный уровень с-MYC также мог быть одним из факторов, спровоцировавших прогрессию ММ.
Экспрессия гена TP53 при рецидиве существенно не изменилась, оставаясь в пределах диапазона значений экспрессии гена TP53 у доноров (табл. 1), и никаких мутаций в нем выявлено не было. При этом экспрессия гена белка mdm2, являющегося ингибитором ТР53, увеличилась в 2,5 раза, что вполне могло повлечь за собой ослабление функции ТР53 при прогрессии заболевания.
Резкое снижение экспрессии всех генов иммуноглобулинов, в том числе и клональных, соответствующих парапротеину G-лямбда, свидетельствовало о наличии глубокого иммунодефицита при прогрессии ММ.
В дебюте заболевания больной не относился к группе высокого риска по клиническим и цитогенетическим данным, однако у него была выявлена клональная гетерозиготная мутация p.Q61P в гене N-RAS, благодаря которой происходит неконтролируемая активация сигнального пути MAPK, ведущая к повышению пролиферативного потенциала опухолевых клеток. После проведения курсов терапии при рецидиве заболевания мутация сохранилась в клональном состоянии. Влияние активирующих мутаций в генах семейства RAS на патогенез ММ исследуется давно и достаточно активно, однако результаты опубликованных работ приводят к противоречивым выводам об их значимости. В исследовании, посвященном изучению дифференциальной экспрессии мутантного и нормального аллелей [11], было показано, что мутантный аллель экспрессируется слабее аллеля «дикого типа». У наблюдавшегося больного нормальная и мутантная копии гена N-RAS в дебюте и рецидиве ММ экспрессировались с одинаковой эффективностью. Оба варианта белка могли играть существенную роль в активации сигнального МАРК-пути при прогрессии заболевания: мутантный белок — за счет потери контроля над ним со стороны цитокинов, нормальный белок — вследствие стимуляции гиперэкспрессией IL-6.
Таким образом, можно сделать вывод о том, что при ММ целесообразно выполнять комплексную молекулярную диагностику, включающую развернутое цитогенетическое исследование, анализ экспрессии и мутационный анализ широкого спектра генов регуляторных белков, так как единичная хромосомная или генная аномалия вряд ли может сама по себе служить определяющим фактором прогноза развития заболевания и его резистентности к применяемой терапии.
1. Prideaux S.M., Conway O’Brien E., Chevassut T.J. The genetic architecture of multiple myeloma. Adv. Hematol. 2014; 864058. DOI: 10.1155/2014/864058
2. Fonseca R., Barlogie B., Bataille R. et al. Genetics and cytogenetics of multiple myeloma: A workshop report. Cancer Res. 2004; 64: 1546–58.
3. Maura F., Degasperi A., Nadeu F. et al. A practical guide for mutational signature analysis in hematological malignancies. Nat Commun. 2019; 10(1): 2969. DOI: 10.1038/s41467-019-11037-8
4. Bolli N., Biancon G., Moarii M. et al. Analysis of the genomic landscape of multiple myeloma highlights novel prognostic markers and disease subgroups. Leukemia. 2018; 32(12): 2604–16. DOI: 10.1038/s41375-018-0037-9
5. Bolli N., Avet-Loiseau H., Wedge D.C. et al. Heterogeneity of genomic evolution and mutational profi les in multiple myeloma. Nat Commun. 2014; 5: 2997. DOI: 10.1038/ncomms3997
6. Szalat R., Avet-Loiseau H., Munshi N.C. Gene Expression Profi les in Myeloma: Ready for the Real World? Clin Cancer Res. 2016; 22(22): 5434–42. DOI: 10.1158/1078-0432.CCR-16-0867
7. Chng W.J., Gonzalez-Paz N., Price-Troska T. et al. Clinical and biological signifi cance of RAS mutations in multiple myeloma. Leukemia. 2008; 22: 2280–4. DOI: 10.1038/leu.2008.142
8. Pylayeva-Gupta Y., Grabocka E., Bar-Sagi D. RAS oncogenes: weaving a tumorigenic web. Nat Rev Cancer. 2011; 11: 761–74.
9. Rasmussen T., Kuehl M., Lodahl M. et al. Possible roles for activating RAS mutations in the MGUS to MM transition and in the intramedullary to extramedullary transition in some plasma cell tumors. Blood. 2005; 105(1): 317–23.
10. Kim S.J., Shin H.T., Lee H.O. et al. Recurrent mutations of MAPK pathway genes in multiple myeloma but not in amyloid light-chain amyloidosis. Oncotarget. 2016; 7(42): 68350–9. DOI: 10.18632/oncotarget.12029
11. Rashid N.U., Sperling A.S., Bolli N. et al. Differential and limited expression of mutant alleles in multiple myeloma. Blood. 2014; 124: 3110–7.
12. Hu Y., Chen E., Wang J. Progress in the identifi cation of gene mutations involved in multiple myeloma. Onco Targets Ther. 2019; 12: 4075–80. DOI: 10.2147/OTT.S205922
13. Andrulis M., Lehners N., Capper D. et al. Targeting the BRAF V600E mutation in multiple myeloma. Cancer Discov. 2013; 3: 862–9. DOI: 10.1158/2159-8290. CD-13-0014
14. Levine A.J. p53, the Cellular Gatekeeper for Growth and Division. Cell. 1997; 88: 323–31.
15. Salmon S.E., Dalton W.S., Grogan T.M. et al. Multidrug-resistant myeloma: laboratory and clinical effects of verapamil as a chemosensitizer. Blood. 1991; 78: 44–50.
16. Brooks T.A., Hurley L.H. Targeting MYC Expression through G-Quadruplexes. Genes Cancer. 2010; 1(6): 641–9.
17. Dang C.V. MYC on the path to cancer. Cell. 2012; 149(1): 22–35. DOI: 10.1016/j.cell.2012.03.003
18. Gao P., Tchernyshyov I, Chang T.C. et al. c-myc suppression of miR-23a/b enhances mitochondrial glutaminase expression and glutamine metabolism. Nature. 2009; 458(7239): 762–5. DOI: 10.1038/nature07823
19. Kuzyk A., Mai S. c-MYC-induced genomic instability. Cold Spring Harb Perspect Med. 2014; 4(4): a014373. DOI: 10.1101/cshperspect.a014373
20. Karlsson A., Giuriato S., Tang F. et al. Genomically complex lymphomas undergo sustained tumor regression upon MYC inactivation unless they acquire novel chromosomal translocations. Blood. 2003; 101(7): 2797–803.
21. Mirabella F., Wu P., Wardell C.P. et al. MMSET is the key molecular target in t(4;14) myeloma. Blood Cancer J. 2013; 3(5): 114. DOI: 10.1038/bcj.2013.9
22. Chesi M., Bergsagel P.L. Molecular pathogenesis of multiple myeloma: basic and clinical updates. Int J Hematol. 2013; 97(3): 313–23. DOI: 10.1007/s12185-013-1291-2
23. Zhan F., Colla S., Wu X. et al. CKS1B, overexpressed in aggressive disease, regulates multiple myeloma growth and survival through SKP2- and p27Kip1- dependent and -independent mechanisms. Blood. 2007; 109(11): 4995–5001.
24. Smol T., Dufour A., Tricot S. et al. Combination of t(4;14), del(17p13), del(1p32) and 1q21 gain FISH probes identifi es clonal heterogeneity and enhances the detection of adverse cytogenetic profi les in 233 newly diagnosed multiple myeloma. Mol Cytogenet. 2017; 10: 26. DOI: 10.1186/s13039-017-0327-3
25. Lobry C., Oh P., Mansour M.R. et al. Notch signaling: switching an oncogene to a tumor suppressor. Blood. 2014; 123(16): 2451–9. DOI: 10.1182/blood-2013-08-355818
26. Colombo M., Mirandola L., Platonova N. et al. Notch-directed microenvironment reprogramming in myeloma: a single path to multiple outcomes. Leukemia. 2013; 27(5): 1009–18. DOI: 10.1038/leu.2013.6
27. Colombo M., Galletti S., Bulfamante G. et al. Multiple myeloma-derived Jagged ligands increases autocrine and paracrine interleukin-6expression in bone marrow niche. Oncotarget. 2016; 7(35): 56013–29. DOI: 10.18632/oncotarget.10820
28. Kishimoto T. The biology of interleukin-6. Blood 1989; 74: 1–10.
29. Frassanito M.A., Cusmai A., Iodice G., Dammacco F. Autocrine interleukin-6 production and highly malignant multiple myeloma: relation with resistance to drug-induced apoptosis. Blood. 2001; 97(2):483–9.
30. Gadó K., Domján G., Hegyesi H., Falus A. Role of interleukin-6 in the pathogenesis of multiple myeloma. Cell Biol Int. 2000; 24(4): 195–209.
31. Herrero A.B., Rojas E.A., Misiewicz-Krzeminska I. et al. Molecular Mechanisms of p53 Deregulation in Cancer: An Overview in Multiple Myeloma. Int J Mol Sci. 2016; 17(12): 2003. DOI:10.3390/ijms17122003
32. Simons A., Shaffer L.G., Hastings R.J. Cytogenetic Nomenclature: Changes in the ISCN 2013 Compared to the 2009 Edition. Cytogenet. Genome Res 2013; 141: 1–6. DOI: 10.1159/000353118
33. Dobin A., Davis C.A., Schlesinger F. et al. STAR: ultrafast universal RNA-seqaligner. Bioinformatics. 2013; 29(1): 15–21. DOI: 10.1093/bioinformatics/bts635
34. Love M.I., Huber W., Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014; 15(12): 550.
35. Rajkumar S.V., Dimopoulos M.A., Palumbo A. et al. International Myeloma Working Group updated criteria for the diagnosis of multiple myeloma. Lancet Oncol. 2014; 15(12): 538–48. DOI: 10.1016/S1470-2045(14)70442-5
36. Kodal J., Vedel-Krogh S., Kobylecki C. et al. TP53 Arg72Pro, mortality after cancer, and all-cause mortality in 105,200 individuals. Sci Rep. 2017; 7: 336. DOI: 10.1038/s41598-017-00427-x
37. Smadja N.V., Bastard C., Brigaudeau C. et al. Hypodiploidy is a major prognostic factor in multiple myeloma. Blood. 2001; 98(7): 2229–38.
38. Chen M.H., Qi C., Reece D., Chang H. Cyclin kinase subunit 1B nuclear expression predicts an adverse outcome for patients with relapsed/refractory multiple myeloma treated with bortezomib. Hum Pathol. 2012; 43(6): 858–64. DOI: 10.1016/j.humpath.2011.07.013
39. Stella F.,Pedrazzini E., Baialardo E. et al. Quantitative analysis of CKS1B mRNA expression and copy number gain in patients with plasma cell disorders. Blood Cells Mol Dis. 2014; 53(3): 110–7. DOI: 10.1016/j.bcmd.2014.05.006
40. Chauhan D., Uchiyama H., Akbarali Y. et al. Multiple myeloma cell adhesioninduced interleukin-6 expression in bone marrow stromal cells involves activation of NF-kappa B. Blood. 1996; 87(3): 1104–12.
ЭПИГЕНЕТИЧЕСКИЕ НАРУШЕНИЯ ПРИ ГЕМОБЛАСТОЗАХ
Вступление
Изучение хромосомных аберраций, ведущих к повреждению нормальной экспрессии генов при гемобластозах, позволило выявить, что в опухолевой клетке, кроме нарушения структуры ДНК, изменения обнаруживаются на эпигенетическом уровне, т.е. на уровне считывания генетической информации с участием белковых и нуклеотидных структур. При этом если значение генетических аберраций при этих заболеваниях в настоящее время достаточно хорошо изучено, эпигенетические нарушения, ассоциированные с развитием лейкемического фенотипа, стали предметом пристального внимания лишь в последнее время.
Что такое эпигенетика?
По современному определению, к эпигенетически обусловленным относят все унаследованные клеткой в процессе деления (митоза или мейоза) особенности регуляции экспрессии генов, не связанные непосредственно с изменением кода ДНК [1]. Осуществляют эпигенетическое влияние на генную экспрессию процессы, влияющие на степень конденсации хроматина – структуры-носителя генетической информации в клетке, состоящей из ДНК и особых белков-гистонов.
При уменьшении конденсации хроматина освобождается доступ факторов транскрипции к ДНК и становится возможным экспрессия генов, то есть последовательное образование РНК (транскрипция) и белка (трансляция). Ведущими процессами, регулирующими структуру хроматина, являются метилирование ДНК, ферментная модификация белков-гистонов и РНК-асссоциированное подавление транскрипции и трансляции [2]. Исследования последнего времени выявили устойчивую связь между всеми тремя компонентами эпигенетической регуляции, их способность к взаимной активации (рис. 1).
Оказалось, что нарушения в одной из систем неизбежно приводит к повреждению механизмов двух других, и, таким образом, вызывает неадекватную экспрессию или, наоборот подавление функционирования огромного количества генов. В связи с обнаружением различных видов нарушений этих процессов, появилась новая концепция развития целого ряда заболеваний – эпигенетическая, возникли термины «эпигенетические болезни» и «эпигенетическая терапия».
Рис. 1. Взаимосвязь механизмов эпигенетической регуляции.
Структура хроматина
Для лучшего понимания процессов считывания генетической информации и их регуляции следует вспомнить, что генетический материал в клетке существует в виде хроматина – структуры, состоящей из ДНК и белков – гистонов. Структурной единицей хроматина является нуклеосома, в состав которой входят часть спирали ДНК, размером 146 пар нуклеотидов (п.н.), совершающая виток в 1,7 раз вокруг белковой структуры – октамера, состоящей из гистонов.
Нуклеосомы связаны между собой участком ДНК около 50 п.н. [3]. Таким образом, хроматин образует динамическую структуру, позволяющую компактно расположить в ядре клетки около 2 м ДНК, сохранив при этом ее способность к функционированию. Для осуществления процесса транскрипции – образования РНК, белкам-транскрипционным факторам необходим доступ к регуляторным последовательностям ДНК. Доступ регулируется двумя взаимосвязанными процессами: метилированием ДНК и ацетилированием (и другими ферментными модификациями) гистонов, которые будут рассмотрены ниже. Схематически процесс изображен на рис. 2.
Рис. 2. Изменение структуры хроматина в процессе метилирования ДНК и ацетилирования гистонов
Метилирование ДНК
Метилированием называется присоединение группы СН3 к атому углерода в 5 позиции азотистого основания цитозина, одного из четырех нуклеотидов, составляющих структуру цепи ДНК (рис. 3). Процесс метилирования является одним и фундаментальных процессов эпигенетического подавления экспрессии генов, закрепленным в процессе эволюции [4,5]. Образование метилцитозина ведет к конденсации хроматина, препятствует присоединению транскрипционных факторов к своим мишеням на ДНК и, таким образом, ингибирует считывание информации.
В процессе естественного развития метилирование является ведущим механизмом осуществления последовательной ткане-специфической экспрессии генов и правильного развития эмбриона, инактивации одной Х-хромосомы у женщин и осуществления механизма импринтинга, позволяющего «замалчивать» один из родительских генов в диплоидном наборе хромосом.
Метилированию в норме подвергается от 2 до 7% всех цитозиновых остатков ДНК клетки. При этом в 70% случаев цитозин метилируется в составе динуклеотидов С-G (CpG). СpG участки, как правило, представляют собой фрагменты ДНК длиной более чем 500 пар нуклеотидов (п.н.) и базируются в зонах инициации считывания информации – промотерах – более чем 40% генов млекопитающих [6,7].
Осуществляется метилирование с помощью ферментов ДНК-метилтрансфераз (DNMT). Метилтранферазы можно условно разделить на две группы – метилирующие ДНК de novo, то есть в тех участках, где ранее не было метилцитозина (DNMT3A и DNMT3B), и «поддерживающие» метилирование в дочерней цепи ДНК, образующейся в процессе репликации (DNMT1), сохраняя, таким образом, структуру, присущую материнской цепи.
Рис. 3. Процесс метилирования
Нарушения метилирования при развитии злокачественных новообразований возникают, как правило, на ранних стадиях. Общий низкий уровень метилирования цитозина (гипометилирование) при этом сочетается с обратным процессом -гиперметилированием CpG участков в промотерах генов-супрессоров опухоли [8,9]. Тотальное гипометилирование ведет к повышению экспрессии протоонкогенов, генов, кодирующих ростовые факторы, а также целого ряда генов, таких, как ген активатора плазминогена урокиназного типа (PLAU), ген гепараназы и кальций-связывающего протеина (S100A4), способствующих метастазированию путем проникновения клетки через стенки кровеносных и лимфатических сосудов [10].
Важную роль играет также гипометилирование специфических олигонуклеотидных элементов – ретротранспозонов – имеющих вирусное происхождение. Ретротранспозоны представляют собой обратные транскрипты вирусной РНК, в норме достаточно произвольно встроенные в структуру хозяйской ДНК. Обычно эти элементы метилированы и поэтому неактивны. Зачастую ретротранспозоны встраиваются в регуляторные зоны генов – интроны. Активация этих элементов при опухолевой трансформации ведет к повышению генетической нестабильности и нарушению нормальной экспрессии хозяйских генов [11].
Причины гипометилирования ДНК в злокачественных клетках до конца не ясны. Гипотеза, предполагающая причиной общий или частичный дефицит метилирующих ферментов не объясняет одновременное гиперметилирование промотеров целого ряда генов – супрессоров опухоли. Некоторые исследователи предполагают, что причиной тотального гипометилирования ДНК в злокачественных клетках является образование определенных изоформ фермента ДНК-метилтрансферазы типа DNMT3B.
Эта форма фермента, связываясь с CpG участками промотеров генов, не приводит к их метилированию, но при этом препятствует связи этих участков с активными формами метилтрансфераз [12,13]. В то же время этот тип фермента и, в меньшей степени, DNMT1 способствуют активному метилированию и подавлению экспрессии генов-супрессоров опухоли. При ОМЛ характерным является высокий уровень экспрессии как DNMT3B, так и DNMT1, что сопровождается гиперметилированием промотеров таких генов – опухолевых супрессоров, как ингибиторы циклин-зависимой киназы (CDKN2A и CDKN2B), ген эстрогенового рецептора I (ESR1), и ген ретинобластомы (RB1) [14]. В последние годы появились данные целого ряда исследований, доказывающих, что ДНК-метилтрансферазы участвуют в образовании белковых комплексов, подавляющих транскрипцию, которые возникают при участии химерных протеинов, таких, как PML/RARA и AML/ETO (1, 15, 16, 17).
Открытие роли метилирования в онкогенезе привело к широкому внедрению препаратов-ингибиторов ДНК-метилтрансфераз. Наиболее известным из них является 5-аза-2’-деоксицитидин (Дакоген), который, встраиваясь в цепочку ДНК вместо цитозина, образует прочную ковалентную связь с ДНК-метилтрансферазами, препятствуя, таким образом, метилированию и снимая блок с репрессированного ранее гена [18].
Модификации гистонов
Следующим важнейшим механизмом эпигенетической регуляции является ферментная модификация основных белков хроматина – гистонов. Гистоны образуют ядро нуклеосомы, вокруг которого формируется виток спирали ДНК. Существует 4 вида белков-гистонов. При формировании ядра нуклеосомы они образуют следующие тесно связанные структуры: тетрамер из протеинов Н3 и Н4 и 2 димера из Н2А и Н2В [19]. Схематически ядро нуклеосомы представлено на рис. 4. Гистон Н1 у высших эукариот осуществляет связывание нуклеосом между собой, способствуя конденсации хроматина [20].
Рис. 4. Модель ядра нуклеосомы (вид сверху): тетрамер Н3 и Н4 и один из димеров Н2А и Н2В (второй димер расположен под первым)
Структура каждого гистона включает глобулярное ядро, обладающее специфическим мотивом – завиток-петля-завиток, осуществляющим димеризацию и свободный «хвостовой» домен, включающий аминокислоты лизин, аргинин и серин. Этот домен обладает положительным электрическим зарядом, и, взаимодействуя с анионными группами цепи ДНК определяет стабильность структуры нуклеосомы [21].
Ацетилирование или метилирование свободной аминогруппы лизина приводит к изменению общего заряда белка и, соответственно, изменению структуры хроматина – освобождается доступ транскрипционных факторов к ДНК (см. рис. 3). Такие же изменения происходят при фосфорилировании гидроксильного остатка серина. В целом, в настоящее время известно более 50 позиций в аминокислотной структуре гистонов, которые могут быть подвержены ферментной модификации (метилированию, ацетилированию, фосфорилированию, АДФ-рибозилированию и т.д.) [22].
Оказалось, что каждое из этих изменений является своеобразным знаком для привлечения различных регуляторных комплексов, каждый из которых определяет особенности транскрипции. Таким образом, сочетание различных модификационных процессов создает так называемый «гистоновый код», во многом определяющий сущность и порядок считывания генетической информации. Оказалось, что информационные возможности гистонового кода едва ли не больше возможностей классического кода ДНК [23, 24].
Процессы ацетилирования и деацетилирования, наиболее изученные к настоящему времени, осуществляются специфическими группами ферментов – гистонацетилазами (или ацетилтрансферазами), или HAT и гистон-деацетилазами, или HDAC. Гистонацетилазы работают, как правило, в составе больших комплексов, которые, зачастую, способны изменять их активность. Интересно, что целый ряд известных транскрипционных факторов таких, как CBP и p300, TAFII250, SRC-1 имеют последовательности, гомологичные структуре HAT, и обладают способностью к самостоятельному ацетилированию гистонов.
Антагонисты ацетилаз – деацетилазы составляют 3 класса, отличающиеся по своей структуре и несколько различающиеся по функции. Классической функцией HDAC является репрессия транскрипции с помощью удаления ацетиловой группы с лизиновых остатков и, соответственно, конденсации структуры нуклеосомы [25]. Однако в последнее время появились данные, что в ряде случаев деацетилирование необходимо для успешного функционирования транскрипционно активных генов [17]. Эти данные подтверждают необычайную сложность и, зачастую, парадоксальность гистонового кода.
Участие гистондеацетилаз в лейкемогенезе хорошо изучено на классических примерах острого промиелоцитарного лейкоза (ОПЛ) и ОМЛ, несущих транслокацию t(8;21) или сопровождающихся аномалиями 16 хромосомы.
При ОПЛ в 95% случаев образуется транслокация t(15;17), ведущая к появлению химерного белка PML/RARA. В норме продукт одного из генов-участников транслокации ядерный рецептор RARA образует комплекс с корепрессорами (SMRT, N-CoR, Sin3a) и гистондеацетилазой HDAC, блокирующий транскрипцию регулируемых генов. Этот комплекс разрушается при присоединении естественного лиганда RARA – ретиноевой кислоты – и транскрипция становится возможна.
При образовании химерного протеина PML/RARA репрессирующий комплекс не реагирует на ретиноевую кислоту в физиологических концентрациях. Осуществляемое гистондеацетилазой HDAC деацетилирование гистонов служит также сигналом для активации других ферментов: гистоновых метилтрансфераз и ДНК-метилтрансфераз, также включающихся в репрессирующий комплекс (рис. 5).
Таким образом осуществляется взаимодействие между двумя механизмами эпигенетической регуляции: модификацией структуры гистонов и метилированием ДНК. Фармакологические концентрации ретиноевой кислоты, в тысячи раз превышающие физиологические, способны вызвать диссоциацию репрессирующего комплекса и запустить механизм дифференцировки [1]. Одновременно ретиноевая кислота приводит к активации «лиганда смерти» – TRAIL (TNF-related apoptosis-inducing ligand), являющегося ключевым компонентом механизма апоптоза [26].
Полностью трансретиноевая кислота (АТРА), как «дифференцирующий агент» в терапии ОПЛ представляет собой типичный пример препарата с эпигенетическим действием: не изменяя структуру ДНК, она приводит к разрушению репрессирующего комплекса, деацетилирующего гистоны регуляторных элементов генов, ответственных за дифференцировку клеток.
Тем не менее, несмотря на достигнутые успехи в понимании механизма лейкемогенеза при ОПЛ остается ряд вопросов, требующих дальнейших исследований: 1) взаимодействует ли химерный онкоген при действии АТРА абсолютно с тем же активирующим комплексом, что и нормальный ядерный рецептор; 2) какова роль доменов, принадлежащих гену PML; 3) значение экспрессии реципрокного химерного протеина и т.д.
Рис 5. Процесс нормального функционирования ядерного рецептора RARA и ингибирование транскрипции химерным протеином PML/RARA
В незначительном числе случаев ОПЛ, сопровождающихся образованием транслокации t(11;17) (PLZF/RARA) и t(11;17) STAT5b/RARA, химерные протеины образуют более прочную связь с репрессирующим комплексом за счет дополнительных сайтов в структуре второго участника химерного белка – продукта генов PLZF или STAT5.
Эта связь не разрушается в присутствии высоких концентраций ретиноевой кислоты именно за счет дополнительных связей репрессирующего комплекса. Следует отметить, что в эксперименте in vitro комбинация полностью трансретиноевой кислоты и ингибиторов гистондеацетилаз, разрушающих дополнительные связи химерных протеинов, позволяет добиться дифференцировки лейкемических клеток [27].
При ОМЛ, сопровождающихся t(8;21) или аномалиями 16 хромосомы с вовлечением генов ЕТО(MTG8), AML1(RUNX1) и CBFβ, происходит нарушение образования нормального транскрипционного комплекса CBF. Этот комплекс состоит из продуктов генов c/EBPα, AML1(RUNX1) и CBFβ и обладает гистонацетилазной активностью, потенцируя экспрессию большой группы генов, отвечающих за нормальную дифференцировку гемопоэтических клеток.
При возникновении генетических аберраций, изменяющих нормальную структуру любого из белков комплекса CBF, нарушается его связь с фактором-гисонацетилазой р300, коактиватором и ядерным рецептором TIF2. При транслокации t(8;21) ведущую роль в нарушении комплекса играет та часть химерного протеина AML/ETO, которая считывается со второго участника транслокации – гена ЕТО(MTG8). В норме этот ген экспрессирует белок, участвующий в репрессирующем комплексе и связывающий уже известные нам гистондеацетилазу HDAC1 и корепрессоры N-CoR и Sin3a.
За счет доменов белка ЕТО, осуществляющих эти связи, химерный протеин AML/ETO приобретает функции репрессора транскрипции и подавляет гены в норме активирующиеся AML1 [28]. В последнее время исследуется огромное количество химических соединений, обладающих функцией ингибиторов гистондеацетилаз, однако большинство из них пока проходят первую или вторую фазу клинических испытаний [29].
Весьма сложными нарушениями сопровождаются повреждения нормального функционирования гена MLL, кодирующего протеин, который обладает в том числе и гистон-метилтрансферазной активностью. Основной функцией этого белка является участие в огромном транскрипционном комплексе, в состав которого входят по меньшей мере 29 других белков, вовлеченных как в ацетилирование, так и в деацетилирование гистонов, в метилирование гистонов и ДНК [30].
Этот комплекс осуществляет регуляцию транскрипции генов гомеобокса, участвующих в эмбриональном развитии и экспрессирующихся в ранних предшественниках гемопоэза. В различных транслокациях с участием MLL (которых известно около 60 для всех видов лейкемии) образуются химерные протеины, потерявшие метилтрансферазную активность за счет отсутствия соответствующего домена в составе химерных генов. Скорее всего, механизм лейкемогенеза связан в данной ситуации с нарушением функционального баланса образующегося транскрипционного комплекса, а также, несомненно, от функций, привнесенных вторым участником транслокации [16].
РНК-ассоциированное подавление экспрессии генов
Роль РНК в подавлении экспрессии генов в последнее время привлекает все большее внимание исследователей. Оказалось, что РНК-индуцированная ингибиция может осуществляться на различных этапах считывания генетической информации. В основе этого процесса лежит феномен интерференции – способность двухцепочечной РНК к эффективному ингибированию экспрессии генов. Двухцепочечная молекула РНК расщепляется на фрагменты длиной 21-25 нуклеотидов, образуя так называемую siRNA (small interfering RNA) [31].
Такие молекулы РНК способны образовывать комплексы с протеинами (RISC – RNA induced silencing complex) и вызывать деградацию матричной РНК, то есть останавливать генную экспрессию на посттранскрипционном уровне – не допускать синтеза белка после успешного образования РНК [32]. Ранее этот механизм считался более характерным для клеток растений, однако в последнее время появились данные, что посттранскрипционная ингибиция возникает и в клетках млекопитающих [31].
РНК участвует также и в подавлении экспрессии генов на уровне транскрипции, вызывая РНК-зависимое метилирование ДНК. Образующиеся из двухцепочечной РНК короткие фрагменты имеют структуру, гомологичную промотерам ряда генов и способны индуцировать метилирование этих промотеров, присоединяясь к ним и активируя ДНК-метилтрансферазы [33].
Более того, siRNA способны вызывать метилирование гистонов, подтверждая тем самым взаимосвязь механизмов эпигенетической регуляции между собой, хотя пока до конца не ясен порядок этих событий и их взаимозависимость [34]. Вполне возможно, что возникающее первоначально метилирование лизина в структуре гистона Н3 ведет в дальнейшем к активации ДНК-метилтрансфераз [35]. Не совсем ясен пока и механизм доставки siRNA к геномной ДНК.
В настоящее время рассматривается несколько возможных вариантов участия siRNA в ингибировании экспрессии генов. Первый механизм – прямое связывание РНК со специфическим транспортным белком Argonaute 2 (возможно в комплексе с гистондеацетилазами) и непосредственное связывание структуры с комплементарной последовательностью в промотере гена с дальнейшим включением механизма деацетилтрования гистонов и остановкой транскрипции.
Два других механизма предполагают связывание siRNA с известными репрессирующими комплексами – Mi2/NuRD и Sin3. Первый из них включает гистондеацетилазы (HDAC 1, 2) и ДНК-метилтрансферазу (DNMT1), т.е. обладает способностью к метилированию ДНК и деацетилированию гистонов. Второй имеет в своем составе гистондеацетилазы (HDAC 1, 2) [35].
Схематически все три механизма изображены на рис. 6.
Рис. 6. Возможные механизмы РНК-ассоциированной ингибиции экспрессии генов.
Следует отметить, что пока нет достоверных данных о роли РНК-опосредованного механизма ингибиции генной экспрессии непосредственно в лейкемогенезе, однако исследование этого феномена будет иметь огромное значение для понимания взаимосвязи различных видов эпигенетической регуляции между собой.
Заключение
Изучение эпигенетической регуляции в последние годы позволило не только добиться выдающихся успехов в понимании механизмов канцерогенеза в целом и лейкемогенеза в частности, но и открыть новую область в терапии – эпигенетическую терапию злокачественных заболеваний. Если генетические нарушения не могут быть скорректированы с помощью влияния извне, и ведут к необратимой потере функции поврежденного гена, нарушения эпигенетической регуляции потенциально обратимы, что позволяет рассчитывать на возможность коррекции имеющихся дефектов с помощью правильно подобранного терапевтического воздействия.
Применение так называемой направленной терапии, веществ, модифицирующих действие гистондеацетилаз и ДНК-метилтрансфераз, синтетических siRNA сделает возможным изменение цикла жизни злокачественной клетки, возврат ее к нормальной дифференцировке и естественному апоптозу, что, несомненно, приведет к возникновению качественно нового этапа в лечении онкологических заболеваний.
Литература:
- Altucci L., Clarke N., Nebbioso A et al. Acute myeloid leukemia: Theraputic impact of epigenetic drugs Int Journ of Biochem and Cell Biol, 2005, 37, 1752-1762
- Egger G., Liang G, Aparicio A., Jones PA. Epigenetics in human disease and prospects for epigenetic therapy. Nature, 2004, 429, 457-463
- Kornberg RD, Lorch U. Twenty-five years of nucleosome, fundamental particle of the eukaryote chromosome. Cell, 1999, 98, 285-294
- Holiday R, Pugh JE. DNA modification mechanism and gene activity during development. Science, 1975, 187, 226-232
- Riggs AD. X inactivation, differentiation and DNA methylation. Cytogenet Cell Genet, 1975, 14, 9-25
- Takai D, Jhones PA. Comprehensive analysis of CpG islands in human chromosomes 21 and 22. Proc Natl Acad Sci USA , 2002, 99, 3740-3745
- Monk M. Epigenetic programming of different gene expression in development and evolution. Dev Genet 1995, 17, 188-197
- Baylin SB, Esteller M, Rountree MR et al. Aberrant patterns of DNA methylation, chromatin formation and gene expression in cancer Hum Mol Genet 2001, 10, 687-692
- Laird PW. Cancer epigenetic. Hum Mol Genet 2005, 14, 65-76
- Luczak MW, Jagodzinski PP. The role of DNA methylation in cancer development. Fol Hist et Cytogen 2006, 44, 143-154
- Carnell AN, Goodman JI. The long (LINEs) and short (SINEs) of it: altered methylation as a precursor of toxicity. Toxicol Sci 2003, 75, 229-235
- Saito Y, Kanai Y, Sakamoto M et al. Expression of mRNA of DNA methyltransferases and methyl-GpC-binding proteins and DNA methylation status on GpC islands and pericentromeric satellite regions during human hepatocarcinogenesis. Hepatology, 200, 33, 561-568
- Weisenberger DJ, Velicescu M, Cheng JC et al. Role of DNA-methyltransferase variant DNMT3b3 in DNA methylation. Mol Cancer Res 2004, 2, 62-67
- Melki JR, Vincent PC, Clark SJ. Concurent DNA hypermethylation of multiple genes in acute myeloid leukemia. Cancer Res 1999, 59, 3730-3740
- Liu S, Shen T, Huynh L. Interplay of RUNX1/MTG8 and DNA methyltransferase 1in acute myeloid leukemia. Cancer Res 2005, 65, 1277-1284
- Di Croce L. Chromatin modifying activity of leukemia associated fusion proteins. Human Mol Genet 2005, 14, 77-84
- Ducasse M, Brown M. Epigenetic aberrations and cancer. Molecular Cancer 2006, 5, 60-70
- Christmas JK. 5-Azacytidine and 5-aza-2-deoxycytidine as inhibitors of DNA methylation: mechanistic studies and their implications for cancer therapy. Oncogene 2002, 21, 5483-5495
- Luger K, Madeer AW, Richmond RK et al. Crystal structure of the nucleosome core particle at 2.8 resolution. Nature 1997, 389, 251-260
- Francis NJ, Kingdton RE, Woodcock CL. Chromatin compaction by a Polycomb group protein complex. Science 2004, 306, 1574-1577
- Hayes JJ, Clark DJ, Wolffe AP. Histone contributions to the structure of DNA in the nucleosome. Proc Natl Acad Sci USA 1991, 88, 6829-6833
- Jenuwein T. The epigenetic magic of histone lysine methylation. Febs J 2006, 273, 3121-3135
- Jenuwein T., Allis CD. Translating the histone code. Science 2001, 293, 1074-1080
- Strahl BD, Allis CD. The language of covalent histone modifications. Nature 2000, 403, 41-45
- Gregoretti IV, Lee YM, Goodson HV. Molecular evolution of histone deacetylase family: functional implications of phylogenetic analysis. J Mol Biol 2004, 338, 17-31
- Clarke N, Nebbioso A, Altucci L, Gronmeyer H. TRAIL. At the center of drugable anti-tumor pathways. Cell Cycle 205, 4, 914-918
- Lin RG, Nagy L, Inoue S. et al. Role of histone deacetylase complex in acute promyelocytic leukemia. Nature 1991, 391, 811-814
- Hiebert SW, Reed-Inderbitzin EF, Amann J et al. The t(8;21) fusion rotein contacts co-repressors and histone deacetylases to repress the transcription of the p14ARF tumor repressor. Blood Cells Mol Dis 2003, 30, 177-183
- Marks PA, Miller T, Richon VM. Histone deacetylases. Curr Opin Pharmacol 2003, 3, 344-351
- Nakamura T., Mori T, Tada S et al. ALL-1 s a histone methyltransferase that assembles a supercomplex of proteins involved in transcriptional regulations. Mol Cell, 10, 1119-1128
- Elbashir SM, Lendeckel W, Tuschl T. RNA interference is mediated by 21- and 22-nucleotide RNAs. Genes and Development 2001, 15, 188-200
- Zeng Y, Cullen BR. RNA interference in human cells is restricted to the cytoplasm. RNA 2002, 8, 855-860
- Morris KV. Small interference RNA-induced transcriptional gene silencing in human cells. Science 2004
- Kawasaki H, Taira K. Induction of DNA methylation and gene silencing in human cells. Nature 2004
- Kawasaki H, Taira K, Morris KV. siRNA induced transcriptional gene silencing in mammalian cells. Cell Cycle 2005, 4, 442-448
#62 Новое решение Visium от 10x Genomics для анализа пространственной экспрессии генов
17.01.2020
Существующие технологии 10x Genomics уже сейчас позволяют анализировать геномы и транскриптомы единичных клеток путем NGS-секвенирования. Но компания решила пойти еще дальше и разработала технологию Visium для анализа распределения экспрессионных профилей клеток в пространстве. Это совершенно новый подход к исследованию динамики молекулярных процессов в биологических образцах.
Компоненты системы Visium
Как и прочие протоколы от 10x Genomics, Visium представляет собой готовое решение и включает:
- Слайды для визуализации образцов и захвата транскриптов,
- Реагенты для захвата транскриптов и синтеза кДНК,
- Слайды и реагенты для оптимизации условий пермеабилизации,
- Реагенты для подготовки РНК-библиотеки (3’),
- Программное обеспечение для биоинформатической обработки данных.
Описание технологии Visium
Общая схема подготовки NGS-библиотеки для анализа пространственной экспрессии генов с помощью технологии Visium выглядит следующим образом.
|
Рис. 1. Общая схема технологии Visium. |
Протокол начинается с подготовки 4-х срезов ткани толщиной 10-50 мкм и их размещении на специальных слайдах в 4-х зонах захвата транскриптов.
|
Рис. 2. Устройство слайда Visium для анализа пространственной экспрессии генов. |
Каждая такая зона состоит из 5 000 точек захвата транскриптов. Точка захвата представляет собой группу олигонуклеотидов, закрепленных на подложке. Диаметр каждой точки – 55 мкм, а расстояние между ними – 100 мкм. В зависимости от типа ткани и толщины среза на одну точку может приходиться 1-10 клеток.
Последовательность олигонуклеотидов включает в себя адаптер R1 (стандартный адаптер TruSeq), баркод Spatial, метку UMI и поли-dT-хвост. Баркод Spatial уникален для каждой точки захвата транскриптов.
После размещении среза ткани на слайде ее фиксируют и окрашивают гематоксилин-эозином. Подготовленный препарат визуализируют с помощью микроскопа и фотографируют.
|
Рис. 3. Микроскопия и фотографирование подготовленного препарата срезов ткани. |
Затем слайд вставляют в специальный адаптер, имеющий отверстия напротив зон захвата. Таким образом образуются лунки, дном которых выступают срезы тканей и куда можно вносить реагенты.
|
Рис. 4. Слайд Visium в адаптере. |
Следующий этап протокола – пермеабилизация клеток. Ее выполняют путем добавления в лунки специальных ферментов и инкубации адаптера со слайдом при 37°C. При этом мРНК покидают образец и связываются с поли-dT-хвостами олигонуклеотидов зон захвата.
|
Рис. 5. Пермеабилизация клеток среза. |
Далее следуют этапы синтеза кДНК методом обратной транскрипции с переключением матрицы, денатурации и удаления мРНК, синтеза второй цепи кДНК, денатурации и переноса второй цепи кДНК в пробирки.
|
Рис. 6. Синтез первой и второй цепи кДНК на слайде Visium. |
На последней стадии вторую цепь кДНК используют для подготовки NGS-библиотек, включающую в себя:
- Оценку качества кДНК,
- Амплификацию кДНК,
- Фрагментацию / восстановление концов / 3′-аденилирование концов,
- Лигирование адаптеров,
- Индексирующую ПЦР,
- Контроль качества библиотеки.
|
Рис. 7. Структура фрагментов библиотеки, подготовленной по технологии Visium. |
Готовую библиотеку можно секвенировать на любой платформе Illumina®. Глубина прочтений должна составлять не менее 50 000 пар ридов на одну зона захвата мРНК (то есть на один баркод Spatial). Режим прочтения – 28 x 120 н.п.
Оптимизация условий пермеабилизации в протоколе Visium
Перед тем как приступать к исследованию новых типов тканей, рекомендуется оптимизировать длительность ферментативной пермеабилизации для максимально эффективного захвата мРНК. Для этих целей 10x Genomics разработал специальный набор слайдов и реагентов.
Слайды для оптимизации имеют уже 8 зон захвата мРНК, но размещают на них всего 7 срезов ткани. Одна зона предназначена для положительного контроля, в качестве которого используют референсную мРНК. Закрепленные на подложке олигонуклеотиды не содержат баркодов и состоят из одного поли-dT-хвоста.
После стандартной подготовки, окраски, фиксации и фотографировании срезов ткани на слайде его вставляют в адаптер и приступают к подбору условий пермеабилизации.
Для этого в лунку без среза ткани для положительного контроля добавляют референсную РНК, в лунку со срезом для положительного контроля не добавляют ничего. В остальные же лунки через определенные промежутки времени добавляют ферментативную смесь для пермеабилизации, инкубируя слайд при 37° C.
Далее лунки промывают и добавляют в них реакционную смесь для обратной транскрипции, которая содержит дЦТФ, меченный флуоресцентным красителем Cyanine 3. И в ходе обратной транскрипциии синтезируется флуоресцентно меченная кДНК.
На следующем этапе с помощью ферментов со слайдов удаляют ткани и методом флуоресцентной микроскопии оценивают эффективность захвата мРНК при разной длительности пермеабилизации. Наилучшие условия используют в дальнейшем при работе с опытными образцами.
| Рис. 8. A — Срезы ткани на слайде Visium для оптимизации условий пермеабилизации.B — Оценка зависимости эффективности захвата мРНК от длительности пермеабилизации: Pos и Neg – положительный и отрицательный контроли; 3, 6, 12, 18, 24 и 30 – длительность пермеабилизации в минутах. |
Обработка данных секвенирования библиотек пространственной экспрессии Visium
Первичную и вторичную обработку данных секвенирования выполняют с помощью бесплатного программного обеспечения Space Ranger и Loupe Browser.
ПО осуществляет выравнивание полученных ридов и фотографий, сделанных на этапе подготовки препарата. Баркод Spatial помогает определить, в какой точке был захвачен тот или иной транскрипт.
Результаты обработки можно вывести в виде наглядных отчетов. Они содержат информацию об общем числе точек, в которых произошел захват, числе ридов, приходящихся на одну точку захвата, числе генов, экспрессирующихся в точках захвата, и многие другие показатели.
|
|
| Рис. 9. Пример отчета, сформированного программным обеспечением Space Ranger. |
Также можно визуализировать пространственную экспрессию конкретных генов и кластеризировать точки захвата на основании экспрессионных профилей нескольких генов.
| Рис. 10. Результаты анализа пространственной экспрессии генов в головном мозге мыши по технологии Visium: A – корональный срез, окрашенный гематоксилин-эозином, B – пространственное распределение меток UMI, C – пространственное распределение экспрессии генов, D – пространственная кластеризация экспрессии 8 генов. |
|
Рис. 11. Пространственная экспрессия генов Hpca (A) и Selenow (B) в головном мозге мыши. Транскрипты обоих генов хорошо представлены в гиппокампе (белый прямоугольник), что согласуется с известными паттернами экспрессии. |
Типы тканей, совместимые с технологией Visium
Технология совместима с различными тканями человека, мыши и крысы. Их фиксацию можно выполнять метанолом.
Ниже перечислены типы тканей, совместимость которых с системой Visium была валидирована:
| Ткань | Толщина среза | Ткань | Толщина среза |
| Человек | Мышь | ||
| Головной мозг (мультиформная глиобластома) | 10 мкм | Головной мозг (здоровый) | 10 мкм |
| Грудь (здоровая) | 10-20 мкм | Глаза (здоровые) | 10 мкм |
| Грудь (инвазивная протоковая карцинома) | 10-20 мкм | Сердце (здоровое) | 10 мкм |
| Грудь (инвазивная лобулярная карцинома) | 10-20 мкм | Почки (здоровые) | 10 мкм |
| Сердце (здоровое) | 10 мкм | Толстый кишечник (здоровый) | 10 мкм |
| Почки (здоровые) | 10 мкм | Печень (здоровая) | 10 мкм |
| Почки (нефрит) | 10 мкм | Легкие (здоровые) | 10 мкм |
| Толстый кишечник (колоректальный рак) | 10-20 мкм | Яичники (здорове) | 10 мкм |
| Легкие (папиллярная карцинома) | 10 мкм | Квадрицепс (здоровый) | 10 мкм |
| Лимфоузлы (здоровые) | 10 мкм | Тонкий кишечник (здоровый) | 10 мкм |
| Лимфоузлы (воспаленные) | 10 мкм | Селезенка (здоровая) | 10 мкм |
| Яичники (опухоль) | 10 мкм | Желудок (здоровый) | 10 мкм |
| Селезенка (воспаленная) | 10 мкм | Семенники (здоровые) | 10 мкм |
| Крыса | Щитовидная железа (здоровая) | 10 мкм | |
| Головной мозг (здоровый) | 10 мкм | Язык (здоровый) | 10 мкм |
| Сердце (здоровое) | 10 мкм | ||
| Почки (здоровые) | 10 мкм | ||
Информация для заказа
Каталог →
| Артикул | Наименование |
| 1000184 | Набор слайдов и реагентов для пространстраственной экспрессии генов Visium Spatial Gene Expression Slide & Reagent Kit, 16 реакций |
| 1000187 | Набор слайдов и реагентов для пространстраственной экспрессии генов Visium Spatial Gene Expression Slide & Reagent Kit, 4 реакции |
| 1000193 | Набор слайдов и реагентов для оптимизации срезов ткани Visium Spatial Tissue Optimization Slide & Reagent Kit, 4 реакции |
| 1000194 | Набор дополнительных аксессуаров Visium Accessory Kit |
| 1000200 | Стартовый набор Visium для пространственного анализа экспрессии Visium Spatial Gene Expression Starter Kit |
| 1000215 | Набор индексов для библиотеки пространственной экспрессии Dual Index Kit TT, Set A 96 реакций |
Английские восклицания!!! Предложения, начинающиеся с How и What
Что такое восклицание
Чудесно! Какой прекрасный день! Изучая английский, мы, конечно же, должны стремиться к естественности разговорной речи (ведь каждый из нас мечтает, чтобы хоть раз в жизни нас приняли за носителя английского). Сегодня мы изучим английские восклицания, благодаря которым ваша английская речь будет звучать более натурально — кроме того, вы сможете более точно выражать свои эмоции.
How wonderful! What a nice day it is! — это примеры восклицаний. Восклицание — это экспрессия, крик души, слова, которые вырываются из сердца, когда мы испытываем сильные эмоции:
Whoa! What a big car! | Ух ты! Какая большая машина! |
How beautiful! | Как красиво! |
Мы рассмотрим два типа восклицаний: те, которые начинаются со слова how («как») и те, которые начинаются словом what («что»).
How | + adjective |
«Как» | прилагательное |
Восклицания, которые начинаются с наречия how («как»)
Восклицания первого типа состоят из наречия how и прилагательного (adjective).
Предположим, ваш друг-астронавт рассказывает вам о своей работе: о трудностях жизни на орбитальной станции, о красоте видов Земли из космоса… И у вас само собой вырывается: How exciting! («Как интересно!»).
Или же друзья пригласили вас познакомиться с их новорожденным сыном, как тут удержишься от восхищенного How cute! («Какой милый!»).
А что, если вы попали в дом с привидениями? How strange, how very strange! («Как странно, как это странно!») — скажете вы.
Восклицания, которые начинаются с местоимения what («что»)
Вторая «восклицательная» конструкция чуть сложнее:
What | + a(n) | + adjective | + subject |
«Что» | неопр. артикль | прилагательное | подлежащее |
Неопределенный артикль a может иметь форму an (если за ним следует слово, начинающееся с гласного звука) или пропадать, в случае если существительное стоит во множественном числе.
What a strange man! | Что за странный человек! |
What a beautiful child! | Что за прелестное дитя! |
Wow! What an exciting movie! | Ух ты! Что за интересное кино! |
Как можно преобразовать повествовательное предложение в восклицательное
«Превращение» полного повествовательного предложения происходит по следующей формуле:
Subject | + verb | + adjective | → | How | + adjective | + subject | + verb |
Подлежащее | сказуемое | прилагательное | «Как» | прилагательное | подлежащее | сказуемое |
Пример:
She is pretty. → How pretty she is! | Она хорошенькая. → Как она хороша! |
Аналогично, конструкция с what приобретает следующий вид:
Subject | + verb | + adjective | → | What | + a(n) | + adjective | + subject | + verb |
Подлежащее | сказуемое | прилагательное | «Что» | неопр. артикль | прилагательное | подлежащее | сказуемое |
Пример:
You make a cute couple. → What a cute couple you make! | Вы прекрасная пара. → Вы такая прекрасная пара! |
Напоминаем, что в случае, если речь идет о множественном числе, неопределенный артикль исчезает:
Grandma, what big eyes do you have! | Бабушка, какие у тебя большие глаза! |
В завершении хочется отметить, что восклицательные предложения характерны скорее для разговорного языка и художественной прозы — в деловой речи использование восклицаний не допускается.
Читаем дальше:
Виды вопросов в английском языке
Что вы не знали о фрагментах предложений в английском
Как правильно объединять простые предложения в сложные
Уровень в сыворотке и почечная экспрессия молекул межклеточной адгезии ICAM-1 у больных с диабетической нефропатией | Бондарь
Диабетическая нефропатия (ДН) – одно из наиболее частых и прогностически неблагоприятных осложнений сахарного диабета (СД). В последние годы ДН заняла ведущую позицию среди причин терминальной почечной недостаточности в развитых странах. В Российской Федерации дополнительные прямые расходы на лечение одного пациента с начальными стадиями ДН составляют 245 $ в год, при наличии ХПН – 2012 $ в год (без учета расходов на гемодиализ и трансплантацию почки). При использовании последних методов стоимость лечения может достигать 45 тыс. $ в год [1].
Ведущими факторами, определяющими развитие и прогрессирование ДН, признаны гипергликемия и связанные с ней метаболические расстройства [2]. Наряду с этим, в последние годы обсуждается роль воспалительных реакций в формировании данного осложнения. Показана роль мигрирующих в почки провоспалительных клеток, прежде всего моноцитов/макрофагов, в развитии гломерулярного и интерстициального фиброза при СД [3–6]. Миграция мононуклеаров и реализация их активности в почках осуществляются в тесной кооперации с резидентными клетками и внеклеточным матриксом. Важнейшую роль при этом играют цитокины и молекулы адгезии [5, 7–9].
Среди адгезивных белков большое внимание привлекли молекулы межклеточной адгезии ICAM-1. Они обеспечивают фиксацию лейкоцитов на поверхности эндотелия и их последующий выход из сосудистого русла в ткани. Данный процесс определяется взаимодействием ICAM-1 с β2-интегринами лейкоцитов [10]. Экспрессия ICAM-1 обнаружена на многих типах клеток, включая эндотелиоциты, гранулоциты, макрофаги, фибробласты и др. В организме ICAM-1 существуют в мембрано-связанной и растворимой форме. Последняя (sICAM-1) обнаруживается в крови и других биологических жидкостях.
При ряде заболеваний почек показана зависимость между изменениями уровня sICAM-1 в крови и почечной экспрессией ICAM-1 [11]. Определения sICAM-1 у больных СД немногочисленны, их результаты не позволяют сделать однозначный вывод о связи уровня sICAM-1 с выраженностью осложнений СД [12–15]. Экспрессия ICAM-1 в почках больных СД не изучена.
Целью настоящего исследования являлось изучение уровня sICAM-1 в сыворотке крови, а также экспрессии ICAM-1 в почках у больных СД 1 с различными стадиями нефропатии.
Материалы и методы исследования
Обследовали 63 больных СД 1 (22 мужчин и 41 женщину, в возрасте 16–54 лет (29,0±9,7 лет)) при длительности заболевания от 1 мес до 34 лет (8,5±8,0 лет). В исследование не включались больные с кетоацидозом, патологией почек недиабетического генеза, с обострением сопутствующих заболеваний. Исследовали суточную протеинурию, альбуминурию, мочевой осадок, креатинин крови с расчетом скорости клубочковой фильтрации (СКФ) по Cockcroft-Gault, УЗИ почек, проводили cуточное мониторирование артериального давления (СМАД). Нормальная экскреция альбумина с мочой (ЭАМ) выявлена у 23 обследованных (1-я группа), у 29 зафиксирована микроальбуминурия (2-я группа), у 11 – протеинурия (3-я группа). Нарушение клиренса креатинина имело место у 7 больных 3-й группы. Клинико-лабораторная характеристика групп представлена в таблице 1. Пациенты с выраженной нефропатией в целом были несколько старше, имели большую длительность СД, более высокие показатели гликемии натощак, протеинурии, креатинина, меньшую СКФ и более высокие показатели АД по сравнению с больными других групп.
Определение концентрации sICAM-1 в сыворотке крови осуществляли методом иммуноферментного анализа с помощью коммерческих наборов Bendek Medsystems, Biosource (Бельгия) с использованием планшеточного ридера «BioRad 680» (BioRad, США) и программного обеспечения Zemfira. Исследования выполнены в гормональной лаборатории ООО «Лабораторная диагностика» (зав. лабораторией – к.м.н. В.В. Романов). Результаты определения sICAM-1 сравнивали с контрольной группой, в которую вошли 10 здоровых лиц (7 мужчин и 3 женщины) 18–44 лет (средний возраст 28,8±9,5 лет).
Типирование ICAM-1 в биоптатах почек проведено у 15 пациентов (5 мужчин и 10 женщин в возрасте 16–41 года, с длительностью СД от 3 мес до 12 лет (в среднем 4,1±3,3 лет). ЭАМ у трех обследованных находилась в диапазоне нормальных значений, у восьми – в диапазоне микроальбуминурии, у одной пациентки выявлена протеинурия (0,8 г/сут). Все больные имели нормальный клиренс креатинина (СКФ от 62,6 до 130,6 мл/мин). Пункционная нефробиопсия выполнялась для оценки выраженности и характера поражения почек при наличии одного или нескольких факторов риска ДН: большая длительность СД, микроальбуминурия, диабетическая ретинопатия и/или артериальная гипертензия. Процедура выполнялась под контролем УЗИ в отделении ультразвуковой диагностики (зав. – А.В. Сасин) Новосибирской Государственной областной клинической больницы. Пациенты давали письменное информированное согласие на проведение биопсии.
Локализацию ICAM-1 в почечных структурах исследовали методом иммуногистохимического типирования c помощью моноклональных антител к CD54 фирмы «Novocastra Laboratories Ltd» (Великобритания). Выраженность иммуногистохимической реакции оценивали полуколичественно. В качестве контроля использовали почки 15 здоровых лиц (5 мужчин и 5 женщин), 25–42 года (средний возраст 33,2±8,1 лет), погибших от черепно-мозговых травм (автокатастрофы). При судебно-медицинском исследовании этих лиц не выявлено признаков хронических заболеваний, отравлений, травматических и иных повреждений почек. Образцы почек забирали не позднее 24 ч от момента биологической смерти.
Протокол исследования одобрен этическим комитетом Новосибирского государственного медицинского университета (протокол заседания № 1/11 от 16.11. 2005 г.).
Статистическая обработка проведена с использованием пакета STATISTICA 6.0. Использованы дисперсионный анализ, корреляционный анализ, многофакторный регрессионный пошаговый анализ. Для оценки межгрупповых различий применяли t-критерий Стьюдента (для двух независимых групп) или ANOVA (для трех или более групп). Критический уровень значимости при проверке статистических гипотез принимали равным 0,05. Данные представлены как средние и стандартные отклонения (M±SD).
Результаты и их обсуждение
Уровень sICAM-1 в крови у больных СД составил 347,2±126,1 нг/мл (153–812 нг/мл), в контрольной группе – 268,3±63,2 нг/мл (170–375 нг/мл). Средний уровень sICAM-1 у больных оказался значимо выше контроля (р=0,02).
Зависимость уровня sICAM-1 от стадии ДН представлена на рис. 1. У пациентов с нормоальбуминурией содержание sICAM-1 практически не отличалось от такового у здоровых лиц (р=0,20). У больных с микроальбуминурией зафиксирована тенденция к повышению уровня молекулы адгезии, хотя и не достигшая степени статической значимости (р=0,07). Достоверное повышение уровня sICAM-1 имело место у больных с протеинурией (р=0,003).
Больные с артериальной гипертензией имели более высокие значения sICAM-1 по сравнению с остальными пациентами (379,8±129,2 и 319,4±118,2 нг/мл соответственно, р=0,04). Содержание sICAM-1 положительно коррелировало с величиной среднесуточного диастолического АД, с индексом времени диастолической артериальной гипертензии (r=0,39; div=0,04 и r=0,38; div=0,05 соответственно), но не с параметрами систолического АД.
Высокий уровень sICAM-1 был характерен для больных с ранним развитием и быстрым прогрессированием нефро- и ретинопатии (371,4±156,3 нг/мл, n=21, р=0,05). Нормальное содержание молекулы адгезии выявлено при медленном развитии микроангиопатий (263,2±64,1 нг/мл). Наличие сопутствующей патологии не оказало влияния на уровень sICAM-1. Не отмечено различий в уровне sICAM-1 в зависимости от курения и особенностей терапии.
При изучении связи между уровнем sICAM-1 и качеством контроля гликемии наименьшие значения sICAM-1 выявлены у пациентов с хорошо компенсированным СД (303,6±100,7 нг/мл, n=14; отличия от контроля незначительны: р=0,35). Если целевой уровень гликемии не был достигнут, то содержание sICAM-1 значимо превышало контроль (355,8±132,6 нг/мл, n=41, р=0,05). Наибольшие уровни sICAM-1 зафиксированы у больных с лабильным диабетом, резкими колебаниями гликемии в течение суток (387,3±123,3 нг/мл, n=8, р=0,02). Уровень sICAM-1 положительно коррелировал с концентрацией глюкозы натощак (r=0,30; р=0,02) и с постпрандиальной гликемией (r=0,34; р=0,007).
Не установлено значимых корреляций между содержанием sICAM-1 и возрастом больных (r=0,16; р=0,01), длительностью СД (r=–0,09; р=0,01), суточной дозой инсулина (r=0,02; р=0,01), уровнем фибриногена (r=–0,02; р=0,01), лейкоцитов (r=0,20; р=0,01) и СОЭ (r=0,21; р=0,01).
При исследовании биоптатов почек выявлены изменения, свойственные начальным стадиям ДН: гипертрофия и гиперклеточность клубочков, очаговое расширение мезангия, расширение просвета и утолщение стенок капиллярных петель, сужение просвета капсулы, утолщение ее париетального листка, дистрофические и атрофические изменения эпителия канальцев, начальные этапы склероза интерстиция и артериол. У двух больных выявлено полное замещение отдельных клубочков соединительной тканью. Объемная доля мезангия (соотношение мезангий/клубочек) составила в среднем 22,8±4,8% (13,1–30,8%).
Иммуногистохимическое исследование с помощью антител к CD54 не выявило положительной реакции на ICAM-1 в препаратах почек контрольной группы. Присутствие ICAM-1 в клубочках обнаружено у 7 из 15 больных СД. Реакция во всех случаях была слабоположительной, ICAM-1 выявлялись в отдельных клубочках на периферии капиллярных петель. Наиболее часто присутствие ICAM-1 обнаруживалось в канальцах: в 8 случаях реакция оказалась положительной (отчетливое окрашивание всех канальцев в препарате), в 5 случаях – слабоположительной (частичное окрашивание отдельных канальцев) и только в 2 случаях – отрицательной. Пример положительной иммуногистохимической реакции на ICAM-1 в клубочке и канальцах почки у больного с СД представлен на рис. 2.
Большинство пациентов с положительной реакцией на ICAM-1 имели повышенную ЭАМ: у 4 выявлена микроальбуминурия, у 1 – протеинурия; нормоальбуминурия сохранялась у 2 обследованных. Среди пациентов с отрицательной реакцией на ICAM-1 нормальная ЭАМ зафиксирована у 5 пациентов, микроальбуминурия – у 3. Различий по фильтрационной функции почки между выделенными подгруппами не найдено (табл. 2). Больные с положительной реакцией на ICAM-1 в клубочках имели более высокие цифры мониторного систолического и диастолического АД и более высокий индекс времени диастолической гипертензии. Кроме того, у этих больных зарегистрированы более высокие показатели гликемии натощак. Не выявлено какой-либо взаимосвязи между наличием ICAM-1 и возрастом больных, дозой инсулина, длительностью СД. Обращает на себя внимание возможность появления ICAM-1 в клубочках при небольшой длительности заболевания (в наших наблюдениях наименьший срок от манифестации СД составлял 7 мес). Прослеживалась тенденция к увеличению объема мезангия у больных с положительной реакцией на молекулы адгезии в клубочках в сравнении с остальными пациентами (24,4±2,3 и 20,9±6,5 соответственно, р=0,2).
При сопоставлении уровня ICAM-1 в почках с концентрацией растворимой формы этих молекул в сыворотке крови оказалось, что больные с положительной реакцией на ICAM-1 в клубочках имели достоверно более высокий уровень циркулирующих молекул адгезии по сравнению с пациентами с отрицательной иммуногистохимической реакцией (391,6±96,7 и 266,6±63,3, р=0,02).
Положительная реакция на ICAM-1 в канальцах, зафиксированная у большинства обследованных пациентов, наблюдалась при длительности СД от 7 мес до 12 лет. Взаимосвязи выраженности реакции в канальцах с возрастом больных, дозой инсулина, уровнем АД, креатинина, СКФ не установлено. Микроальбуминурия на нижней границе диапазона (31,6 мг/сут) выявлена у одной больной с отрицательной реакцией на ICAM-1. Четыре пациента со слабоположительной реакцией на ICAM-1 имели нормальную ЭАМ, у 1 выявлена микроальбуминурия. Среди пациентов с положительной реакцией нормальная ЭАМ зафиксирована у 2, микроальбуминурия – у 5, протеинурия – у 1. Уровень гликемии натощак у больных с положительной реакцией на ICAM-1 оказался выше, чем у пациентов со слабоположительной реакцией (10,8±2,4 и 7,0±3,3 ммоль/л, р=0,05). Гликемия после еды показала аналогичную тенденцию (9,3±2,9 и 7,3±1,4 ммоль/л, р=0,13).
Прослеживался очевидный параллелизм в изменении уровня ICAM-1 в клубочках и канальцах. У пациентов с отрицательной и слабоположительной реакцией на ICAM-1 в канальцах, за исключением одного случая, ICAM-1 в клубочках не выявлялись. Напротив, у 6 из 8 пациентов с положительной реакцией на ICAM-1 в канальцах данные молекулы выявлялись и в клубочках.
Проведенные исследования свидетельствуют о том, что развитие ДН у больных СД 1 сопровождается заметным увеличением содержания растворимой формы sICAM-1 в сыворотке крови и усилением синтеза ICAM-1 в клубочках и канальцах почек.
Известно, что sICAM-1 появляются в сыворотке в результате протеолиза и слущивания с ICAM-1-позитивных клеток. Ранее в экспериментах показано возрастание синтеза ICAM-1 в эндотелии при воздействии высокого уровня глюкозы [16, 17]. В работе М.В. Шестаковой и соавт. [18] показано увеличение экспрессии ICAM-1 на гранулоцитах и лимфоцитах крови у больных СД 1, имеющих протеинурию. Можно предположить, что источниками избыточного количества циркулирующих sICAM-1 у больных СД являются клетки сосудистого эндотелия и лейкоциты крови.
Основным фактором, определяющим повышение sICAM-1 в кровотоке, оказался уровень гликемии. Ранее было показано, что улучшение контроля гликемии на фоне интенсивной инсулинотерапии у больных СД 2 сопровождается снижением концентрации молекул адгезии в крови [19]. Это согласуется с результатами экспериментальных работ, показавших стимулирующее влияние супрафизиологических количеств глюкозы на экспрессию молекул адгезии в эндотелиальных клетках [16, 17]. Установлено, что воздействие избытка глюкозы приводит к увеличению адгезии моноцитов [20] и нейтрофилов [16] к сосудистому эндотелию. Аналогичные эффекты оказывают гликированные белки [21]. Таким образом, стимулируя синтез молекул адгезии, гипергликемия запускает развитие иммуновоспалительных реакций в сосудах.
В связи с этим увеличение содержания sICAM-1 в сыворотке крови можно использовать как маркер интенсивности хронического сосудистого воспаления у больных СД. Клиническая значимость данного параметра определяется его тесной взаимосвязью с прогрессированием микроангиопатий. Поскольку содержание sICAM-1 не коррелирует с «традиционными» маркерами воспаления (такими, как количество лейкоцитов, СОЭ и фибриноген), его исследование имеет самостоятельное диагностическое значение.
Как показали наши исследования, уровень sICAM-1 в кровотоке у больных СД 1 в определенной степени отражает активацию синтеза ICAM-1 в клубочках почек. Повышение количества ICAM-1 в почках, однако, обнаруживалось на более ранних стадиях ДН, чем изменения уровня sICAM-1 в кровотоке. Появление ICAM-1 в клубочках и канальцах почек в количествах, выявляемых иммуногистохимическим методом, зафиксировано нами на стадии микроальбуминурии, а в некоторых случаях даже при нормальной экскреции альбумина с мочой.
Полученные данные согласуются с результатами работ, в которых обнаружена повышенная экспрессия ICAM-1 в клубочках [22–25] и тубулоинтерстиции почек [23, 25] на различных экспериментальных моделях СД. Известно, что активация синтеза молекул адгезии является характерным признаком целого ряда заболеваний почек, в патогенезе которых присутствует воспалительный компонент. Повышенная экспрессия ICAM-1 в клубочках обнаружена при различных морфологических формах гломерулонефритов [26, 27], васкулите Шенляйн–Геноха, люпус-нефрите [26]. Активацию экспрессии ICAM-1 в канальцах и/или интерстиции наблюдали при гломерулонефритах [26], тубулоинтерстициальных нефритах [28], реакции отторжения почечного трансплантата [29]. Таким образом, активация синтеза адгезивных белков носит характер универсальной реакции. Вместе с тем факторы, запускающие эту реакцию при разных формах патологии различны.
Проведенные исследования показали взаимосвязь между качеством контроля гликемии и уровнем ICAM-1 в почках. Это соответствует представлениям о том, что гипергликемия является наиболее вероятной пусковой причиной активации синтеза ICAM-1 в почках при СД. In vitro показано, что высокий уровень глюкозы приводит к возрастанию продукции ICAM-1 в эндотелиальных [30] и мезангиальных [31] клетках. Помимо гипергликемии, синтез ICAM-1 в мезангиоцитах может запускать оксидативный стресс [32], а также механическое напряжение в условиях внутриклубочковой гипертензии [33].
Гиперэкспрессия молекул адгезии является необходимым условием привлечения в почки мононуклеарных провоспалительных клеток. Возрастание их числа в клубочках и интерстиции почек при СД зафиксировано в ряде клинических [3, 5] и экспериментальных [6, 22, 23, 25, 34, 35] работ. Инфильтрирующие почки провоспалительные клетки могут включаться в патогенез нефросклероза, вырабатывая фиброгенные факторы роста и цитокины. При СД макрофагальная инфильтрация почечных клубочков и канальцев взаимосвязана с выраженностью фиброза, альбуминурией, артериальной гипертензией и функцией почек [3, 6]. В эксперименте показано, что нейтрализующие ICAM-1 антитела препятствуют развитию макрофагальной инфильтрации клубочков при СД [7]. Мыши линии db/db (модель СД 2) с «нокаутированным» геном ICAM-1, в значительной степени «защищены» от развития нефропатии: у них медленнее нарастает альбуминурия, менее выражены лейкоцитарная инфильтрация и морфологические изменения в клубочках и канальцах, медленнее развивается нефросклероз [8]. Это позволяет рассматривать ICAM-1 в качестве одного из медиаторов, участвующих в развитии ДН.
Дальнейшее изучение роли воспалительных реакций в формировании ДН представляется перспективным для изучения механизмов развития и разработки новых подходов к лечению этого грозного осложнения.
Выводы
1. Выявлена активация синтеза молекул межклеточной адгезии ICAM-1 в почках и повышение растворимой формы данных молекул (sICAM-1) в сыворотке крови у больных СД 1.
2. Активация синтеза ICAM-1 в канальцах и клубочках почек наблюдается на самых ранних стадиях нефропатии, даже при небольшой длительности СД.
3. Повышение уровня sICAM-1 в крови наиболее характерно для клинически выраженных стадий ДН.
4. Активация синтеза ICAM-1 при СД связана с качеством гликемического контроля, степенью альбуминурии, наличием артериальной гипертензии.
5. Изменения экспрессии ICAM-1 в почках и их уровня в кровотоке подтверждают роль воспалительных реакций в формировании ДН.
1. Сунцов Ю.И., Дедов И.И. Государственный регистр больных сахарным диабетом — основная информационная система для расчета экономических затрат государства на сахарный диабет и их прогнозирование. // Сахарный диабет 2005; 2: 2-5.
2. Дедов И.И., Шестакова М.В. Диабетическая нефропатия. М.: Универсум Паблишинг; 2000.
3. Furuta T., Saito T., Ootaka T. et al. The role of macrophages in diabetic glomerulosclerosis. // Am J Kidney Dis 1993; 21 (5): 480-485.
4. Mezzano S., Droguett A., Burgos M.E. et al. Renin-angiotensin system activation and interstitial inflammation in human diabetic nephropathy. // Kidney Int Suppl 2003; 86: S64-S70.
5. Mezzano S., Aros C., Droguett A. et al. NF-kappaB activation and overexpression of regulated genes in human diabetic nephropathy. // Nephrol Dial Transplant 2004; 19 (10): 2505-2512.
6. Chow F., Ozols E., Nikolic-Paterson D.J. et al. Macrophages in mouse type 2 diabetic nephropathy: correlation with diabetic state and progressive renal injury. // Kidney Int 2004; 65 (1): 116-128.
7. Sugimoto H., Shikata K., Hirata K. et al. Increased expression of intercellular adhesion molecule-1 (ICAM-1) in diabetic rat glomeruli: glomerular hyperfiltration is a potential mechanism of ICAM-1 upregulation. // Diabetes 1997; 46: 2075-2081.
8. Chow F.Y., Nikolic-Paterson D.J., Ozols E. et al. Intercellular adhesion molecule-1 deficiency is protective against nephropathy in type 2 diabetic db/db mice. // J Am Soc Nephrol 2005; 16 (6): 1711-1722.
9. Chow F.Y., Nikolic-Paterson D.J., Ozols E. et al. Monocyte chemoattractant protein-1 promotes the development of diabetic renal injury in streptozotocin- treated mice. // Kidney Int 2006; 69 (1): 73-80.
10. Пальцев М.А., Иванов А.А., Северин С.Е. Межклеточные взаимодействия. М: Медицина; 2003 .
11. Lhotta K., Schlogl A., Kronenberg F. et al. Soluble intercellular adhesion molecule-1 (ICAM-1) in serum and urine: correlation with renal expression of ICAM-1 in patients with kidney disease. // Clin Nephrol 1997; 48 (2): 85-91.
12. Fasching P., Veitl M., Rohac M. et al. Elevated concentrations of circulating adhesion molecules and their association with microvascular complications in insulin-dependent diabetes mellitus. // J Clin Endocrinol Metab 1996; 81 (12): 4313-4317.
13. Guler S., Cakir B., Demirbas B. et al. Plasma soluble intercellular adhesion molecule 1 levels are increased in type 2 diabetic patients with nephropathy. // Horm Res 2002; 58 (2): 67-70.
14. Boulbou M.S., Koukoulis G.N., Makri E.D. et al. Circulating adhesion molecules levels in type 2 diabetes mellitus and hypertension. // Int J Cardiol 2005; 98 (1): 39-44.
15. Nelson C.L., Karschimkus C.S., Dragicevic G. et al. Systemic and vascular inflammation is elevated in early IgA and type 1 diabetic nephropathies and relates to vascular disease risk factors and renal function. // Nephrol Dial Transplant 2005 20 (11): 2420-2426.
16. Omi H., Okayama N., Shimizu M. et al. Participation of high glucose concentrations in neutrophil adhesion and surface expression of adhesion molecules on cultured human endothelial cells: effect of antidiabetic medicines. // J Diabetes Complications 2002; 16 (3): 201-208.
17. Altannavch T.S., Roubalova K., Kucera P., Andel M. Effect of high glucose concentrations on expression of ELAM-1, VCAM-1 and ICAM-1 in HUVEC with and without cytokine activation. // Physiol Res 2004; 53 (1): 77-82.
18. Шестакова М.В., Кочемасова Т.В., Горелышева В.А. и др. Роль молекул адгезии (ICAM-1 и Е-селектина) в развитии диабетических микроангиопатий. // Тер архив 2002; 6: 24-27.
19. Albertini J.P., Valensi P., Lormeau B. et al. Elevated concentrations of soluble E-selectin and vascular cell adhesion molecule-1 in NIDDM. Effect of intensive insulin treatment. // Diabetes Care 1998; 21 (6): 1008-1013.
20. Manduteanu I., Voinea M., Serban G., Simionescu M. High glucose induces enhanced monocyte adhesion to valvular endothelial cells via a mechanism involving ICAM-1, VCAM-1 and CD18. // Endothelium 1999; 6 (4): 315-324.
21. Desfaits A.C., Serri O., Renier G. Gliclazide reduces the induction of human monocyte adhesion to endothelial cells by glycated albumin. // Diabetes Obes Metab 1999; 1 (2): 113-120.
22. Sassy-Prigent C., Heudes D., Mandet C. Early glomerular macrophage recruitment in streptozotocin-induced diabetic rats. // Diabetes 2000; 49 (3): 466-475.
23. Usui H., Shikata K., Matsuda M. et al. HMG-CoA reductase inhibitor ameliorates diabetic nephropathy by its pleiotropic effects in rats. // Nephrol Dial Transplant 2003; 18: 265-272.
24. Lee F.T., Cao Z., Long D.M. et al. Interactions between angiotensin II and NF-kappaB-dependent pathways in modulating macrophage infiltration in experimental diabetic nephropathy. // J Am Soc Nephrol 2004; 15 (8): 2139-2151.
25. Qi X.M., Wu G.Z., Wu Y.G. et al. Renoprotective Effect of Breviscapine through Suppression of Renal Macrophage Recruitment in Streptozotocin-Induced Diabetic Rats. // Nephron Exp Nephrol 2006; 104 (4): e147-e157.
26. Lhotta K., Neumayer H.P., Joannidis M. et al. Renal expression of intercellular adhesion molecule-1 in different forms of glomerulonephritis. // Clin Sci (Lond) 1991; 81 (4): 477-481.
27. Nguyen T.T., Shou I., Funabiki K. et al. Correlations among expression of glomerular intercellular adhesion molecule 1 (ICAM-1), levels of serum soluble ICAM-1, and renal histopathology in patients with IgA nephropathy. // Am J Nephrol 1999; 19 (4): 495-499.
28. Shappell S.B., Mendoza L.H., Gurpinar T. et al. Expression of adhesion molecules in kidney with experimental chronic obstructive uropathy: the pathogenic role of ICAM-1 and VCAM-1. // Nephron 2000; 85 (2): 156-166.
29. Andersen C.B., Blaehr H., Ladefoged S., Larsen S. Expression of the intercellular adhesion molecule-1 (ICAM-1) in human renal allografts and cultured human tubular cells. // Nephrol Dial Transplant 1992; 7 (2): 147-154.
30. Baumgartner-Parzer S.M., Wagner L., Pettermann M. et al. Modulation by high glucose of adhesion molecule expression in cultured endothelial cells. // Diabetologia 1995; 38 (11): 1367-1370.
31. Park C.W., Kim J.H., Lee J.H. et al. High glucose-induced intercellular adhesion molecule-1 (ICAM-1) expression through an osmotic effect in rat mesangial cells is PKC-NF-kappa B-dependent. // Diabetologia 2000; 43 (12): 1544-1553.
32. Satriano J.A., Banas B., Luckow B. et al. Regulation of RANTES and ICAM-1 expression in murine mesangial cells. // J Am Soc Nephrol 1997; 8 (4): 596-603.
33. Riser B.L., Varani J., Cortes P. et al. Cyclic stretching of mesangial cells up-regulates intercellular adhesion molecule-1 and leukocyte adherence: a possible new mechanism for glomerulosclerosis. // Am J Pathol 2001; 158 (1): 11-17.
34. Tone A., Shikata K., Sasaki M. et al. Erythromycin ameliorates renal injury via anti-inflammatory effects in experimental diabetic rats. // Diabetologia 2005; 48 (11): 2402-2411.
35. Biswas S.K., Lopes de Faria J.B. Hypertension induces oxidative stress but not macrophage infiltration in the kidney in the early stage of experimental diabetes mellitus. // Am J Nephrol 2006; 26 (5): 415-422.
Алгебраические выражения — объяснения и примеры
Алгебра — интересный и увлекательный раздел математики, в котором числа, фигуры и буквы используются для выражения задач. Независимо от того, изучаете ли вы алгебру в школе или сдаете какой-то тест, вы заметите, что почти все математические задачи представлены словами.
Следовательно, необходимость переводить письменные текстовые задачи в алгебраические выражения возникает тогда, когда нам нужно их решить.
Большинство алгебраических задач со словами состоят из рассказов или примеров из реальной жизни.Другие — простые фразы, такие как описание математической задачи. Из этой статьи вы узнаете, как написать алгебраических выражений из простых задач со словами, а затем перейти к слегка сложным задачам со словами.
Что такое алгебраическое выражение?
Многие люди попеременно используют алгебраические выражения и алгебраические уравнения, не подозревая, что это совершенно разные термины.
Алгебраика — это математическая фраза, в которой две стороны фразы соединены знаком равенства (=).Например, 3x + 5 = 20 — это алгебраическое уравнение, где 20 представляет собой правую часть (RHS), а 3x +5 представляет собой левую часть (LHS) уравнения.
С другой стороны, алгебраическое выражение — это математическая фраза, в которой переменные и константы объединяются с помощью операционных символов (+, -, × & ÷). В алгебраическом символе отсутствует знак равенства (=). Например, 10x + 63 и 5x — 3 являются примерами алгебраических выражений.
Давайте рассмотрим терминологию, используемую в алгебраических выражениях:
- Переменная — это буква, значение которой нам неизвестно.Например, x — это наша переменная в выражении: 10x + 63.
- Коэффициент — это числовое значение, используемое вместе с переменной. Например, 10 — это переменная в выражении 10x + 63.
- Константа — это терм, который имеет определенное значение. В этом случае 63 — это константа в алгебраическом выражении, 10x + 63.
Существует несколько типов алгебраических выражений, но основной тип включает:
- Мономиальное алгебраическое выражение
Этот тип выражения имеет только один член, например 2x, 5x 2 , 3xy и т. д.
Алгебраическое выражение, содержащее два, в отличие от термов, например, 5y + 8, y + 5, 6y 3 + 4 и т. Д.
Это алгебраическое выражение с более чем одним членом и с ненулевыми показателями степени переменные. Пример полиномиального выражения: ab + bc + ca и т. Д.
Другие типы алгебраических выражений:
Числовое выражение состоит только из чисел и операторов. В числовое выражение не добавляется никакая переменная. Примеры числовых выражений: 2 + 4, 5-1, 400 + 600 и т. Д.
Это выражение содержит переменные вместе с числами, например, 6x + y, 7xy + 6 и т. Д.
Как решить алгебраическое выражение?
Цель решения алгебраического выражения в уравнении — найти неизвестную переменную. Когда два выражения приравниваются, они образуют уравнение, и поэтому становится легче найти неизвестные члены.
Чтобы решить уравнение, поместите переменные с одной стороны, а константы — с другой. Вы можете изолировать переменные, применяя арифметические операции, такие как сложение, вычитание, умножение, деление, квадратный корень, кубический корень и т. Д.
Алгебраические выражения всегда взаимозаменяемы. Это означает, что вы можете переписать уравнение, поменяв местами LHS и RHS.
Пример 1
Вычислите значение x в следующем уравнении
5x + 10 = 50
Решение
Заданное уравнение как 5x + 10 = 50
- Изолируйте переменные и константы;
- Вы можете сохранить переменную на левой стороне, а константы на правой.
5x = 50-10
5x = 40
Разделите обе части на коэффициент переменной;
x = 40/5 = 8
Следовательно, значение x равно 8.
Пример 2
Найдите значение y, когда 5y + 45 = 100
Решение
Изолировать переменные от констант;
5y = 100 -45
5y = 55
Разделим обе части на коэффициент;
y = 55/5
y = 11
Пример 3
Определите значение переменной в следующем уравнении:
2x + 40 = 30
Решение
Разделите переменные из константы;
2x = 30-40
2x = -10
Разделите обе стороны на 2;
x = -5
Пример 4
Найдите t, когда 6t + 5 = 3
Решение
Отделите константы от переменной,
6t = 5 — 3
6t = -2
Разделим обе части на коэффициент,
t = -2/6
Упростим дробь,
t = -1/3
Практические вопросы
1.Если x = 4 и y = 2, найдите следующие выражения:
a. 2г + 4
б. 10х + 40л;
г. 15л — 5х
д. 5x + 7
e. 11y + 6
ф. 6x — 2
г. 8лет — 5
ч. 60 — 5x — 2y
2. Сэм кормит свою рыбу одинаковым количеством корма (пусть равным x ) трижды в день. Сколько еды он накормит рыбу в неделю?
3. Нина испекла по 3 кекса для сестры и по 2 кекса для каждой подруги (пусть равно x ).Сколько всего кексов она испекла?
4. У Джонса на ферме 12 коров. Большинство коров дают 30 литров молока в день (пусть равно x ). Сколько коров не дают 30 литров молока в день?
Предыдущий урок | Главная страница | Следующий урокЧасти выражения
Алгебраические выражения — это комбинации переменные , числа и хотя бы одну арифметическую операцию.
Например, 2 Икс + 4 у — 9 является алгебраическим выражением.
Срок: Каждое выражение состоит из терминов. Термин может быть числом со знаком, переменной или константой, умноженной на переменную или переменные.
Фактор: То, что умножается на другое. Фактор может быть числом, переменной, термином или более длинным выражением. Например, выражение 7 Икс ( у + 3 ) имеет три фактора: 7 , Икс , а также ( у + 3 ) .
Коэффициент: Числовой коэффициент выражения умножения, содержащего переменную. Рассмотрим выражение на рисунке выше, 2 Икс + 4 у — 9 . В первом семестре 2 Икс , коэффициент равен 2 : во втором семестре, 4 у , коэффициент равен 4 .
Постоянный: Число, значение которого не может быть изменено.В выражении 2 Икс + 4 у — 9 , срок 9 является константой.
Как условия: Термины, содержащие одинаковые переменные, такие как 2 м , 6 м или 3 Икс у а также 7 Икс у . Если в выражении есть несколько постоянных членов, они также похожи на термины.
|
|
|
|
|
|
|
|
|
|
Пример:
Определите термины, такие как термины, коэффициенты и константы в выражении.
9 м — 5 п + 2 + м — 7
Во-первых, мы можем переписать вычитания как добавления.
9 м — 5 п + 2 + м — 7 знак равно 9 м + ( — 5 п ) + 2 + м + ( — 7 )
Так что термины находятся 9 м , ( — 5 п ) , м , 2 , а также ( — 7 ) .
Как условия — это термины, содержащие одинаковые переменные.
9 м а также 9 м пара как условия . Постоянные условия 2 а также — 7 также похожи на термины.
Коэффициенты — числовые части термина, содержащего переменную.
Итак, вот коэффициенты находятся 9 , ( — 5 ) , а также 1 . ( 1 коэффициент при члене м .)
В постоянный термины — это термины без переменных, в данном случае 2 а также — 7 .
Алгебраические выражения должны быть написаны и интерпретированы осторожно.Алгебраическое выражение 5 ( Икс + 9 ) является нет эквивалентно алгебраическому выражению, 5 Икс + 9 .
Посмотрите разницу между двумя выражениями в таблице ниже.
| Словесные фразы | Алгебраическое выражение |
| В пять раз больше числа и девяти | 5 ( Икс + 9 ) |
| Девять больше, чем в пять раз больше | 5 Икс + 9 |
При написании выражений для неизвестных величин мы часто используем стандартные формулы.Например, алгебраическое выражение «расстояние, если скорость 50 миль в час, а время Т часов «это D знак равно 50 Т (по формуле D знак равно р Т ).
Выражение вроде Икс п называется властью. Здесь Икс это база, а п — показатель степени. Показатель степени — это количество раз, когда основание используется в качестве фактора.Словосочетание для этого выражения: » Икс к п th власть.»
Вот несколько примеров использования экспонент.
| Словесные фразы | Алгебраическое выражение |
| Семь раз м в четвертой степени | 7 м 4 |
| Сумма Икс в квадрате и 12 времена у | Икс 2 + 12 у |
| Икс раз в кубе у в шестой степени | Икс 3 ⋅ у 6 |
Алгебраическое выражение: примеры и выражения
Алгебра — интригующий и восхитительный раздел математики, в котором числа, фигуры и буквы используются для выражения задач.Независимо от того, изучаете ли вы алгебру в колледже или готовитесь к экзамену, вы наверняка увидите, что почти все математические вопросы выражаются в алгебраическом выражении.
Следовательно, потребность в преобразовании составных словесных проблем в алгебраические выражения возникает тогда, когда нам нужно их решать.
Многие задачи с алгебраическим выражением состоят из рассказов или случаев из реальной жизни. Другие — это базовые выражения, например, краткое изложение математической задачи. Этот пост действительно узнает, как составлять алгебраические выражения из простых словесных задач, после этого прорыва к легким сложным задачкам со словами.
Подробнее об алгебраических выраженияхМногие люди взаимно используют алгебраические выражения и алгебраические уравнения, не зная, что эти термины различны.
Алгебраическое выражение — это математическая фраза, в которой знак равенства соединяет две стороны фразы (= -RRB-. Например, 3x + 5 = 20 — это алгебраическое уравнение, где 20 обозначает правую часть (RHS), а также 3x +5 представляет собой левую часть формулы (LHS).
Напротив, алгебраическое выражение — это математическое выражение, в которое переменные и константы включаются с использованием операционных символов (+, -, × & ÷).В алгебраическом символе отсутствует знак равенства (= -RRB-. Например, 10x + 63, а также 5x– 3 являются экземплярами алгебраических выражений.
Давайте оценим терминологию, используемую в алгебраическом выражении:
Переменная — это буква, значение которой нам неизвестно. Например, x — это наша переменная в выражении: 10x + 63.
Коэффициент — это числовая ценность, используемая вместе с переменной. Например, 10 — это переменная в выражении 10x + 63.
Согласованный — это термин, имеющий гарантированное значение. В этой ситуации 63 — это константа в алгебраическом выражении, 10x + 63.
Есть несколько типов алгебраических выражений; однако знаменательный вид состоит из:
Мономиальное алгебраическое выражениеЭтот тип выражения состоит только из одного члена, например, 2x, 5x 2,3 xy и так далее.
Биномиальное выражениеАлгебраическое выражение, имеющее два, в отличие от членов, например, 5y + 8, y +5, 6y3 + 4 и т. Д.
Полиномиальное выражениеЭто алгебраическое выражение, содержащее более одного члена и неэкспоненты переменных.Примером полиномиального выражения является мышца живота + b c + ca и так далее.
Другие виды алгебраических выражений:
Числовое выражение:Математическое выражение включает числа, а также драйверы. В числовое выражение никакая переменная не добавляется. Примеры числовых выражений: 2 +4, 5-1, 400 +600 и т. Д.
Выражение переменной:Это выражение, которое включает переменные наряду с числами, например 6x + y, 7xy +6 и т. Д.
Решение алгебраических выраженийФункция решения алгебраического выражения в уравнении заключается в обнаружении неизвестной переменной. Когда два выражения связаны, они образуют уравнение, и, следовательно, становится менее сложным обращаться к незнакомым терминам.
Читайте также: Что такое агенты социализации?
Чтобы исправить формулу, но переменные с одной стороны и константы с другой стороны. Переменные могут быть разделены с помощью арифметических операций, таких как улучшение, сокращение, воспроизведение, отдел, квадратный корень, начало координат и т. Д.
Алгебраическое выражение всегда совместимо. Это означает, что вы можете переформулировать формулу, поменяв местами LHS и RHS.
Пример 1Определите значение x по следующей формуле
5x + 10 = 50
Решение
Предоставляется формула как 5x + 10 = 50
Изолируйте переменные и константы;
Вы можете поддерживать переменную в LHS, а также константы в RHS.
5x = 50-10
Вычтите константы;
5x = 40
Разделите обе стороны коэффициентом переменной;
х = 40/5 = 8
По этой причине значение x равно 8.
Пример 2Определите значение y, когда 5y + 45 = 100
Решение
Отделить переменные от констант;
5лет = 100-45
5лет = 55
Разделить обе стороны коэффициентом;
г = 55/5
г = 11
10 английских выражений и их значений
Изучая новый язык, мы обычно в первую очередь концентрируемся на изучении основных моментов — грамматики и словарного запаса.Хотя это действительно важно — и это то, что мы обсуждаем на наших онлайн-курсах английского языка, — также важно для изучения идиом, выражений и распространенных фраз при изучении языков.
Это поможет вам выйти за рамки грамматически правильных, так что вы больше походите на носителя языка и получите представление о культуре людей, говорящих на этом языке. При изучении любого нового языка важны фразы и выражения, поэтому давайте взглянем на наиболее часто используемых английских выражений :
Вот 10 английских выражений, которые необходимо знать и их значения
1.Лучшее после нарезанного хлеба
Честно говоря, это выражение не обязательно используется очень часто, но оно имеет большее влияние, когда используется. Этот «титул» следует присваивать только великим идеям или нововведениям, желательно более гениальным, чем изобретение предварительно нарезанного хлеба. Я, например, не совсем уверен, что такого хорошего в нарезанном хлебе. Нарезать хлеб не так уж и сложно, а неразрезанный буханка остается свежей еще день или два …
2. Вызов принят
Это выражение приобрело популярность благодаря постоянному использованию Нила Патрика Харриса в роли Барни Стинсона в телешоу «Как я встретил вашу маму».Его можно использовать по-разному, но он наиболее уместен, когда кто-то утверждает, что что-то невозможно или не следует делать по какой-либо причине. Пример: «Ты никогда не сможешь съесть весь этот торт!» — «Вызов принят!»
3. Переходите этот мост, когда подойдете к нему
Это полезное выражение очень любят прокрастинаторы, потому что это удобный способ отложить или избежать столкновения с проблемой! Если кто-то укажет на потенциальную проблему в будущем, вы можете просто отклонить его, сказав: «Мы перейдем этот мост, когда подойдем к нему»; буквально означает «Я не хочу сейчас об этом думать.Будем надеяться, что этого не произойдет, но если это произойдет, тогда мы решим проблему ».
4. Не кладите все яйца в одну корзину
Есть друг, который думает, что он начинающий писатель, и думает о том, чтобы бросить свою повседневную работу, чтобы писать полный рабочий день, чтобы закончить «роман века»? Вы можете посоветовать ему не «класть все яйца в одну корзину». Большинство людей, вероятно, хранят яйца в холодильнике… но это выражение используется, когда мы хотим предостеречь других, чтобы они не полагались только на одну возможность — оставлять свои варианты открытыми.Однако если вы писатель и ваши друзья говорят вам это, вам следует либо отказаться от этого романа, либо найти себе новых друзей!
5. Голландское мужество
«Голландское мужество» — это скорее эвфемизм пьяной храбрости. Если вы столкнулись с неудобной задачей, которую, по вашему мнению, можно решить только с «голландской храбростью», это означает, что вы предпочли бы быть пьяным (или хотя бы немного навеселе), выполняя указанную задачу. Это английское выражение особенно актуально для изучения языка , поскольку даже ученые уже признали, что несколько стаканов выпивки могут помочь вам свободно говорить на любом языке, который вы выучили.Но будьте осторожны, не переусердствуйте; полная потеря речи связана с сильным пьянством!
тысяч общеупотребительных выражений на английском языке • 7ESL
Выражений на английском! список тысяч английских выражений, часто используемых в повседневном общении. Изучение этих распространенных выражений поможет вашему английскому звучать более плавно и естественно, как у носителя языка.
Английские выражения
Что такое английские выражения?
английских выражений, также широко известных как выражения, — это слова или группы слов, которые при определенном использовании передают определенное значение.Выражения бывают разных форм, например, некоторые из них — это словосочетания, другие — общие фразы, а другие — идиомы или даже фразовые глаголы. Вот более подробный взгляд на каждый из этих видов выражений.
Совместное использование
Словосочетания — это выражения, состоящие из двух или более слов, которые соответствуют определенной манере изложения вещей. Как правило, слова имеют большее значение при использовании вместе, чем при использовании по отдельности.
Например, « компьютерный монитор ».Словосочетания также являются противопоставлениями, например, « мертвый серьезный ».
Штифт
Общие фразы
В английском языке много фраз, и они довольно часто используются большинством англоговорящих и поэтому понятны большинству англоговорящих.
Например:
- Давно не видел
- Хорошая
- Что случилось
Фразовые глаголы
Согласно своему названию, фразовые глаголы состоят из глагола и частицы.Частица — это предлог или наречие. Эта комбинация приводит к созданию нового слова или фразы.
Например:
- Мне надоело (скучать) своей работой.
- извините; Я м не с (согласен) с вами по этому поводу.
Штифт
Какие английские поговорки распространены?
Английская поговорка — распространенное выражение, обычно передающее определенную мудрость и совет.Эти высказывания включают пословицы и даже идиомы. Высказывания также могут относиться к выражениям, относящимся к определенному человеку. Например, высказывания Соломона о молодежи.
Идиомы
Идиома — это набор из двух или более слов, значение которых превышает буквальное значение отдельных слов. Идиомы также включают разговорные выражения.
Например: «когда он услышал новости, он пошел на бананы ».
Обычно, когда дело доходит до английского языка, идиомы отличаются от пословиц в том, что касается высказываний, основанных на том факте, что они будут короче.
В то время как пословицы представляют собой законченные предложения, идиомы обычно представляют собой короткие неполные утверждения. Примеры идиом, из которых состоят изречения, включают следующее: на мгновение окунуться в кусты.
Как правило, идиомы необходимо сочетать с другими словами, чтобы правильно передать их значение.
Штифт
Притчи
Слово пословицы часто считается синонимом слова изречения. Обычно они являются частью каждой культуры, и у них очень жесткая структура, в результате чего пропуск одного слова может полностью изменить их значение.
Некоторые распространенные английские пословицы включают: действия говорят громче, чем слова, каждое облако имеет серебряную подкладку, картинка стоит тысячи слов, лучше поздно, чем никогда, и так далее.
Важность изучения общих выражений и поговорок
Выучить английские выражения и распространенные изречения очень важно для любого говорящего по-английски. Во-первых, эти утверждения упрощают обсуждение некоторых моментов без использования множества других слов.
Кроме того, использование выражений и поговорок — это то, что вы найдете в любом языке.Следовательно, нельзя сказать, что вы достаточно владеете каким-либо языком, не овладев некоторыми из этих выражений и высказываний.
Кроме того, важно изучать английские выражения и поговорки, потому что это означает, что вы можете понимать, что говорят люди, не понимая их неправильно. Выражения и поговорки часто используют слова, которые передают определенное значение, выходящее за рамки самих слов.
Например, « пнуть ведро » буквально не ударяет ногой по ведру, а скорее означает, что кто-то прошел мимо.Поэтому, не изучая английские выражения и распространенные поговорки, вы постоянно неправильно понимаете то, что говорят многие опытные ораторы.
Кроме того, изучение выражений и поговорок может помочь вам улучшить свои навыки критического мышления, поскольку вы получите некоторую мудрость из того, что говорят некоторые из этих выражений. Например, поговорка: « Я никогда не учился у человека, который со мной соглашался, » поможет вам понять, что конструктивное отношение к критике — хороший способ улучшить ваше обучение, выявляя свои ошибки и работая над ними.
Тысячи общеупотребительных английских выражений
Когда вы слушаете английский язык, вы обязательно слышите много выражений, которые могут не иметь смысла с первого взгляда. Но это нормально, так как у нас есть подборка отличных статей, которые точно покажут вам, к чему прислушиваться.
Короче говоря, выражение — это слово или фраза, которые выражают мысль, идею или чувство. В английском языке выражения используются очень часто, и некоторые из них могут показаться бессмысленными или бессмысленными.Например, не кладите все яйца в одну корзину. Вы можете подумать, что говорящий говорит о яйцах, но на самом деле они пытаются сказать, что не следует полагаться исключительно на что-то одно. Это показывает нам, что когда вы слышите выражение, используемое в разговоре на английском языке, это не всегда буквальная фраза.
Поскольку этот тип речи очень часто используется в повседневной беседе, а также в письменном тексте, для вас важно хорошо знать некоторые из наиболее распространенных выражений, которые могут использовать носители английского языка.По мере того, как вы продолжите свое путешествие по изучению английского языка, вы, несомненно, научитесь все больше и больше выражений. Просмотрите некоторые из этих распространенных и найдите время, чтобы попытаться понять, что, по вашему мнению, это означает, прежде чем смотреть определение. Вы можете удивиться, когда обнаружите, что все не всегда так, как кажется.
Словосочетания на английском языке.
Общие фразы на английском языке.
английских фразовых глаголов.
Английские идиомы.
Видео с английскими выражениями
70+ видеоуроков по распространенным выражениям, часто используемым в повседневной беседе на английском языке.
Формальные и неформальные выражения в английском языке.
Множественные средства действия и выражения
Обеспечение множественных средств действия и выражения (MMAE) — это принцип универсального UDL, который получил название «как» обучения (Rose & Meyer, 2002).Национальный центр UDL описывает множественные средства действия и выражения следующим образом:
Учащиеся различаются способами, которыми они могут ориентироваться в учебной среде и выражать то, что они знают. Например, люди со значительными двигательными нарушениями (например, церебральный паралич), те, кто борется со стратегическими и организационными способностями (расстройства управляющих функций), те, у кого есть языковой барьер, и т. Д. Очень по-разному подходят к учебным задачам. Некоторые могут хорошо выражать свои мысли в письменном виде, но не в речи, и наоборот.Также следует признать, что действия и выражения требуют большого количества стратегии, практики и организации, и это еще одна область, в которой учащиеся могут отличаться. На самом деле не существует одного средства действия и выражения, которое было бы оптимальным для всех учащихся; предоставление вариантов действий и самовыражения имеет важное значение. (Национальный центр UDL, Принцип II, 2011 г.)
Центр предоставляет расширенные Руководства с примерами под каждой контрольной точкой, чтобы лучше понять способы обеспечения множественных средств действия и выражения.Ниже приведены некоторые конкретные примеры множественных средств действия или выражения, которые может использовать преподаватель послешкольного образования.
Некоторые способы обеспечения множественных средств действия и самовыражения в послесредних классах
- Предоставить варианты ответа учащегося
- Разрешить использование нескольких форматов для демонстрации знаний — устной или письменной
- Используйте различные форматы оценки
- Установить основу для планирования с помощью подробных описаний заданий
- Поощрять использование технологий, чтобы учащиеся точно выражали свое понимание
Вернуться к ресурсам UDL
примеров выражений SQL
примеров выражений SQLПервые шесть примеров в этом разделе относятся к сетке. который содержит три столбца из таблицы сотрудников: emp_id , dept_id , и зарплата .
Пример 1
Выражение <50000 в
Строка критериев в столбце зарплата столбца
grid извлекает информацию о сотрудниках, чья зарплата меньше
чем 50 000 долларов.
Оператор SELECT , созданный InfoMaker это:
ВЫБЕРИТЕ employee.emp_id, employee.dept_id, сотрудник. зарплата ОТ сотрудника ГДЕ employee.salary <50000
Пример 2
Выражение 100 в
Строка критериев в столбце DeptId столбца
grid извлекает информацию о сотрудниках, принадлежащих к отделу
100.
Оператор SELECT , созданный InfoMaker это:
ВЫБЕРИТЕ employee.emp_id, employee.dept_id, сотрудник. зарплата ОТ сотрудника ГДЕ employee.dept_id = '100'
Пример 3
Выражение > 300 в
строка Criteria в столбце EmpId и выражение <50000 в
Строка критериев в столбце Зарплата столбца
grid получить информацию о любом сотруднике, чей идентификатор
более 300 и с зарплатой менее 50 000 долларов.
Оператор SELECT , созданный InfoMaker это:
ВЫБЕРИТЕ employee.emp_id, employee.dept_id, сотрудник. зарплата ОТ сотрудника ГДЕ (employee.emp_id> '300') И employee.salary <'50000'
Пример 4
Выражения 100 в
строка Criteria и > 300 в
строка Или для столбца DeptId вместе
с выражением <50000 в
строка Criteria в столбце Salary , получить
информация для сотрудников:
Отдел 100 и имеют зарплату менее 50 000 долл. США
или
Отдел, ID которого больше 300, неважно какая у них зарплата
Оператор SELECT , созданный InfoMaker это:
ВЫБЕРИТЕ сотрудника.emp_id, employee.dept_id, сотрудник. зарплата ОТ сотрудника ГДЕ (employee.dept_id = '100') И (emplyee.salary <'50000') ИЛИ (employee.dept_id> '300')
Пример 5
Выражение IN (100,200) in
строка критериев в столбце DeptId в сетке
извлекает информацию для сотрудников, которые находятся в отделе 100 или 200.
Оператор SELECT , созданный InfoMaker это:
ВЫБЕРИТЕ сотрудника.emp_id,
employee.dept_id,
сотрудник. зарплата
ОТ сотрудника
ГДЕ employee.dept_id IN ('100,200')
Пример 6
В этом примере показано использование слова И в
строка Или критерии. В строке критериев > = 500 - это
в столбце EmpId и > = 30000 -
в столбце Зарплата . В строке "Или" - И <= 1000 находится в столбце EmpId и AND <= 50000 - это
в столбце Зарплата .Эти критерии извлекают
информация для сотрудников, имеющих ID сотрудника от 500 до 1000
и зарплата от 30 000 до 50 000 долларов.
Оператор SELECT , созданный InfoMaker это:
ВЫБЕРИТЕ employee.emp_id, employee.dept_id, сотрудник. зарплата ОТ сотрудника ГДЕ (((employee.emp_id> = '500') И (employee.salary> = '30000') И (employee.emp_id <= '1000') И (employee.salary <= '50000')))
Пример 7
В сетке с тремя столбцами: emp_last_name , emp_first_name ,
и зарплата , выражения НРАВИТСЯ
C% в строке Criteria и LIKE
G% в строке или столбца emp_last_name получить информацию о сотрудниках, чьи фамилии начинаются
с C или G.
Оператор SELECT , созданный InfoMaker это:
ВЫБЕРИТЕ employee.emp_last_name, employee.emp_first_name, сотрудник. зарплата ОТ сотрудника ГДЕ (((employee.emp_last_name LIKE 'C%')) ИЛИ ((employee.emp_last_name LIKE 'G%'))).