BrainWave| GE Healthcare (Russia)
Врачи все чаще выбирают функциональную магнитно-резонансную томографию (фМРТ) в качестве средства диагностики для картирования нейрофункций головного мозга. Метод фМРТ выполняет картирование неврологических функций по данным МРТ головного мозга на основе BOLD-контрастности (blood oxygenation level dependent contrast — контрастность, зависящая от степени насыщения крови кислородом) и является неинвазивным нерадиоактивным методом, который не требует введения радиоактивного изотопа или контрастного вещества. Данный метод измеряет повышенный уровень кислорода в исследуемой области, который является реакцией на внешние раздражители.
Ценная информация, получаемая в результате анализа карт, созданных с помощью этого метода, впоследствии используется в хирургических инвазивных процедурах. Таким образом технология, которая помогает создавать точные отчеты по конкретным исследованиям, чрезвычайно важна.
Программное обеспечение для визуализации BrainWave — это простой в использовании инструмент анализа и просмотра изображений, отражающих работу головного мозга и полученных с помощью приложения BrainWave RT.
Особенности системы:
- Обеспечивает быструю ретроспективную коррекцию движения, обусловленного подвижностью пациента.
- Активационные карты, создаваемые с помощью анализа на базе обобщенной линейной модели (Generalized Lineal Model, GLM).
- Отвечающие современным требованиям методы визуализации для совмещения результатов анализа с анатомическими данными.
- Четкая визуализация с цветными картами активации и интерактивной пороговой классификацией.
- Возможность сохранения форм в частной DICOM-структуре для быстрого анализа.
- Полная интеграция с системами видеосъемки, передачи по сети и архивирования AW.
- Полное соответствие стандарту DICOM.
- Возможность передавать данные активации в виде наборов данных трехмерных DICOM-изображений в высоком разрешении в рамках неврологических исследований.


Ощущения
Ощущения определяют как процесс отражения отдельных свойств предметов и явлений объективного мира при их непосредственном воздействии на рецепторы.
К этому можно, пожалуй, добавить лишь то, что ощущения отражают и состояние организма самого субъекта с помощью рецепторов, расположенных в его теле. Ощущения являются исходным источником познания, важным условием формирования психики и ее нормального функционирования. С этим, однако, согласны не все. Достаточно вспомнить мнение Р. Декарта, считавшего, что ощущения вводят нас в заблуждение, а истинные знания дает лишь разум, мышление.
Й. Мюллер, а позже и Б. Рассел утверждали, что ощущения не дают знаний о мире, следовательно, мир непознаваем. Б. Рассел ссылается при этом на явления фантомных болей (например, локализованных в ампутированной части руки или ноги). Й. Мюллер предложил «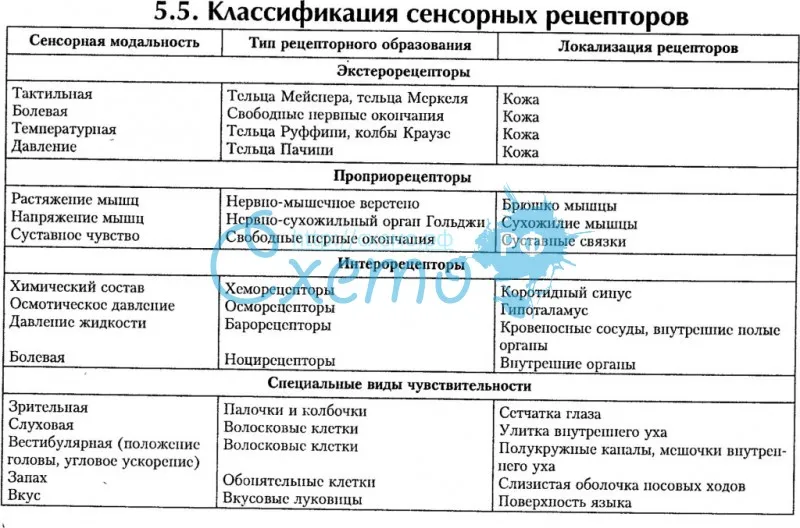 В результате ощущение не отражает свойства раздражителя, оно лишь субъективное состояние сознания. Хотя действительно механическое воздействие на глаз вызывает зрительные ощущения («искры из глаз», цветные пятна), легко показать, что в целом ощущения правильно отражают свойства объективного мира, а приведенный факт свидетельствует скорее о тонкой специализации органов чувств, в процессе эволюции подготовленных к отражению определенных форм энергии. Критерием для отделения ложных показаний органов чувств от истинных является действие, практика.
В результате ощущение не отражает свойства раздражителя, оно лишь субъективное состояние сознания. Хотя действительно механическое воздействие на глаз вызывает зрительные ощущения («искры из глаз», цветные пятна), легко показать, что в целом ощущения правильно отражают свойства объективного мира, а приведенный факт свидетельствует скорее о тонкой специализации органов чувств, в процессе эволюции подготовленных к отражению определенных форм энергии. Критерием для отделения ложных показаний органов чувств от истинных является действие, практика.
Потребность в постоянном получении ощущений хорошо проявляется в том случае, когда внешние раздражители отсутствуют (при сенсорной изоляции). Как показали эксперименты, в этом случае психика перестает нормально функционировать: возникают галлюцинации, нарушается мышление, отмечается патология восприятия своего тела и т.д. Специфические проблемы психологического характера возникают при сенсорной депривации, т.е. при ограничении притока внешних воздействий, что хорошо известно на примере развития психики людей слепых или глухих, а также плохо видящих и слышащих.
Ощущения человека чрезвычайно разнообразны, хотя со времен Аристотеля очень долго говорили лишь о пяти чувствах — зрении, слухе, осязании, обонянии и вкусе. В XIX в. знания о составе ощущений резко расширились в результате описания и изучения их новых видов, таких как вестибулярные, вибрационные, «мышечно-суставные» или кинестетические и т.д., а также путем дифференциации некоторых сложных сенсорно-перцептивных образований: например, осязание — это сочетание тактильных, температурных, болевых ощущений и кинестезии, а в тактильных ощущениях можно выделить ощущения прикосновения и давления. Увеличение количества видов ощущений поставило задачу их классификации.
Основные классификации ощущений
Известны несколько попыток классификации ощущений по разным основаниям или принципам. Явно неудачным оказалось предложение сгруппировать сенсорные функции по пространственному или временному признаку. Например, зрение и вестибулярные ощущения относили к «пространственным» чувствам, а слух и обоняние — к «временным», хотя дифференцировка пространственных и временных свойств объекта, как отмечает Б. Г. Ананьев, относится к ощущениям любой модальности. Не стало популярным и предложение В. Вундта группировать ощущения по видам раздражителей, которые их вызывают — механических, физических и химических, например зрительные и слуховые ощущения относятся к «
Г. Ананьев, относится к ощущениям любой модальности. Не стало популярным и предложение В. Вундта группировать ощущения по видам раздражителей, которые их вызывают — механических, физических и химических, например зрительные и слуховые ощущения относятся к «
Более детально разработанной и известной является классификация, использующая уровневый подход (Г. Хэд). Протопатическая чувствительность, более ранняя по происхождению и примитивная, тесно связана с эмоциями, далека от мышления, она менее дифференцированная и локализованная. Относящиеся к ней ощущения трудно разделять на категории и обозначать словами, описывать. Эпикритическая чувствительность выше по уровню, возникает позже и обладает, по сути, противоположными характеристиками: связь с мышлением, отдаленность от эмоциональных состояний, большая дифференцированность, категориальные названия для ощущений (красный, синий цвет, а не «запах мяты» или « В работе едва ли не каждого органа чувств есть элементы как протопатической, так и эпикритической чувствительности, хотя их соотношение неодинаково.
В работе едва ли не каждого органа чувств есть элементы как протопатической, так и эпикритической чувствительности, хотя их соотношение неодинаково.
Наконец, классификация Ч. Шеррингтона оказалась самой удачной, продуманной и логически строгой. Ч. Шеррингтон выделил три вида рецептивных полей: интероцептивные, проприоцептивные и экстероцептивные. В первом случае рецепторы находятся во внутренних органах (например, в стенках желудка), во втором — в мышцах, связках и сухожилиях, а третье рецептивное поле совпадает с наружной поверхностью организма и полностью открыто для внешних воздействий.
Интроцептивные ощущения сигнализируют о состоянии внутренних процессов организма. Они являются наиболее древними и элементарными, но очень полезны как сигналы соматических болезней или просто отклонений от нормального функционирования организма. Хорошо известно, как трудно иногда описать эти ощущения в кабинете врача. К тому же импульсы от внутренних органов, если они слабы, часто не вызывают осознанных ощущений.
Проприоцептивные ощущения играют важнейшую роль в регуляции движений (достаточно вспомнить, что бывает при попытке идти, когда человек «отсидел ногу»), они дают информацию о положении в пространстве тела и его частей, без них было бы невозможно осязание.
Третью, самую большую группу ощущений (экстероцептивные), Ч. Шеррингтон разделил на контактные и дистантные. Первые вызываются приложением воздействия непосредственно к поверхности тела, а вторые возникают, когда раздражитель действует с некоторой дистанции (обоняние, слух, зрение). В процессе эволюции именно дистантные экстероцептивные ощущения начинают играть все большую роль в познании окружающего и в организации поведения, поскольку они обеспечивают важное преимущество, позволяя заблаговременно получать нужную информацию об изменениях в среде и своевременно на них реагировать.
Проблема классификации ощущений все еще далека от решения. Б.Г. Ананьев отмечает, что в последние десятилетия не было разработано новых принципов их систематики и классификации, хотя научные знания об отдельных сенсорных системах и общих законах их развития заметно расширились и углубились.
Свойства ощущений
Каким бы ни было ощущение, его можно описать с помощью нескольких характеристик, свойств, присущих ему. Первая из них — модальность.
Модальность является качественной характеристикой, в которой проявляется специфичность ощущения как простейшего психического сигнала по сравнению с сигналом нервным (Л.М. Веккер). Прежде всего, выделаются такие виды ощущений, как зрительные, слуховые, обонятельные и т.д. Однако и каждый вид ощущений имеет свои модальные характеристики. Для зрительных ощущений таковыми могут быть цветовой тон, светлота, насыщенность; для слуховых — высота тона, тембр, громкость; для тактильных — твердость, шероховатость и т.д. Характеристики ощущения могут совпадать или не совпадать с физическим описанием качеств раздражителя, которые соответствуют этим модальным характеристикам. Пример совпадения — твердость и упругость, а несовпадения — цветовой тон, соответствующий частоте электромагнитных колебаний.
Еще одна (пространственная) характеристика ощущений — их локализация. Иногда (как, например, в случае болевых и интероцептивных, «внутренних» ощущений) локализация затруднена, неопределенна. Другая проблема состоит в объяснении «объективированности» ощущений, их «вынесенности» вне нас, хотя вызывающие их физиологические процессы протекают в анализаторе. Этот вопрос подробно обсуждает А.Н. Леонтьев. Именно предметность, т.е. отнесенность к действительности, создает ощущение как психическое явление. Умение «проецировать» ощущения во вне приобретается, вероятно, очень рано, и решающую роль играют при этом практические действия, моторика. Сначала предметная действительность выступает как объект приспособления организма, осуществляющегося в реальных контактах с ней. Интересна в этом отношении «проблема зонда»: когда мы пишем или режем что-то, ощущения локализованы на кончике ручки или ножа, т.е. совсем не там, где зонд контактирует с кожей, воздействует на нее.
Иногда (как, например, в случае болевых и интероцептивных, «внутренних» ощущений) локализация затруднена, неопределенна. Другая проблема состоит в объяснении «объективированности» ощущений, их «вынесенности» вне нас, хотя вызывающие их физиологические процессы протекают в анализаторе. Этот вопрос подробно обсуждает А.Н. Леонтьев. Именно предметность, т.е. отнесенность к действительности, создает ощущение как психическое явление. Умение «проецировать» ощущения во вне приобретается, вероятно, очень рано, и решающую роль играют при этом практические действия, моторика. Сначала предметная действительность выступает как объект приспособления организма, осуществляющегося в реальных контактах с ней. Интересна в этом отношении «проблема зонда»: когда мы пишем или режем что-то, ощущения локализованы на кончике ручки или ножа, т.е. совсем не там, где зонд контактирует с кожей, воздействует на нее.
Интенсивность — это классическая количественная характеристика.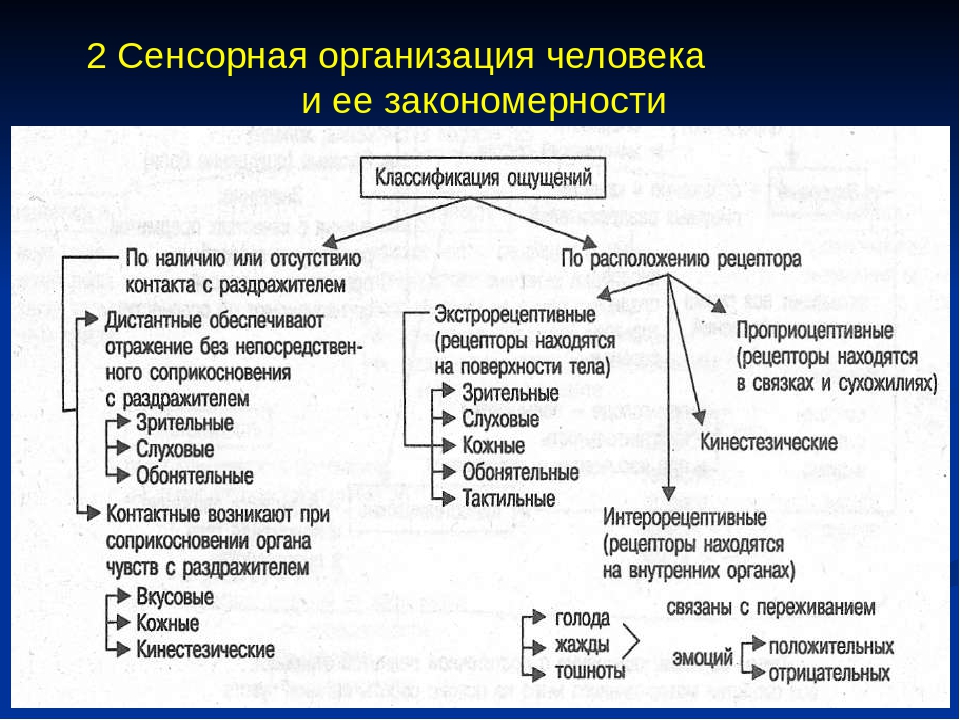 Проблема измерения интенсивности ощущения является одной из главных в психофизике. Г. Фехнер считал, что субъект не может непосредственно количественно оценить свои ощущения. Однако С. Стивене не согласился с этим. Им были разработаны так называемые прямые методы оценки интенсивности ощущения, например, когда испытуемый должен оценить в каких-то единицах (баллах, процентах и т.д.) величину раздражителя по сравнению с образцом.
Проблема измерения интенсивности ощущения является одной из главных в психофизике. Г. Фехнер считал, что субъект не может непосредственно количественно оценить свои ощущения. Однако С. Стивене не согласился с этим. Им были разработаны так называемые прямые методы оценки интенсивности ощущения, например, когда испытуемый должен оценить в каких-то единицах (баллах, процентах и т.д.) величину раздражителя по сравнению с образцом.
Основной психофизический закон отражает связь между величиной ощущения и величиной действующего раздражителя. Известны такие варианты основного психофизического закона, как логарифмический закон Г. Фехнера, степенной закон С. Стивенса, а также предложенный Ю.М. Забродиным обобщенный психофизический закон. Законы Фехнера и СТивенса оказываются при этом частными случаями последнего.
Следующая (временная) характеристика ощущения — его длительность. Ощущение возникает позже, чем начинает действовать раздражитель, и не исчезает сразу с его прекращением. Период от начала действия раздражителя до возникновения ощущения называют латентным (скрытым) периодом ощущения. Он неодинаков для разных видов ощущений (для тактильных — 130 мс, для болевых — 370 мс, для вкусовых — 50 мс) и может резко меняться при заболеваниях нервной системы.
Период от начала действия раздражителя до возникновения ощущения называют латентным (скрытым) периодом ощущения. Он неодинаков для разных видов ощущений (для тактильных — 130 мс, для болевых — 370 мс, для вкусовых — 50 мс) и может резко меняться при заболеваниях нервной системы.
После прекращения действия раздражителя его след некоторое время сохраняется в виде последовательного образа, который может быть либо положительным (соответствующим по характеристикам стимулу), либо отрицательным (обладающим противоположными характеристиками, например, окрашенным в дополнительный цвет). Положительные последовательные образы мы обычно не замечаем из-за их кратковременности. Лучше всего изучены зрительные последовательные образы, хотя они имеют место и в ощущениях других модальностей. Последовательные образы в основном определяются процессами на периферии анализатора, но зависят также и от нейродинамики в его центральном отделе. Например, его длительность в зрительной сфере резко увеличивается у больных, страдающих галлюцинациями.
Возможности анализаторов отражать отдельные свойства раздражителей или тонкие различия между ними характеризуют пороги ощущений. Нижний абсолютный порог — это минимальная величина раздражителя, вызывающая ощущение. Верхним абсолютным порогом называют максимальную величину раздражителя, при которой ощущение исчезает либо качественно меняется (например, превращается в болевое). Минимальное изменение интенсивности раздражителя или его другого свойства, вызывающее изменение ощущения, — это разностный (или дифференциальный) порог. Величина, обратно пропорциональная порогу ощущения, называется чувствительностью. Наличие порогов предохраняет нас от информационной перегрузки и некоторых биологически вредных воздействий.
Частное от деления разностного порога на исходную величину раздражителя, от которой он увеличивается или уменьшается, называют иногда относительным порогом. Эта величина (в отличие от разностного порога) в широком диапазоне изменения стимула является постоянной для ощущений определенной модальности. Например, для ощущения давления она равна приблизительно V30, для силы звука — Vio, а для силы света — Vioo-Последнее означает, что к 100 одинаковым лампочкам нужно добавить еще одну такую же, чтобы изменение освещения стало заметным.
Например, для ощущения давления она равна приблизительно V30, для силы звука — Vio, а для силы света — Vioo-Последнее означает, что к 100 одинаковым лампочкам нужно добавить еще одну такую же, чтобы изменение освещения стало заметным.
От порога осознанного ощущения необходимо отличать физиологический порог. Он оказывается преодоленным, когда энергии воздействия достаточно для мозгового возбуждения. Порог осознанного ощущения всегда выше физиологического: для возбуждения рецептора в сетчатке достаточно 1 фотона, но светящаяся точка может быть видна лишь при действии 5—8 фотонов. Между этими порогами лежит подпороговая зона,’ или область субсенсорных раздражителей, которые не ощущаются, однако вызывают ряд объективно регистрируемых реакций (например, таких как кожно-гальванический или улитко-зрачковый рефлекс). Физиологический порог — это величина достаточно стабильная, поскольку он в основном определяется генетически.
Изучением и измерением порогов ощущений занимается психофизика, основателем которой считают Г. Фехнера (1860), Острая дискуссия развернулась вокруг понятия нижнего абсолютного порога. Пороговая концепция считала сенсорный ряд дискретным. Нижний абсолютный порог при этом рассматривается как ноль на шкале ощущений, и, начиная с этой границы, раздражитель всегда вызывает ощущение. Такому взгляду противоречили факты непостоянства получаемого в опыте значения порога. Возникла концепция непрерывности сенсорного ряда, согласно которой от теоретического понятия порога как определенной точки на континууме следует отказаться. Изменение же операционального порога, получаемого в процессе измерения, объяснялось постоянно меняющимся соотношением между благоприятными и неблагоприятными факторами, влияющими на порог. При этом сколь угодно слабый раздражитель иногда может вызывать ощущение. К.В. Бардин подробно анализирует разные подходы к решению пороговой проблемы. Лучшим ее решением сейчас является, вероятно, психофизическая модель теории обнаружения сигнала, по которой сенсорный эффект от раздражителя всегда суммируется с обусловленным собственным возбуждением в сенсорной системе.
Фехнера (1860), Острая дискуссия развернулась вокруг понятия нижнего абсолютного порога. Пороговая концепция считала сенсорный ряд дискретным. Нижний абсолютный порог при этом рассматривается как ноль на шкале ощущений, и, начиная с этой границы, раздражитель всегда вызывает ощущение. Такому взгляду противоречили факты непостоянства получаемого в опыте значения порога. Возникла концепция непрерывности сенсорного ряда, согласно которой от теоретического понятия порога как определенной точки на континууме следует отказаться. Изменение же операционального порога, получаемого в процессе измерения, объяснялось постоянно меняющимся соотношением между благоприятными и неблагоприятными факторами, влияющими на порог. При этом сколь угодно слабый раздражитель иногда может вызывать ощущение. К.В. Бардин подробно анализирует разные подходы к решению пороговой проблемы. Лучшим ее решением сейчас является, вероятно, психофизическая модель теории обнаружения сигнала, по которой сенсорный эффект от раздражителя всегда суммируется с обусловленным собственным возбуждением в сенсорной системе. Для принятия решения о том, был ли сигнал на фоне шума, испытуемый использует критерий, который выбирается им в зависимости от ряда факторов (например, от цены ошибок типа «пропуск сигнала» и «ложная тревога»).
Для принятия решения о том, был ли сигнал на фоне шума, испытуемый использует критерий, который выбирается им в зависимости от ряда факторов (например, от цены ошибок типа «пропуск сигнала» и «ложная тревога»).
При измерении порогов ощущений они в разное время у одного и того же человека могут значительно различаться. Это объясняется действием многих факторов. Одни из них — экстренные —• изменяют пороги быстро, но не надолго. Другие — длительного действия — вызывают постепенное и устойчивое изменение порогов ощущений. Примером первых факторов может быть сенсорная адаптация, а ко вторым относится возраст. Кроме того, все эти факторы иногда делят на внешние (влияние окружения) и внутренние (изменения в организме).
Сенсорная адаптация — это изменение порогов ощущений при действии постоянного раздражителя. При полной адаптации ощущение вообще отсутствует. Таким образом, не допускается перераздражение анализаторов и обеспечивается чувствительность к очень слабым воздействиям. Особенно ярко адаптация выражена в тактильных, температурных, обонятельных и зрительных ощущениях. Например, после пребывания в темноте в течение часа световая чувствительность увеличивается примерно в 200 000 раз. Практически отсутствует адаптация к звуковым и болевым воздействиям. Адаптация идет с отрицательным ускорением, т.е. в первое время наиболее быстро. Она зависит от интенсивности раздражителя и площади, на которую он действует.
Особенно ярко адаптация выражена в тактильных, температурных, обонятельных и зрительных ощущениях. Например, после пребывания в темноте в течение часа световая чувствительность увеличивается примерно в 200 000 раз. Практически отсутствует адаптация к звуковым и болевым воздействиям. Адаптация идет с отрицательным ускорением, т.е. в первое время наиболее быстро. Она зависит от интенсивности раздражителя и площади, на которую он действует.
Чувствительность возрастает при действии слабого около порогового (или порогового) раздражителя соответствующей модальности. Это явление было изучено А.И. Бронштейном и названо им сенсибилизация, хотя часто данный термин используют в другом значении. Например, А.Р. Лурия относит к сенсибилизации случаи повышения чувствительности под влиянием физиологических или психологических изменений организма.
На пороги ощущений влияет мотивация, биологическая или социальная значимость раздражителя. Например, при создании интересной игровой ситуации дети демонстрируют более высокую остроту зрения по сравнению с ее измерением в обычных лабораторных условиях. Очень слабо светящаяся точка становится видимой лишь после того, как ей придается сигнальное значение (в эксперименте Г.В. Гершуни испытуемые, заметив точку, могли избежать удара током).
Пороги ощущений можно значительно понизить специальными упражнениями, тренировкой. А.Н. Леонтьев, используя пропевание испытуемыми предъявляемых им для различения звуков, добивался того, что за несколько часов тренировки пороги снижались в 6—8 раз. При накоплении профессионального опыта к длительной тренировке добавляется фактор значимости, поэтому результаты особенно впечатляющи. Например, опытный шлифовальщик на глаз замечает просвет в 0,0005 мм, а непрофессионал — 0,01 мм.
С возрастом, под влиянием роста и созревания соответствующих структур мозга, пороги ощущений у ребенка понижаются. В частности, хорошо известно, что по мере взросления значительно улучшается цветоразличение и повышается острота зрения. При старении процесс идет в обратном направлении. Так же постепенно теряется чувствительность к высокочастотным звукам.
Изменения в обменных процессах, эндокринные расстройства (в частности, гиперфункция щитовидной железы) тоже влияют на пороги. У беременных обостряется обонятельная чувствительность, но повышаются пороги зрительных и слуховых ощущений, что биологически полезно.
К экстренным факторам можно отнести утомление, снижающее чувствительность, и воздействие некоторых фармакологических препаратов и химических веществ.
Изменить пороги ощущений могут «побочные» раздражители, т.е.» воздействия другой модальности. Наконец, еще один способ — создание условной временной связи. Если слово «темнота» сопровождать включением света, вырабатывается второ-сигнальный условный рефлекс, когда произнесение этого слова будет повышать световую чувствительность.
Мозг человека функционирует как единая, целостная система, поэтому процессы, протекающие в одном анализаторе, зависят от процессов в других органах чувств. Эту идею целостности организма (и нервной системы в частности) анализирует Б.Г. Ананьев, обсуждая сенсорно-перцептивную организацию человека и называя головной мозг единым гигантским анализатором. Рассмотрим два проявления взаимодействия органов чувств.
В результате действия раздражителей на один анализатор могут повышаться или понижаться пороги ощущений другой модальности. Это явление изучено СВ. Кравковым, и, по его данным, оно наблюдается в отношении всех видов ощущений. Например, сильный шум снижает остроту центрального зрения, а слабый — повышает. Под влиянием сладкого, соленого и кислого чувствительность зрения повышается, а под влиянием горького — снижается. Изменения могут достигать нескольких десятков процентов от первоначальной величины и сохраняться долго. При психической патологии и поражении мозга (в частности, после сотрясения мозга) взаимодействие анализаторов часто усиливается, ослабляется или извращается, что целесообразно использовать в целях диагностики. Результат изменения порогов при воздействии на другой анализатор легко заметить и без лабораторных исследований. Еще М.В. Ломоносов писал, что на морозе цвета ярче. Более громким кажется голос лектора в темноте, когда он использует диапозитивы.
Предложено несколько теорий, объясняющих механизмы данного явления: взаимодействие процессов в близко расположенных афферентных нервных волокнах разных анализаторов; вегетативная нервная система как главный посредник в межанализаторных влияниях. Еще один подход отводит решающую роль межцентральным влияниям в коре головного мозга, активации центральных отделов анализаторов (например, нюханье веществ, не имеющих запаха, также повышает зрительные пороги). Единая общая теория пока отсутствует.
Другое проявление взаимодействия анализаторов — явление синестезии. В узком (строгом) значении — это возникновение ощущений одной модальности при действии раздражителя другой модальности. Истинная синестезия встречается очень редко (один случай на несколько тысяч человек).
Синестезией иногда также называют появление целостных образов-представлений при действии раздражителя другой модальности. Наконец, о синестезичности говорят и при характеристике стимула одной модальности в терминах качеств другой модальности ощущений (острый звук, прохладный цвет и т.д.) или когда удается легко подбирать устойчивое соответствие между запахом и цветом, запахом и звуком и т.д.
Для объяснения синестезии часто ссылаются на выработку устойчивых условных временных связей между анализаторами (обычно в детском возрасте). Это соответствует, в частности, большим качественным индивидуальным различиям, когда у разных людей какой-то звук связан с разными цветами. В то же время есть данные о том, что в основе синестезии лежат объективные свойства раздражителей (как правило, более темные оттенки серого цвета подбирают к запахам веществ, в молекулах которых больше атомов углерода). Известно также, что синестезия ярче выражена у людей с повышенной возбудимостью подкорковых образований, она может повышаться в период беременности и при приеме наркотиков.
Интересен один аспект синестезии, изучаемый А.П. Журавлевым в русле так называемой фоносемантики. Были установлены типичные зуко-цветовые соответствия: А — густо-красный, Е — зеленый, И — синий и т.д. Анализ стихотворений известных поэтов показал, что во многих случаях цветовая палитра, описанная словами, хорошо соответствует цветовому значению звукобукв, встречающихся в тексте значительно чаще среднего показателя.
Классификации рецепторов — Технарь
Сенсоры различают по специфичной чувствительности к различным стимулам, по строению и местоположению. Специфичная чувствительность рецепторов к раздражителям различной природы (механическим, хим, температурным и т. д.) обоснована различными механизмами управления ионными каналами плазматических мембран, состояние которых определяет появление рецепторного потенциала и переход от физиологического покоя к возбуждению. Стимулы, к которым сенсоры более чувствительны, именуются адекватными (лат. adaequatus — приравненный).
Механорецепторы возбуждаются посильнее всего вследствие деформации их клеточной мембраны при давлении либо растяжении, к ним относятся тактильные сенсоры кожи, проприоцепторы мускул и сухожилий, слуховые и вестибулярные сенсоры во внутреннем ухе, барорецепторы и волюморецепторы, находящиеся во внутренних органах и кровяных сосудах. Хеморецепторы возбуждаются вследствие присоединения к ним определенных хим молекул, они представлены обонятельными и вкусовыми сенсорами, также хемочувствительнымирецепторами внутренних органов и кровеносных сосудов.
Для расположенных в сетчатке глаза фоторецепторов адекватным раздражителем являются поглощенные ими кванты света, для терморецепторов (холодовых и термических) — конфигурации температуры.
В итоге действия раздражителя практически у всех рецепторов возрастает проницаемость клеточной мембраны для катионов, что приводит к ее деполяризации. Исключением из общепринятого правила являются фоторецепторы, где после поглощения энергии квантов света в связи особенностями управления ионными каналами происходит гиперполяризация мембраны. Изменение величины мембранного потенциала рецепторов в ответ на действие стимула представляет собой рецепторный потенциал — входной сигнал первичных сенсорных нейронов. Если величина рецепторного потенциала достигнет критичного уровня деполяризации либо превзойдет его, генерируются потенциалы деяния, при помощи которых сенсорные нейроны передают в центральную нервную систему информацию о действующих стимулах.
Информация о силе действующего на сенсоры стимула кодируется 2-мя методами: частотой потенциалов деяния, возникающих в сенсорном нейроне (частотное кодирование), и числом сенсорных нейронов, возбудившихся в ответ на действие стимула. При увеличении силы действующего на сенсоры раздражителя увеличивается амплитуда рецепторного потенциала, что, обычно, сопровождается повышением частоты потенциалов деяния в сенсорном нейроне первого порядка. Чем обширнее имеющийся частотный спектр потенциалов деяния у сенсорных нейронов, тем большее число промежных значений силы раздражителя способна различать сенсорная система. Первичные сенсорные нейроны схожей модальности различаются порогом возбуждения, потому при действии слабеньких стимулов возбуждаются только более чувствительные нейроны, но с повышением силы раздражителя на него реагируют и наименее чувствительные нейроны, имеющие более высочайший порог раздражения. Чем больше первичных сенсорных нейронов возбудится сразу, тем посильнее будет их совместное действие на общий нейрон второго порядка, что в конечном итоге отразится на личной оценке интенсивности действующего раздражителя.
Продолжительность чувства находится в зависимости от реального времени меж началом и прекращением воздействия на сенсоры, также от их возможности уменьшать либо даже прекращать генерацию нервных импульсов при длительном действии адекватного стимула. При продолжительном действии стимула порог чувствительности рецепторов к нему может повышаться, что определяется как адаптация рецепторов. Механизмы адаптации не схожи в сенсорах различных модальностей, посреди их различают стремительно адаптирующиеся (к примеру, тактильные сенсоры кожи) и медлительно адаптирующиеся сенсоры (к примеру, проприоцепторы мускул и сухожилий). Стремительно адаптирующиеся сенсоры посильнее возбуждаются в ответ на резвое нарастание интенсивности стимула (фазический ответ), а их стремительная адаптация содействует освобождению восприятия от на биологическом уровне малозначительной инфы (к примеру, контакт меж кожей и одежкой). Возбуждение медлительно адаптирующихся рецепторов не достаточно находится в зависимости от скорости конфигурации стимула и сохраняется при его продолжительном действии (тонический ответ), потому, к примеру, неспешная адаптация проприоцепторов позволяет человеку получать подходящую ему для сохранения позы информацию в течение всего нужного времени.
Есть сенсорные нейроны, генерирующие потенциалы деяния спонтанно, т. е. при отсутствии раздражения (к примеру, сенсорные нейроны вестибулярной системы), такая активность именуется фоновой. Частота нервных импульсов в этих нейронах может возрастать либо уменьшаться зависимо от интенсивности действующего на вторичные сенсоры стимула, не считая того, она может определяться направлением, в каком отклоняются чувствительные волоски механорецепторов. К примеру, отклонение волосков вторичных механорецепторов в одну сторону сопровождается увеличением фоновой активности сенсорного нейрона, которому они принадлежат, а в обратную сторону — снижением его фоновой активности. Обозначенный метод рецепции позволяет получать информацию и об интенсивности стимула, и о направлении, в каком он действует.
Рецептивные поля органов эмоций живых системРецептивным полем именуется область, занимаемая совокупой всех рецепторов, стимуляция которых приводит к возбуждению сенсорного нейрона (рис. 17.1). Наибольшая величина рецептивного поля первичного сенсорного нейрона определяется местом, которое занимают все ветки его периферического отростка, а число рецепторов, имеющихся в этом пространстве, показывает на плотность иннервации. Высочайшая плотность иннервации смешивается, обычно, с малыми размерами рецептивных полей и, соответственно, высочайшим пространственным разрешением, позволяющим различать стимулы, действующие на примыкающие рецептивные поля. Малые рецептивные поля типичны, к примеру, для центральной ямки сетчатки и для пальцев рук, где плотность рецепторов существенно выше, чем на периферии сетчатки либо в коже спины, для которых свойственны большая величина рецептивных полей и наименьшее пространственное разрешение. Рецептивные поля примыкающих сенсорных нейронов могут отчасти перекрывать друг дружку, потому информация о действующих на их стимулах передается не по одному, а по нескольким параллельным аксонам, что увеличивает надежность ее передачи.
Рис. 12.2 Рецептивные поля первичных сенсорных нейронов и сенсорных нейронов второго порядка. Рецептивные поля первичных сенсорных нейронов ограничены областью их чувствительных окончаний. Рецептивное поле переключательного нейрона появляется из суммы рецептивных полей конвергирующих к нему первичных сенсорных нейронов. Раздражение центральной либо периферической области рецептивного поля сенсорного нейрона второго и последующих порядков сопровождается обратным эффектом. Как видно на схеме, раздражение центра рецептивного поля вызовет возбуждение проекционного нейрона, а раздражение периферической области — торможение при помощи интернейронов переключательного ядра (латеральное торможение). В итоге контраста, создаваемого меж центром и периферией рецептивного поля, выделяется информация для передачи на последующий иерархический уровень.
Величина рецептивных полей сенсорных нейронов второго и последующих порядков больше, чем у первичных сенсорных нейронов, так как центральные нейроны получают информацию от нескольких конвергирующих к ним нейронов предыдущего уровня. От центра рецептивного поля информация передается конкретно к сенсорным нейронам последующего порядка, а от периферии — к тормозным интернейронам переключательного ядра, потому центр и периферия рецептивного поля являются реципрокными по отношению друг к другу. В итоге сигналы от центра рецептивного поля беспрепятственно добиваются последующего иерархического уровня сенсорной системы, тогда как сигналы, поступающие от периферии рецептивного поля, тормозятся (в другом варианте организации рецептивного поля легче пропускаются сигналы от периферии, а не от центра). Такая многофункциональная организация рецептивных полей обеспечивает выделение более важных сигналов, просто различаемых на контрастном с ними фоне.
Сенсорные нейроны низшего иерархического уровня способны передавать электронные сигналы сразу нескольким нейронам последующего уровня благодаря дивергенции к ним собственных аксонов. Это увеличивает надежность передачи инфы от 1-го иерархического уровня к другому и позволяет сохранять чувствительность сенсорной системы при утрате отдельных нейронов. Возбуждение нейронов более высочайшего иерархического уровня определяется не только лишь действием на их нейронов предшествующего уровня, да и механизмом латерального торможения, имеющегося в переключательных ядрах (рис. 12.3).
Рис. 12.3 Схема латерального торможения в сенсорных системах.
Степень возбуждения переключательных нейронов определяется интенсивностью раздражения центра либо периферии его рецептивного поля. В итоге возбуждение будет передаваться от одной части рецептивного поля на фоне торможения либо прерывания сигналов от другой его части, что делает эффект контрастирования, содействующий различению действующих стимулов.
Латеральное торможение происходит вследствие активации тормозных интернейронов переключательного ядра коллатералями возбуждающих нейронов. Чем посильнее возбужден релейный сенсорный нейрон, тем больше он активирует тормозные нейроны, которые подавляют активность примыкающих релейных нейронов. Релейные нейроны, передающие сигналы от центра рецептивного поля, посильнее других увеличивают активность тормозных нейронов, а их воздействие на примыкающие релейные нейроны делает еще огромным контраст меж возбужденными и тормозимыми нейронами, выделяя линию передачи избранных сигналов.
Механизмы реализации отрицательной и положительной оборотных связейВысшие уровни сенсорной системы регулируют переработку инфы в низших по отношению к ним переключательных ядрах средством нисходящего торможения либо усиления передаваемых сигналов. Нисходящее торможение (рис.12.3) происходит вследствие активации тормозных нейронов переключательного ядра, деятельность которых определяют нисходящие пути, начинающиеся на более высочайшем иерархическом уровне сенсорной системы. В итоге нисходящего торможения увеличивается порог афферентной синаптической передачи в переключательном ядре низшего уровня. Нисходящее торможение представляет собой регуляцию сенсорного восприятия по механизму отрицательной оборотной связи. Тормозные интернейроны переключательных ядер уменьшают частоту передаваемых на последующий уровень сигналов, в связи с чем, сенсорное чувство ослабляется.
Нисходящее усиление происходит при помощи возбуждающих интернейронов переключательного ядра, которые активизируются аксонами нейронов более высочайшего иерархического уровня. Вследствие этого в управляемом по механизму положительной оборотной связи переключательном ядре снижается порог синаптической передачи, что делает вероятным прохождение относительно слабеньких сигналов на последующий иерархический уровень. Одновременное внедрение устройств отрицательной и положительной оборотной связи позволяет освобождаться от лишней инфы, подавлять «шум», т. е. нейронную активность, не содержащую физиологически важных сообщений, и сразу выделять и усиливать сигналы, на которых будет сосредоточено внимание.
Большая часть раздражителей, воспринимаемых сенсорными системами, оказывает на их общее воздействие, так как сразу возбуждает различные типырецепторов. К примеру, прикасающийся к ограниченному участку кожи предмет может быть гладким либо шершавым, теплым либо прохладным, сухим либо мокроватым, а его давление на кожу может усиливаться либо слабеть, он может передвигаться в том либо ином направлении.
Зрительные чувства представляют собой сочетание определенной формы воспринимаемого предмета, его движения и композиции разных цветов, заполняющих ту либо иную деталь формы. Сенсорные системы организованы так, что информация о каждом компоненте всеохватывающего раздражителя передается сразу по нескольким параллельным путям, любой из которых относится к определенной субмодальности, как, к примеру, форма, движение либо цвет наблюдаемого зрительного объекта. В каждом из таких путей независимо друг от друга перерабатываются сигналы, несущие информацию о различных качествах подействовавшего на сенсоры всеохватывающего раздражителя. При всем этом различные каналы, применяемые для передачи сенсорной инфы, не дублируют друг дружку, в чем проявляется принцип многоканальности, присущий всем сенсорным системам. Высочайшая упорядоченность нейронных переключений на каждом иерархическом уровне обеспечивает поступление инфы от каждого рецептивного поля к подходящим конкретно ему кортикальным колонкам проекционной коры. Нужное для целостного восприятия раздражителя объединение инфы, относящейся к различным субмодальностям, происходит в сенсорных регионах коры.
Личное сенсорное восприятие, абсолютный порог чувства. Закон Вебера—Фехнера.Любая сенсорная система принимает действие адекватного стимула в ограниченном спектре значений его силы. Меньший по интенсивности стимул, способный вызвать чувство, именуется абсолютным порогом чувства. Его величина устанавливается опытным методом, она не схожа у различных людей и может изменяться у 1-го и такого же человека зависимо от многофункционального состояния, различаясь, к примеру, при хорошей работоспособности и утомлении. Величина абсолютного порога чувства выше абсолютного порога первичных сенсорных нейронов, потому что возбуждение отдельных чувствительных нейронов либо их маленький группы не непременно приводит к возбуждению высших уровней сенсорной системы вследствие тормозных процессов на прошлых иерархических уровнях. Потому не каждый стимул, возбуждающий сенсоры первичного сенсорного нейрона, лично чувствуется и осознается.
Величина, на которую один стимул надпорогового спектра должен отличаться от другого, чтоб их разницу можно было лично различить, получила заглавие дифференциального порога либо порога различения. Ощущаемый прирост интенсивности раздражения (дифференциальный порог) должен превосходить ранее действовавший стимул на определенную и постоянную величину.
Закон Вебера справедлив для различных модальностей, но только в границах раздражения средней силы, а при слабеньких и сильных стимулах способность к различению силы стимулов у человека понижается. Предложена также ступенчатая шкала интенсивности чувств, в какой за ноль принимается величина абсолютного порога, а зависимость меж интенсивностью стимула и чувством выражается как:
, (12.1)где Е — интенсивность чувства, к — неизменная величина, S — интенсивность стимула,
— абсолютный порог.Эта зависимость, получившая заглавие закона Вебера—Фехнера, указывает, что линейное повышение интенсивности чувства отражает логарифмический рост интенсивности стимула. Закон Вебера—Фехнера охарактеризовывает личную способность человека различать действующие стимулы (шкала различения), но не оценивает саму интенсивность чувств, так как этот закон основан на допущении равенства всех ее приростов, как к слабеньким, так и к сильным стимулам.
Личная оценка интенсивности раздражителя (высококачественное различие) растет при увеличении различия меж пороговой и действующей силой стимула (количественное различие). Но зависимость меж силой стимула и чувством не схожа при разной интенсивности стимулов, а поэтому имеет не линейный, а степенной нрав. Для оценки интенсивности чувств всего спектра употребляется шкала Стивенса, устанавливающая зависимость чувств от силы стимула в виде степенной функции:
, (12.2)где Е — интенсивность чувства, S — действующая сила стимула,
— абсолютный порог, к — константа шкалы, n — показатель степени, который находится в зависимости от сенсорной модальности (к примеру, для восприятия светового чувства он составляет 0,33, громкости звука — 0,6, а для проприоцептивных чувств — 1,7).Пространственные свойства действующих стимулов, нужные для их различения, зависят от специфичных особенностей каждой сенсорной системы и величины рецептивных полей. Прикосновение к коже дистальной фаланги пальца руки 2-ух ножек циркуля с расстоянием меж ними 2 мм чувствуется раздельно, но чтоб почувствовать раздельное прикосновение к коже спины, ножки циркуля нужно раздвинуть до 60 мм. Пространственное восприятие этих тактильных стимулов находится в зависимости от размеров соответственных рецептивных полей: раздельное чувство может быть только при условии раздражения каждой ножкой циркуля независящего рецептивного поля. Только тогда информация о каждом стимуле будет перерабатываться раздельно на каждом уровне организации сенсорной системы, включая проекционную область коры. Подобная ситуация имеет место при восприятии 2-ух точек зрительного поля: они не соединяются в одну, если отражаемые ими световые лучи попадут на различные рецептивные поля сетчатки. Имеет значение и степень контраста меж действующим стимулом и его фоном: отлично контрастируемые объекты (к примеру, темное на белоснежном) различаются легче, чем не достаточно контрастируемые (темное на сероватом).
Рецептивные поля различающихся своими сенсорами сенсорных нейронов перекрываются, потому при действии на кожу комплекса стимулов сразу возбуждаются различные виды рецепторов, что позволяет чувствовать все динамические и статические характеристики такового комплекса. Обработка и анализ инфы сигналов от разных рецепторов происходит на высших уровнях сенсорной системы, формирующих всеохватывающее восприятие действующих на кожа стимулов. Плотность механорецепторов в различных участках кожи не схожа, чем определяются различные характеристики пространственного дифференциального порога, т. е. меньшего расстояния меж 2-мя точками, раздражение каждой из которых чувствуется раздельно.
Классификация рецепторов. Общие свойства рецепторов. — Студопедия
Функциональное назначение рецепторов заключается в выявлении и различении раздражителей с разной модальности. Поскольку раздражители внешней или внутренней среды имеют различную природу, а нервные центры «понимают только один язык», т.е. нервный импульс (ПД), то к вышеперечисленным функциям рецептора предстоит еще и преобразования разной модальности раздражения в ПД.
Классификация:
I.По характеру действия раздражителя:
1. Экстерорецепторы (рецепторы органов чувств).Как правило, высокоспецифичны к определенному раздражителю – мономодальные (закон специфической нервной энергии Мюллера).
2. Интерорецепторы (рецепторы внутренних органов). Могут быть мономодальные (например, хеморецепторы в стенке аорты чувствительны к СО2, рН, О2). Но большинство интерорецепторов полимодальные, т.е. могут реагировать на широкий диапазон раздражителей.
3. Проприорецепторы (рецепторы мышц, связок, суставов), полимодальные.
II. По модальности:
1. Механорецепторы – располагаются в периферических отделах соматической (скелетно-мышечной), слуховой и вестибулярной систем.
2. Терморецепторы – располагаются в коже, внутренних органах и центральных термочувствительных нейронах, делятся на тепловые и холодовые.
3. Хеморецепторы. У наземных животных находятся в периферических отделах обонятельной и вкусовой сенсорных систем, а также в сосудах и тканях (информация о химическом составе внутренней среды).
4. Фоторецепторы – в глазу.
5. Электрорецепторы – в боковой линии рыб, у некоторых амфибий и т.д.
6. Болевые (ноцицептивные) рецепторы – во всех органах.
III. По механизмам образования нервного импульса:
1. Первично-чувствующие рецепторы:раздражитель действует на дендрит сенсорного нейрона, изменяется проницаемость клеточной мембраны к ионам (в основном к Na+), образуется локальный электрический потенциал (рецепторный потенциал), который электротонически распространяется вдоль мембраны к аксону (таблица 1). На мембране аксона образуется потенциал действия, передаваемый далее в ЦНС.
Сенсорный нейрон с первично-чувствующим рецептором представляет собой биполярный нейрон, на одном полюсе которого располагается дендрит с ресничкой, а на другом – аксон, передающий возбуждение в ЦНС. Примеры: проприорецепторы, терморецепторы, обонятельные клетки.
2. Вторично-чувствующие рецепторы: в них раздражитель действует на рецепторную клетку, в ней возникает возбуждение (рецепторный потенциал). На мембране аксона рецепторный потенциал активирует выделение нейромедиатора в синапс, в результате чего на постсинаптической мембране второго нейрона (чаще всего биполярного) образуется генераторный потенциал, который и приводит к образованию потенциала действия на соседних участках постсинаптической мембраны (таблица 1). Далее этот потенциал действия передается в ЦНС. Примеры: волосковые клетки уха, вкусовые
IV.По дальности расположения воспринимаемого стимула рецепторы являются
1. дистантными (слух, зрение),
2. контактными (осязание, обоняние, вкус).
3. интероцепторами — это рецепторы, воспринимающие раздражители из внутренней среды организма, (рецепторы сосудов, внутренних органов, а также рецепторы двигательного аппарата, называемые проприоцепторами).
Свойства рецепторов.
1. Высокая возбудимость. Так, для возбуждения фоторецептора сетчатки достаточно одного кванта света, для обонятельного рецептора — одной молекулы пахучего вещества.
2. Адаптация — уменьшение возбудимости рецепторов при длительном действии раздражителя (только темновая адаптация фоторецепторов приводит к повышению их возбудимости). Адаптация рецепторов выражается в снижении амплитуды РП и, как следствие, в уменьшении частоты импульсации в афферентном волокне.
3. Спонтанная активность, т.е. способность возбуждаться без действия раздражителя, присуща проприорецепторам, фоно-, фото-, вестибуло-, термо-, хеморецепторам. Эта способность связана со спонтанным колебанием проницаемости клеточной мембраны, перемещением ионов и периодической деполяризацией рецептора, которая, достигая критического уровня, приводит к генерации потенциала действия в афферентном нейроне.
При возбудимости рецепторов, обладающих более высокой фоновой активностью, даже слабый раздражитель способен значительно повысить частоту импульсации в них. Фоновая активность рецепторов участвует в поддержании тонуса ЦНС
Классификация модальности стимула от высокочастотной активности МЭГ человека между субъектами
Abstract
Анализ отдельных испытаний может выявить значимую динамику мозга, которая не видна при усреднении по испытаниям. Однако низкое отношение сигнал / шум (SNR) может препятствовать использованию методов однократного анализа и декодирования. В этом исследовании мы исследуем применимость однократного подхода к декодированию модальности стимула из магнитоэнцефалографической (МЭГ) высокочастотной активности.Чтобы классифицировать слуховое и визуальное представление слов, мы объединяем реконструкцию источника луча с методом случайной классификации лесов. Чтобы сделать вывод на групповом уровне, классификация встроена во внутрипредметную структуру. Мы показываем, что SNR гамма-излучения за одно испытание обеспечивает хорошую производительность классификации (точность по предметам: 66,44%). Это означает, что характеристики высокочастотной активности имеют высокую согласованность между испытаниями и субъектами. Классификатор случайного леса присвоил информационную ценность активности как слуховой, так и зрительной коры с высокой пространственной специфичностью.С течением времени мощность гаммы была наиболее информативной во время предъявления стимула. Среди всех частотных диапазонов полоса от 75 Гц до 95 Гц была наиболее информативной полосой частот как в зрительной, так и в слуховой областях. Широкий диапазон гамма-частот (от 55 Гц до 125 Гц), особенно в визуальных областях, способствовал успешной классификации. Таким образом, мы демонстрируем выполнимость подходов с одним испытанием для декодирования модальности стимула у субъектов от высокочастотной активности и описываем различительную гамма-активность во времени, частоте и пространстве.
Информация об авторе
Усреднение мозговой активности по испытаниям — мощный способ увеличить отношение сигнал / шум в данных MEG. Этот подход, однако, потенциально скрывает значимую динамику мозга, которая разворачивается на уровне одного исследования. Единичные пробные анализы были успешно применены к временной области или низкочастотной колебательной активности; его применение к высокочастотной активности МЭГ затруднено из-за низкой амплитуды этих сигналов. В настоящем исследовании мы показываем, что модальность стимула (визуальное или слуховое представление слов) может быть успешно декодирована из однократной высокочастотной активности MEG путем комбинирования реконструкции источника с алгоритмом классификации случайного леса.Этот подход позволяет выявить закономерности активности выше 75 Гц как в зрительной, так и в слуховой коре, подчеркивая важность высокочастотной активности для обработки доменно-специфических стимулов. Таким образом, наши результаты расширяют предыдущие выводы, выявляя высокочастотную активность в слуховой коре, связанную со слуховыми словесными стимулами в данных МЭГ. Кроме того, принятая межпредметная структура предполагает высокую согласованность между индивидуумами в высокочастотных моделях активности.
Образец цитирования: Westner BU, Dalal SS, Hanslmayr S, Staudigl T (2018) Межсубъектная классификация модальности стимула от высокочастотной активности МЭГ человека.PLoS Comput Biol 14 (3): e1005938. https://doi.org/10.1371/journal.pcbi.1005938
Редактор: Дэниел Буш, Университетский колледж Лондона, ВЕЛИКОБРИТАНИЯ
Поступила: 27.06.2017; Принята к печати: 20 декабря 2017 г .; Опубликован: 12 марта 2018 г.
Авторские права: © 2018 Westner et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Данные размещены в Open Science Framework, DOI 10.17605 / OSF.IO / M25N4: https://osf.io/M25N4/.
Финансирование: Эта работа была поддержана ERA-Net NEURON через Федеральное министерство образования и исследований Германии, http://www.neuron-eranet.eu/, грант 01EW1307, SSD; Европейский исследовательский совет, https://erc.europa.eu/, стартовый грант 640488 для SSD; Deutsche Forschungsgemeinschaft, http://www.dfg.de/, грант программы Emmy Noether HA 5622 / 1-1, SH; Европейский исследовательский совет, https: // erc.europa.eu/, Грант Консолидатора 647954, SH; Фонд Вольфсона и Королевское общество, https://royalsociety.org/grants-schemes-awards/grants/wolfson-research-merit/, SH; и Horizon 2020 Европейского союза, https://ec.europa.eu/programmes/horizon2020/, 661373, TS. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Со времени первых сообщений об активности кортикального гамма-диапазона [1, 2] эти высокочастотные ответы были связаны с множеством мозговых процессов и умственных задач, например, зрительным восприятием и обработкой [3–6], слуховым восприятием. [7, 8] или память [9–12]. Хотя были предложены многочисленные теории о происхождении и функции этих высокочастотных колебаний и их связи с более низкими частотами, такими как тета и альфа (например, [13-15]), продолжаются споры о том, отражают ли отклики гамма-диапазона узкополосные колебания или мощность широкополосного доступа увеличивается, возможно, отражая увеличение пиковой активности [6, 16, 17].Одним из препятствий на этом пути является характеристика 1/ f частотного спектра мощности мозга и активность гамма-диапазона с низким отношением сигнал / шум (SNR) в записях магнитоэнцефалографии (MEG) или электроэнцефалографии (EEG). Чтобы увеличить SNR, часто используется пробное усреднение для устранения случайной дисперсии. Однако этот подход потенциально может скрыть или отменить значимую активность мозга [18]. В самом деле, потенциалы локального поля и данные электрокортикографии обезьян выявили систематические изменения мощности и частоты гамма-излучения от опыта к опыту в зрительном [19] и в задаче памяти [20].Важно отметить, что средние значения по испытаниям в этих исследованиях демонстрируют классический устойчивый гамма-эффект, указывая на то, что ответы от одного испытания имеют решающее значение для понимания динамики мозга [18]. Одним из эффективных подходов к оценке информации, полученной в результате однократного испытания, являются методы многомерного декодирования. Однако остается неясным, применимы ли такие методы к данным МЭГ в гамма-диапазоне с низким отношением сигнал / шум. В настоящей статье мы исследуем прогностическую ценность мощности гамма-излучения при однократном испытании в отношении модальности предъявления стимула (слуховое или визуальное представление слов) в данных МЭГ человека.Хотя сопоставимые контрасты использовались для тестирования производительности классификатора или в качестве примеров наборов данных (например, [21, 22]), нашей целью было раскрыть однократные высокочастотные паттерны в данных МЭГ человека. Чтобы декодировать информацию о стимуле-модальности из частотно-временных данных, мы использовали комбинацию формирования диаграммы направленности [23] и классификации случайного леса [24]. Этот подход был встроен в структуру перекрестной проверки между субъектами, где классификатор был протестирован на единичных испытаниях невидимых субъектов для оценки общности пространственной частотно-временной модели.Наши результаты подтверждают, что гамма-отношение сигнал / шум в единичных испытаниях достаточно высоко для достижения стабильной точности классификации, значительно превышающей вероятность. Интересно, что модель классификации придает высокую информационную ценность широкой полосе пропускания в гамма-диапазоне. Кроме того, мы показываем, что характеристики гамма-активности достаточно схожи в разных испытаниях и даже у испытуемых, чтобы обеспечить надежную классификацию.
Материалы и методы
Заявление об этике
Исследование было одобрено институциональным наблюдательным советом Университета Констанца и в соответствии с Хельсинкской декларацией.
Участников
Всего в этом эксперименте МЭГ приняли участие 24 участника (17 женщин; средний возраст = 22 года, диапазон = 19–26 лет; 21 правша). Трое участников были исключены из-за технических проблем, один из-за чрезмерного шума окружающей среды. Здесь представлены данные остальных 20 участников. Все участники дали письменное информированное согласие до начала эксперимента и получили зачетные единицы курса или номинальную финансовую компенсацию за участие.Все участники были носителями немецкого языка и сообщили о нормальном зрении или его исправлении до нормального, а также об отсутствии в анамнезе неврологических заболеваний.
Части этих данных были опубликованы в [12] в отношении вопросов и анализов независимых исследований.
Конструкция, методика и материалы
Эксперимент состоял из фазы исследования и последующего теста распознавания. Здесь представлены только данные, полученные на этапе исследования. На этапе исследования участникам предъявляли слова либо визуально (проецируемое по центру экрана), либо слухом (через неферромагнитные трубки к обоим ушам).Продолжительность визуального представления слова определялась продолжительностью соответствующего аудиофайла, то есть временем произнесения слова (средняя продолжительность = 697 мс, с, , , , = 119 мс). Каждое слово сопровождалось фиксирующим крестом. Суммарная длительность креста слова и фиксации составила 2000 мс. Участникам было предложено подсчитать количество слогов в слове и указать с помощью нажатия кнопки, имеет ли слово два слога. Вопросительный знак (макс. Продолжительность 1500 мс) побуждал к ответу.Нажатие кнопки завершает отображение вопросительного знака. Перед каждым пунктом был представлен крест фиксации с переменной продолжительностью (от 1000 мс до 1500 мс). После фазы кодирования участники выполнили задачу отвлечения внимания и фазу проверки неожиданного распознавания.
Стимулы состояли из 420 несвязанных немецких существительных, сгруппированных в три списка по 140 слов. Половина слов каждого списка состояла из двух слогов, другая половина — из одного, трех или четырех слогов. Два списка были представлены на этапе исследования и один список на этапе тестирования.Распределение списков по этапам изучения или тестирования было сбалансировано для всех участников. Задания были представлены в случайном порядке с ограничением, что не более 5 слов одной и той же модальности и не более 5 слов из одного и того же условия были представлены последовательно.
Сбор и предварительная обработка данных MEG
ДанныеMEG были записаны с помощью 148-канального магнитометра (MAGNES 2500 WH, 4D Neuroimaging, Сан-Диего, США) в положении лежа на спине внутри комнаты с магнитным экраном.Данные непрерывно записывались с частотой дискретизации 678,17 Гц и полосой пропускания от 0,1 до 200 Гц, а затем субдискретизировались до 300 Гц для уменьшения вычислительной нагрузки. Вся обработка данных перед классификацией выполнялась с использованием FieldTrip [25], набора инструментов MATLAB с открытым исходным кодом для анализа данных MEEG. Данные были разделены на отдельные испытания с периодами от 1500 мс до представления элемента до 4000 мс после представления элемента. Испытания были визуально проверены на предмет артефактов, зараженные испытания были отклонены.После этого испытания были скорректированы на моргания, движения глаз и сердечные артефакты с использованием независимого компонентного анализа (ICA).
Реконструкция источника
Для сопоставления с индивидуальным структурным магнитно-резонансным изображением (доступно для 17 из 20 участников; для остальных трех участников мы использовали аффинную трансформацию мозга MNI-шаблона; Монреальский неврологический институт, Монреаль, Канада), форма участника Голова, а также три маркера (назион, левый и правый слуховой проход) и расположение катушек индикатора положения головы (HPI) были оцифрованы перед экспериментом с использованием 3D-сканера Fastrak Polhemus (Polhemus, Colchester, VT, USA).
Космическая активность с одним пробным источником была реконструирована с использованием формирователя луча с линейно ограниченной минимальной дисперсией (LCMV) [23] с нормализацией веса (индекс нейронной активности; [23, 26]). Сначала был вычислен пространственный фильтр с использованием реалистичной модели головы с одной оболочкой [27], основанной на отдельном структурном магнитно-резонансном изображении (МРТ) и исходной модели с точками сетки, покрывающими весь объем мозга (разрешение: 15 мм). Ковариационная матрица данных вычислялась для значений от -500 мс до 1000 мс относительно предъявления стимула.Чтобы учесть ранговый дефицит данных, который был введен применением ICA, ковариационная матрица была регуляризована путем загрузки ее диагонали 5% мощности датчика. Впоследствии к единичным испытаниям применялся пространственный фильтр для получения виртуальных электродов во всех точках сетки.
Для классификации колебательной активности были рассчитаны временные частотные представления для каждого виртуального электрода с применением быстрого преобразования Фурье.Активность гамма-диапазона оценивалась с использованием частотного сглаживания (подход с несколькими конусами Слепианской последовательности), что давало полосы частот шириной 20 Гц с центрами 35 Гц, 65 Гц, 85 Гц, 115 Гц и 135 Гц. Мощность рассчитывалась отдельно для длинных временных окон 250 мс от -500 мс до 1000 мс, а активность после стимула затем выражалась как относительное изменение базовой мощности, поскольку использование относительного изменения помогает преодолеть проблемы, возникающие из-за 1/ f форма данных МЭГ.
Классификация случайных лесов
Алгоритм случайного леса [24], метод ансамбля, объединяет результаты нескольких классификаторов.Эти так называемые базовые обучающиеся представляют собой деревья классификации и регрессии [28], которые разделяют данные с помощью двоичного разбиения. Целью этого процесса разделения является уменьшение примесей в отношении меток классов в дочерних узлах, которые возникают в результате этого разделения: предпочтительно, чтобы все наблюдения из одного класса приходили в один и тот же узел. При каждом разбиении алгоритм дерева сначала ищет предиктор, который максимизирует чистоту дочерних узлов, а затем лучшую точку разбиения в этом предикторе.В случайных лесах теперь растет множество деревьев; однако каждое из этих деревьев построено на выборке исходных данных, и в каждом разбиении выполняется поиск только случайной подвыборки всех предикторов. Дисперсия, вносимая этой случайностью, приводит к надежному прогнозу агрегированной модели. Кроме того, этот подход позволяет случайному лесу особенно хорошо справляться с сильно коррелированными переменными-предикторами [29], что представляет особый интерес при работе с данными MEEG. Кроме того, данные с большим количеством предикторов, чем наблюдений (маленькие задачи n , большие p ) также эффективно обрабатываются, поскольку поиск переменных-предикторов выполняется последовательно [30], что делает этот подход особенно интересным при работе с многомерными МЭЭГ в пространстве источников. данные.Для каждого предсказателя алгоритм возвращает оценку того, насколько важна эта переменная для предсказания модели. Используемая здесь версия основана на уменьшении примесей, вводимом переменной-предиктором во всех деревьях, которая измеряется индексом Джини [28, 29, 31].
Классификация случайных лесов была выполнена с использованием модуля scikit-learn для Python [32]. Целью декодирования было классифицировать испытания по модальности стимулов: зрительные или слуховые. Предикторами были тройки [воксель, момент времени, частотная полоса], что дало в целом 16 624 предиктора.Для каждого субъекта более распространенный класс (слуховая или зрительная стимуляция) был подвергнут субдискретизации, так что каждый набор данных содержал равные номера испытаний для обоих случаев. Общее количество испытаний по всем предметам составило 4270 испытаний.
Классификация была встроена в структуру перекрестной проверки по предметам: классификатор был обучен на данных от всех, кроме одного предмета, а затем протестирован на данных этого оставленного предмета. Эта процедура была повторена для всех 20 субъектов, так что каждый набор данных использовался в качестве набора для тестирования один раз.Такой подход гарантирует, что классификатор никогда не тестируется на данных, на которых он был обучен, и, таким образом, контролирует возможное переоснащение классификатора. Кроме того, он позволяет оценить предсказуемость данных в отношении переменной ответа для разных субъектов.
Каждая из 20 моделей перекрестной проверки агрегировала результаты 15 000 классификационных деревьев, где каждое дерево было построено на выборке всех наблюдений в обучающем наборе. Чтобы гарантировать, что модель включала достаточное количество деревьев, эффективность классификации была оценена с 25 000 деревьев для двух складок [33], что дало результаты, сопоставимые с более разреженной моделью.При каждом двоичном разбиении алгоритм рассматривал предикторы, чтобы найти лучшее разбиение. Точности тестовых наборов данных, а также значения переменных были объединены в свернутых сводках перекрестной проверки. Затем производительность классификатора была проверена на уровне вероятности 50% с использованием биномиального теста [34], поскольку тест, основанный на перестановках, был вычислительно невозможен.
Сравнение с опорными векторными машинами
Производительность алгоритма случайного леса сравнивалась с точностью, полученной путем подгонки двух моделей опорных векторных машин (SVM) [35, 36] к данным.Общая процедура была такой, как описано выше, вместо модели случайного леса, однако либо линейная SVM, либо нелинейная SVM с ядром радиальной базисной функции (RBF) были установлены с использованием модуля scikit-learn в Python. Параметр штрафа был установлен на 1.0 в обеих моделях, а коэффициент ядра — 1/ N features для ядра RBF.
Производительность модели случайного леса была впоследствии проверена по сравнению с двумя моделями SVM путем применения точного критерия Фишера к полученным точностям классификации.
Результаты
Чтобы оценить прогностическую ценность мощности гамма-излучения в одном исследовании в отношении модальности стимула, мы использовали данные МЭГ от 20 субъектов и приняли схему классификации между субъектами. Сначала источник данных был восстановлен с помощью формирователя луча с линейно ограниченной минимальной дисперсией (LCMV), впоследствии мы использовали алгоритм случайного леса для классификации модальности предъявления стимула (слуховой или визуальный).
Модель случайного леса классифицировала слуховые и зрительные испытания с 66.44% точности, что значительно лучше случайности (биномиальный тест, n испытаний = 4270, p <0,001). Как показывает матрица неточности на рис. 1A, точность слуховых проб была немного лучше (69,60%), чем визуальных (63,19%). В принятой 20-кратной схеме перекрестной проверки каждая кратность соответствовала данным одного субъекта, следовательно, классификатор всегда тестировался на данных одного субъекта, который не был включен в построение модели.Точность классификатора по 20 точкам перекрестной проверки изображена на рис. 1В. Эффективность различных складок различается: от 50,98% до 84,86%, однако точность для всех складок, кроме трех, превышает 60% (обратите внимание, что складки, поскольку они являются частью всей модели классификатора, не проверяются на значение). Хорошая производительность классификатора указывает на то, что модели мощности гамма-излучения являются удивительно стабильными для испытаний и даже для испытуемых. Для сравнения этого межпредметного подхода с внутрипредметным анализом см. Дополнительный рисунок в Приложении S1.
Рис. 1. Результаты классификатора.
A Матрица неточностей, показывающая долю правильно классифицированных испытаний (диагональных) и неправильно классифицированных испытаний. B Классификатор точности. Средние синие полосы представляют собой точность теста в 20 кратных точках, где каждая складка является предметом, по которому классификатор не был обучен. Темно-синяя полоса показывает общую точность классификатора, которая была проверена на уровне вероятности.
https://doi.org/10.1371/journal.pcbi.1005938.g001
Классификатор случайного леса предоставляет важность переменной в качестве оценки важности для каждого предиктора в модели. Эта мера указывает на информационную ценность данного предиктора для различения двух классов, слуховой и визуальной модальности. На рис. 2 представлены самые высокие 2% значений важности переменных, то есть те тройки [воксель, момент времени, частотная полоса], которые были наиболее информативными для разделения данных. Это ограничение было выбрано, потому что ограничения более высоких значений (> 2%) включали бы объекты с переменной важностью, равной нулю, т.е.е., переменные, которые не вносили информацию в классификатор. Не только зрительные, но и слуховые области внесли свой вклад в модель, хотя зрительные области давали больше информации, чем слуховая кора. Интересно, что нижние частотные диапазоны от 25 Гц до 45 Гц и от 55 до 75 Гц не имели такого значения, как диапазон от 75 до 95 Гц. Даже частоты выше 100 Гц внесли свой вклад в модель как в зрительной, так и в правой слуховой коре. Однако мощность гамма выше 125 Гц не внесла существенных изменений в классификационную модель.
Рис. 2. Переменные значения.
2% наиболее важных предикторов показаны во времени, частоте и пространстве. Более высокий показатель важности переменной означает, что этот предиктор имел более высокую информативную ценность в модели случайного леса для разделения данных на испытания со слуховым и визуальным восприятием. Ортогональные виды сосредоточены на вокселе, показывающем наибольшую важность переменной.
https://doi.org/10.1371/journal.pcbi.1005938.g002
Все временные окна, кроме последнего (от 750 мс до 1000 мс), поставляли информацию классификатору, на более высоких частотах казалось, что проигрываются более ранние временные окна более выраженная роль по сравнению с более низкими гамма-частотами.На рис. 3 показаны частотно-временные представления переменной важности для вокселей зрительных и слуховых пиков: воксель зрительного пика (координаты MNI: [−4 −100 12]) попадает в левую известковую борозду, воксель слухового пика (координаты MNI: [ 68 -20 10]) в правую верхнюю височную извилину (метки определены с помощью атласа автоматизированной анатомической маркировки (AAL) [37]). Частотно-временные представления этих двух пиковых вокселей (рис. 3) подтверждают закономерность, очевидную для всех вокселей (рис. 2). Таким образом, диапазон от 75 Гц до 95 Гц давал характерный и стабильный паттерн активности как в слуховой, так и в зрительной коре.Зрительный отклик, в частности, характеризовался широкополосным увеличением гаммы в диапазоне от 55 Гц до 125 Гц. Слуховой ответ имел информационную ценность в перекрывающемся, но более узком частотном диапазоне (от 75 Гц до 125 Гц).
Рис. 3. Переменные значения вокселей зрительных и слуховых пиков.
A Расположение пиков вокселей для слуховой и зрительной коры (сравните пики вокселей с рис. 2) B Частотно-временное представление переменных значений в этих пиках вокселей.Черные ящики указывают те переменные, которые входили в 2% наиболее информативных предикторов.
https://doi.org/10.1371/journal.pcbi.1005938.g003
Чтобы исследовать лежащие в основе изменения мощности гамма-излучения, ранжирование переменных по важности сравнивалось с различиями в мощности между слуховыми и зрительными пробами. С этой целью изменения слуховой и зрительной мощности относительно исходного уровня были усреднены по испытаниям и субъектам, и была вычислена разница между зрительным и слуховым состоянием.Эти различия изображены на фиг. 4A: пространственная диаграмма мощности показана для временного окна от 250 мс до 500 мс и двух частотных диапазонов (от 75 Гц до 95 Гц, вверху, и от 105 Гц до 125 Гц, внизу на фиг. 4A). Красные цвета относятся к более высокой мощности гаммы в условиях зрения, а синие цвета — к более высокой мощности в условиях слуха. Черные линии окружают те воксели, которые были среди 2% наиболее информативных предикторов для классификатора. На рис. 4В показаны лежащие в основе отношения мощности гамма-излучения для тех же пиковых вокселей, что и на рис. 3.Интересно, что анализ классификатора, основанный на единичных испытаниях, также оценил предикторы как высокоинформативные там, где разница в средних значениях мала, что наиболее очевидно для частотно-временного представления слухового состояния (от 75 Гц до 95 Гц, от 500 мс до 750 мсек). ).
Рис. 4. Базовая гамма-мощность.
На этом рисунке показана разница в усредненной мощности гаммы при визуальном и слуховом представлении слов. A Пространственное представление мощности гамма-излучения для двух частотных диапазонов (75–95 Гц, верхняя и 105–125 Гц, нижняя).Красные оттенки представляют более высокую мощность гаммы в среднем по зрительным пробам, синие цвета обозначают более высокую мощность гаммы в среднем для слухового состояния. Черные прямоугольники указывают 2% наиболее информативных предикторов, как показано на рис. 2. B Гамма-мощность в вокселях визуальных и слуховых пиков. Показана разница между визуальным и слуховым состоянием, черные ящики снова указывают на наиболее информативные предикторы для модели классификатора.
https://doi.org/10.1371/journal.pcbi.1005938.g004
Чтобы получить общую оценку относительной производительности случайных лесов на данных MEG, мы сравнили этот подход с результатами машин опорных векторов (SVM), широко используемого метода многомерного анализа данных. Мы применили два типа SVM, линейную SVM и нелинейную SVM с ядром радиальной базисной функции (RBF). Результаты этого сравнения показаны на рис. 5. Линейная SVM дала точность 63,07% и показала значительно худшие результаты, чем модель случайного леса (двусторонний точный тест Фишера, шансы , , отношение , = 86.79%, p = 0,002, 0,004 исправлено). SVM RBF показал себя немного лучше, чем модель случайного леса с точностью 68,34% (двусторонний точный тест Фишера, шансы , соотношение , , = 91,1%, p, = 0,047, 0,094 с поправкой). Обратите внимание, что сравнение между нелинейной SVM и моделью случайного леса не выдерживает поправки на множественные сравнения (поправка Бонферрони для двух точных тестов Фишера).
Рис. 5. Сравнение с моделями SVM.
Сравнение производительности алгоритма случайного леса с моделями SVM. A Результаты линейной SVM по сравнению с результатами алгоритма случайного леса в свертках перекрестной проверки. Случайный лес показал значительно лучшие результаты, чем линейный SVM ( p = 0,002, 0,004 с поправкой Бонферрони). B Результаты нелинейной SVM с ядром RBF по сравнению с результатами случайного леса. Нелинейная SVM дала более высокую точность классификации, чем модель случайного леса ( p = 0.047), применение поправки Бонферрони для множественных сравнений делает этот эффект несущественным ( p исправлено = 0,094).
https://doi.org/10.1371/journal.pcbi.1005938.g005
Обсуждение
В данной работе мы исследовали прогностическую ценность однократной гамма-мощности для классификации модальности стимулов. Это было сделано в рамках перекрестной проверки между субъектами, что позволило нам оценить не только стабильность гамма-модели в испытаниях, но и по субъектам.
Декодирование высокочастотной активности MEEG на основе однократного испытания может быть проблематичным из-за низкого SNR: в то время как интракраниально записанная высокочастотная активность до 180 Гц использовалась для декодирования движений [38, 39], подходы, сопоставимые с данными MEEG не увенчались успехом [40, 41]. Некоторые исследования могут показать вклад высокой мощности гамма-излучения (наряду с более низкой колебательной активностью) в общую производительность классификатора [42, 43]. В этом исследовании мы успешно декодировали модальность стимула исключительно из высокочастотной активности: модель классификатора смогла правильно классифицировать 66.44% испытаний, основанных на их источнике, реконструировали структуру гамма-активности, надежно отличая визуальное представление слова от слухового. Таким образом, отношение сигнал / шум гамма-мощности за одно испытание в данных МЭГ на уровне источника было достаточно высоким для успешного применения многомерного анализа за одно испытание. Интересно, что больше слуховых (69,60%), чем зрительных проб (63,19%) были классифицированы правильно, хотя зрительные области дали больше информации классификатору. Одним из возможных объяснений этого может быть то, что присущие классификатору значения отсечки мощности гамма-излучения в зрительных вокселях были довольно консервативными и, следовательно, пропускали небольшое увеличение гамма-излучения в зрительной коре при зрительных испытаниях, но все же надежно определяли отсутствие зрительной активности в слуховых испытаниях.
Классификационная модель была построена для разных субъектов с использованием 20-кратной перекрестной проверки между субъектами, при которой классификатор был обучен на 19 субъектах, а затем протестирован на данных 20 оставшихся субъектов -го . Следовательно, испытания любого конкретного субъекта были классифицированы с помощью модели, построенной на данных, полученных от разных субъектов. Используя этот подход, мы оценили общие закономерности в испытаниях и субъектах. Шаблон точности для разных складок был выше 60% для всех, кроме трех субъектов.Низкая точность указывает либо на более высокие уровни шума у этих участников, либо на модели активности, которые отклоняются от общепринятого консенсуса, выявленного с помощью модели случайного леса. Общая точность классификации 66,44% сопоставима с предыдущими отчетами о классификации данных МЭГ между субъектами (например, [44]).
Важность переменных указывает, какие предикторы использовались моделью для получения результатов классификации, обеспечивая общую картину для испытаний и субъектов, которые различали эти два состояния.Очевидно, что в модели использовалась активность гамма-диапазона как в зрительной, так и в слуховой областях, хотя зрительная кора головного мозга была более важной, чем слуховая кора, что выражалось более высокой важностью переменных ранжирования. В целом, широкий диапазон частот и временной интервал 750 мс включали активацию гамма-диапазона, соответствующую модели случайного леса.
В этом исследовании мы показываем возможность применения алгоритма случайного леса [24] к данным, локализованным по времени и частоте в одном испытании.Благодаря непараметрическому, нелинейному подходу и способности обрабатывать многомерные наборы данных с сильно коррелированными предикторами, этот метод хорошо подходит для данных MEG (см. Также [45–48]) и может обнаруживать тонкие различия, скрытые в усредненных данных. . В отличие от подходов, сравнивающих усредненные временные курсы, многомерный анализ, представленный здесь, также решает проблему множественного сравнения, которая присуща применению одномерных тестов для временных точек или аналогичных многомерных данных.
Еще одним преимуществом этого метода является возможность напрямую сравнивать предикторы (например, диапазоны частот) друг с другом относительно их важности в модели: например, мы можем утверждать, что полоса частот от 75 Гц до 95 Гц является наиболее важной. частотный диапазон, и что зрительная кора имеет более высокую информационную ценность для классификации, чем слуховая кора.
Сравнение алгоритма случайного леса с методами SVM показывает, что он явно превосходит линейную модель SVM в этом наборе данных.SVM с нелинейным ядром показала производительность, сравнимую с производительностью случайного леса, однако использование нелинейных SVM происходит за счет интерпретируемости и удобства использования. Хотя можно оценить относительную важность различных предикторов с помощью случайных лесов, это затруднено с помощью нелинейных SVM, поскольку классификация не вычисляется в исходном пространстве признаков. Более того, хотя SVM часто требует обширного поиска оптимальных настроек параметров, применение алгоритма случайного леса менее сложно и требует меньшей тонкой настройки.
По нашим данным, левая первичная зрительная кора была наиболее информативной для классификации среди всех областей мозга. Кроме того, более высокие зрительные области оцениваются как высокоинформативные, что согласуется с локализацией визуальных ответов гамма-диапазона в исследованиях внутричерепной электроэнцефалографии (iEEG) и МЭГ (например, [5, 49–51]). Далее классификация определила слуховые области как информативные. Хотя иЭЭГ надежно показывает высокие гамма-ответы на слуховые стимулы [52–55], слуховая высокочастотная активность выше 75 Гц лишь редко была показана в исследованиях МЭГ: примеры включают высокие гамма-ответы на звук и восприятие высоты звука [56, 57].В пределах слуховых областей наиболее важный воксель в наших данных был расположен в правой верхней височной извилине, что согласуется с исследованиями iEEG, изучающими фонемы и обработку текста [7, 55], и упомянутыми выше исследованиями MEG. Следующие важные области включали извилину Хешля и височную плоскость. Интересно, что правая слуховая кора показала более высокую важность с большим количеством задействованных вокселов по сравнению с левой слуховой корой, хотя стимулы были словами и обычно должны вызывать связанную с речью активность, локализованную в левом полушарии [54, 55].Это может быть объяснено тем фактом, что в обоих условиях слова использовались в качестве стимулов, и, таким образом, языковая активность левого полушария не способна различать слуховые и зрительные испытания.
Наиболее важные для классификации временные окна охватывают от 0 мс до 750 мс после начала стимула, в то время как последнее временное окно (от 750 мс до 1000 мс) не показывает каких-либо значений важности переменных высокого ранга, что означает, что гамма-активность была наиболее информативной для классификатор при предъявлении слова (среднее значение = 700 мс).
Информативные предикторы в слуховых областях, однако, были обнаружены только между 250 мс и 750 мс, хотя в предыдущих исследованиях сообщалось о ранних слуховых (высоких) гамма-ответах после фонемных или словесных стимулов (например, [7, 55, 58, 59]).
Как в зрительной, так и в слуховой областях мозга наиболее важной полосой частот была полоса от 75 Гц до 95 Гц. В частности, в окне от 250 мс до 500 мс эта полоса частот показала превышающую информативную ценность для классификации. Тем не менее, визуальные области в целом обеспечивали информативные предикторы в широком диапазоне частот (от 55 Гц до 125 Гц).Это указывает на лежащую в основе широкополосную гамма-активность в единичных испытаниях, а не на узкополосный отклик, который обычно вызывается высококонтрастными стимулами, такими как решетки (например, [5, 51]). Высокочастотная активность, полезная для классификации, аналогична визуально индуцированной широкополосной гамма-активности, описанной в исследованиях iEEG и MEG [6, 49, 60–62]. Видал и др. [61], например, описывают более низкую полосу частот от 45 Гц до 65 Гц и высокую гамма-активность от 70 Гц до 120 Гц в своем исследовании MEG по визуальной группировке.Связанная с чтением широкополосная высокочастотная активность выше 50 Гц описана в исследованиях iEEG [63–66]. Кроме того, по сравнению с узкополосными откликами, вызванными высококонтрастными стимулами, такими как решетки, которые обычно сосредоточены на более низких частотах (например, 50 Гц [5] или 60 Гц [51]), наши результаты дали частоту от 75 Гц до 95 Гц. группа как самая информативная.
В слуховой области наиболее важные переменные были сосредоточены в диапазонах частот от 75 Гц до 95 Гц и от 105 Гц до 125 Гц.Это согласуется с исследованиями iEEG по обработке слогов и текстов, которые сообщают о гамма-откликах от 80 Гц до 200 Гц [54, 67]. Таким образом, наши результаты могут отражать нижний предел высокого гамма-отклика, описанного в этих исследованиях, потенциально обрезанный низким SNR выше 125 Гц.
Подводя итог, мы показали, что однократная пробная гамма-активность может быть успешно использована для классификации модальности стимула. Важно отметить, что успешная классификация между субъектами показала, что активность гамма-диапазона в одном испытании имеет высокую согласованность между индивидуумами.В будущих исследованиях, посвященных изучению дискриминационной, а не последовательной активности в испытаниях, следует изучить возможности, предоставляемые настоящим подходом. Классификатор определил как зрительную, так и слуховую области как информативные с высокой пространственной специфичностью. Наши результаты, кроме того, предполагают, что высокочастотная активность в однократном испытании после визуального представления слова характеризуется скорее широкополосным, чем узкополосным ответом.
Дополнительная информация
S1 Приложение. Внутрипредметная классификация.
Точности классификатора, полученные при обучении и тестировании модели случайного леса внутри, а не между предметами. В этой процедуре была принята схема перекрестной проверки с разделением половин, включающая как настройку формирователя луча, так и моделирование классификации.
https://doi.org/10.1371/journal.pcbi.1005938.s001
(PDF)
Благодарности
Мы благодарим Анн-Кристин Ромбах, Леону Хеллвиг, Марину Кёпфер и Джанин Вайхерт за помощь в сборе данных, а также Сабину Леске и Цветана Попова за ценные обсуждения.
Ссылки
- 1. Грей С.М., Сингер В. Стимул-специфические нейронные колебания в ориентационных столбцах зрительной коры головного мозга кошек. Труды Национальной академии наук. 1989; 86: 1698–1702.
- 2. Gray CM, König P, Engel AK, Singer W. Колебательные реакции в зрительной коре головного мозга кошек демонстрируют межколоночную синхронизацию, которая отражает глобальные свойства стимула. Природа. 1989; 338: 334–337. pmid: 2922061
- 3. Hoogenboom N, Schoffelen JM, Oostenveld R, Parkes LM, Fries P.Локализация визуальной активности человека в гамма-диапазоне по частоте, времени и пространству. NeuroImage. 2006. 29 (3): 764–773. pmid: 16216533
- 4. Далал С.С., Видаль Дж. Р., Хамаме С. М., Оссандон Т., Бертран О., Лашо Дж. П. и др. Охватывает богатый спектр человеческого мозга: от медленных волн до гаммы и выше. Структура и функции мозга. 2011; 216: 77–84. pmid: 21437655
- 5. Muthukumaraswamy SD, Singh KD, Swettenham JB, Jones DK. Визуальные гамма-колебания и вызванные ответы: изменчивость, повторяемость и структурная корреляция МРТ.NeuroImage. 2010. 49 (4): 3349–3357. pmid: 19944770
- 6. Гермес Д., Миллер К.Дж., Ванделл Б.А., Винавер Дж. Стимул-зависимость гамма-колебаний в зрительной коре головного мозга человека. Кора головного мозга. 2014. 25 (9): 2951–2959. pmid: 24855114
- 7. Крон Н.Э., Боутман Д., Гордон Б., Хао Л. Индуцированная электрокортикографическая гамма-активность во время слухового восприятия. Клиническая нейрофизиология. 2001; 112: 565–582. pmid: 11275528
- 8. Brosch M, Budinger E, Scheich H.Связанные со стимулами гамма-колебания в слуховой коре приматов. Журнал нейрофизиологии. 2002. 87 (6): 2715–2725. pmid: 12037173
- 9. Фелл Дж., Клавер П., Ленертц К., Грюнвальд Т., Шаллер С., Элгер К.Э. и др. Формирование памяти человека сопровождается ринально-гиппокампальной связью и разделением. Природа Неврологии. 2001; 4: 1259–1264. pmid: 11694886
- 10. Осипова Д., Такашима А., Остенвельд Р., Фернандес Г., Марис Э., Дженсен О. Тета и гамма-колебания предсказывают кодирование и извлечение декларативной памяти.Журнал неврологии. 2006. 26 (28): 7523–7531. pmid: 16837600
- 11. Дженсен О, Кайзер Дж, Лашо Дж. П. Колебания гамма-частоты человека, связанные с вниманием и памятью. Тенденции в неврологии. 2007. 30: 317–324. pmid: 17499860
- 12. Staudigl T, Hanslmayr S. Колебания теты при кодировании опосредуют контекстно-зависимую природу эпизодической памяти человека. Текущая биология. 2013. 23 (12): 1101–1106. pmid: 23746635
- 13. Акам Т, Куллманн ДМ.Колебательное мультиплексирование популяционных кодов для избирательной коммуникации в мозге млекопитающих. Обзоры природы Неврология. 2014; 15: 111–122. pmid: 24434912
- 14. Дженсен О., Мазахери А. Формирование функциональной архитектуры за счет колебательной альфа-активности: стробирование путем торможения. Границы неврологии человека. 2010; 4 (186).
- 15. Фрайз П. Ритмы познания: общение через связность. Нейрон. 2015; 88 (1): 220–235. pmid: 26447583
- 16. Миллер KJ, Соренсен LB, Ojemann JG, Den Nijs M.Масштабирование по степенному закону электрического потенциала поверхности мозга. Вычислительная биология PLoS. 2009; 5 (12).
- 17. Рэй С., Маунселл Дж. Х. Различия в гамма-частотах зрительной коры ограничивают их возможное использование в вычислениях. Нейрон. 2010. 67 (5): 885–896. pmid: 20826318
- 18. Стоукс М., Спаак Э. Важность однократного анализа в когнитивной нейробиологии. Тенденции в когнитивных науках. 2016; 20 (7): 483–486. pmid: 27237797
- 19. Ловет Э., Робертс М.Дж., Босман, Калифорния, Фрайс П., Де Верд П.Области V1 и V2 показывают связанные с микросаккадом 3-4 Гц ковариации мощности и частоты гамма-излучения. Европейский журнал нейробиологии. 2016; 43: 1286–1296. pmid: 26547390
- 20. Лундквист М., Роуз Дж., Герман П., Бринкат С.Л., Бушман Т.Дж., Миллер Е.К. В основе рабочей памяти лежат гамма- и бета-всплески. Нейрон. 2016; 90: 152–164. pmid: 26996084
- 21. Guimaraes MP, Wong DK, Uy ET, Grosenick L, Suppes P. Классификация записей MEG за один раз. IEEE Transactions по биомедицинской инженерии.2007. 54 (3): 436–443. pmid: 17355055
- 22. Грамфорт А., Луесси М., Ларсон Э., Энгеманн Д.А., Стромайер Д., Бродбек С. и др. Анализ данных МЭГ и ЭЭГ с помощью MNE-Python. Границы неврологии. 2013; 7: 267. pmid: 24431986
- 23. Ван Вин Б.Д., ван Дронгелен В., Юхтман М., Сузуки А. Локализация электрической активности мозга с помощью линейно ограниченной пространственной фильтрации с минимальной дисперсией. Транзакция IEEE по биомедицинской инженерии. 1997. 44 (9): 867–880.
- 24.Брейман Л. Случайные леса. Машинное обучение. 2001. 45 (1): 5–32.
- 25. Остенвельд Р., Фрис П., Марис Э., Шоффелен Дж. М.. FieldTrip: программное обеспечение с открытым исходным кодом для расширенного анализа данных МЭГ, ЭЭГ и инвазивных электрофизиологических данных. Вычислительный интеллект и нейробиология. 2011.
- 26. Секихара К., Нагараджан СС. Адаптивные пространственные фильтры для электромагнитной визуализации мозга. Springer Science & Business Media; 2008.
- 27. Нольте Г.Теорема о магнитном поле свинца в квазистатическом приближении и ее использование для прямого расчета магнитоэнцефалографии в реалистичных объемных проводниках. Физика в медицине и биологии. 2003. 48 (22): 3637–3652. pmid: 14680264
- 28. Брейман Л., Фридман Дж., Стоун С.Дж., Ольшен Р.А. Деревья классификации и регрессии. CRC пресс; 1984.
- 29. Катлер А., Катлер Д. Р., Стивенс-младший. Древовидные методы. В: Анализ многомерных данных в исследованиях рака. Springer New York; 2009 г.п. 1–19.
- 30. Штробл С., Малли Дж., Тутц Г. Введение в рекурсивное разбиение: обоснование, применение и характеристики деревьев классификации и регрессии, мешков и случайных лесов. Психологические методы. 2009; 14 (4): 323. pmid: 19968396
- 31. Джини С. Вариабитта и взаимообусловленность: Contributo allo Studio delle Distribuzioni e delle Relazioni Statistiche. Tipografia di Cuppini; 1912.
- 32. Pedregosa F, Varoquaux G, Gramfort A, Michel V, Thirion B, Grisel O и др.Scikit-learn: Машинное обучение на Python. Журнал исследований в области машинного обучения. 2011; 12: 2825–2830.
- 33. Лиау А., Винер М. Классификация и регрессия методом randomForest. Новости R. 2002. 2 (3): 18–22.
- 34. Комбриссон Э., Джерби К. Случайное превышение уровня шанса: оговорка о теоретических уровнях шанса в классификации сигналов мозга и статистической оценке точности декодирования. Журнал методов неврологии. 2015; 30: 126–136.
- 35. Босер Б.Е., Гийон И.М., Вапник В.Н.Алгоритм обучения для классификаторов оптимальной маржи. В: Материалы пятого ежегодного семинара по теории вычислительного обучения. ACM; 1992. стр. 144–152.
- 36. Кортес Ч., Вапник В. Сети опорных векторов. Машинное обучение. 1995. 20 (3): 273–297.
- 37. Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, Delcroix N и др. Автоматическая анатомическая маркировка активаций в SPM с использованием макроскопической анатомической парцелляции головного мозга одного пациента MNI MRI.NeuroImage. 2002. 15: 273–289. pmid: 11771995
- 38. Leuthardt EC, Schalk G, Wolpaw JR, Ojemann JG, Moran DW. Интерфейс мозг-компьютер, использующий электрокортикографические сигналы у людей. Журнал нейронной инженерии. 2004; 1 (2): 63. pmid: 15876624
- 39. Rickert J, de Oliveira SC, Vaadia E, Aertsen A, Rotter S, Mehring C. Кодирование направления движения в различных частотных диапазонах потенциалов локального поля моторной коры. Журнал неврологии. 2005. 25 (39): 8815–8824.pmid: 16192371
- 40. Уолдерт С., Прейссл Х., Демандт Э., Браун С., Бирбаумер Н., Аэрцен А. и др. Направление движения руки расшифровывается по данным МЭГ и ЭЭГ. Журнал неврологии. 2008. 28 (4): 1000–1008. pmid: 18216207
- 41. Quandt F, Reichert C, Hinrichs H, Heinze HJ, Knight RT, Rieger JW. Единичное испытание различения движений отдельных пальцев на одной руке: комбинированное исследование МЭГ и ЭЭГ. NeuroImage. 2012. 59 (4): 3316–3324. pmid: 22155040
- 42. Fuentemilla L, Penny WD, Cashdollar N, Bunzeck N, Düzel E.Периодическое воспроизведение в рабочей памяти с тета-связью. Текущая биология. 2010. 20 (7): 606–612. pmid: 20303266
- 43. Шульц Э., Жердин А., Тиманн Л., Плант С, Плонер М. Расшифровка чувствительности человека к боли на основе многомерного анализа данных ЭЭГ. Кора головного мозга. 2011. 22 (5): 1118–1123. pmid: 21765182
- 44. Оливетти Э., Киа С.М., Авесани П. Декодирование MEG по предметам. В: Распознавание образов в нейровизуализации, Международный семинар 2014 г. IEEE; 2014 г.п. 1–4.
- 45. Фрайван Л., Лвизи К., Хасауна Н., Венц Х., Дикхаус Х. Автоматическая система идентификации стадий сна на основе частотно-временного анализа одного канала ЭЭГ и случайного лесного классификатора. Компьютерные методы и программы в биомедицине. 2012; 108 (1): 10–19. pmid: 22178068
- 46. Леманн С., Кениг Т., Джелич В., Причеп Л., Джон Р. Э., Валунд Л. О. и др. Применение и сравнение алгоритмов классификации для распознавания болезни Альцгеймера по электрической активности мозга (ЭЭГ).Журнал методов неврологии. 2007. 161 (2): 342–350. pmid: 17156848
- 47. Бентлемсан М., Земури Е.Т., Бухаффра Д., Яхья-Зубир Б., Ферруджи К. Случайный лес и общие пространственные паттерны банка фильтров для классификации двигательных образов на основе ЭЭГ. В: 5-я Международная конференция по интеллектуальным системам, моделированию и имитационному моделированию (СМИБ). IEEE; 2014. с. 235–238.
- 48. Donos C, Dümpelmann M, Schulze-Bonhage A. Алгоритм раннего обнаружения приступов, основанный на внутричерепной ЭЭГ и классификации случайных лесов.Международный журнал нейронных систем. 2015; 25 (05): 1550023. pmid: 26022388
- 49. Таллон-Бодри C, Бертран O, Hénaff MA, Isnard J, Fischer C. Внимание по-разному модулирует колебания гамма-диапазона в боковой затылочной коре и веретенообразной извилине человека. Кора головного мозга. 2005. 15 (5): 654–662. pmid: 15371290
- 50. Swettenham JB, Muthukumaraswamy SD, Singh KD. Спектральные свойства индуцированных и вызванных гамма-колебаний ранней зрительной коры головного мозга человека на движущиеся и неподвижные раздражители.Журнал нейрофизиологии. 2009. 102 (2): 1241–1253. pmid: 19515947
- 51. Тан Х.Р., Гросс Дж., Улхас П. Измерения датчика и источника визуально индуцированных колебаний гамма-диапазона очень надежны. NeuroImage. 2016; 137: 34–44. pmid: 27153980
- 52. Эдвардс Э., Солтани М., Деуэлл Л.Й., Бергер М.С., Найт РТ. Высокая гамма-активность в ответ на отклоняющиеся слуховые стимулы, записанные непосредственно из коры головного мозга человека. Журнал нейрофизиологии. 2005. 94 (6): 4269–4280. pmid: 16093343
- 53.Биде-Кооле А., Фишер С., Баучет Ф., Агера П. Е., Бертран О. Нейронный субстрат одновременного восприятия звука: прямые электрофизиологические записи из слуховой коры головного мозга человека. Границы неврологии человека. 2007; 1. pmid: 18958219
- 54. Канолти Р.Т., Солтани М., Далал С.С., Эдвардс Э., Дронкерс Н.Ф., Нагараджан С.С. и др. Пространственно-временная динамика обработки текста в мозгу человека. Границы неврологии. 2007; 1:14.
- 55. Эдвардс Э., Солтани М., Ким В., Далал С.С., Нагараджан С.С., Бергер М.С. и др.Сравнение времени — частотные характеристики и связанный с событием потенциал звуковых речевых стимулов в коре головного мозга человека. Журнал нейрофизиологии. 2009. 102 (1): 377–386. pmid: 19439673
- 56. Schepers IM, Hipp JF, Schneider TR, Röder B, Engel AK. Функционально специфическая колебательная активность коррелирует между зрительной и слуховой корой у слепых. Головной мозг. 2012. 135 (3): 922–934. pmid: 22366801
- 57. Седли В., Теки С., Кумар С., Оверат Т., Барнс Г. Р., Гриффитс Т. Д..Отклики высоты тона гамма-диапазона в слуховой коре человека, измеренные с помощью магнитоэнцефалографии. NeuroImage. 2012; 59 (2): 1904–1911. pmid: 21925281
- 58. Эдвардс Э., Нагараджан С.С., Далал С.С., Канолти Р.Т., Кирш Х.Э., Барбаро Н.М. и др. Пространственно-временная визуализация корковой активации во время генерации глаголов и именования картинок. NeuroImage. 2010. 50 (1): 291–301. pmid: 20026224
- 59. Штейншнайдер М., Нурски К.В., Кавасаки Х., Оя Х., Брюгге Дж. Ф., Ховард М.А. Внутричерепное исследование речевой активности заднебоковой верхней височной извилины.Кора головного мозга. 2011. 21 (10): 2332–2347. pmid: 21368087
- 60. Lachaux JP, George N, Tallon-Baudry C, Martinerie J, Hugueville L, Minotti L, et al. Многоликая реакция гамма-диапазона на сложные визуальные раздражители. NeuroImage. 2005. 25 (2): 491–501. pmid: 15784428
- 61. Видал Дж. Р., Чаумон М., О’Реган Дж. К., Таллон-Бодри С. Визуальная группировка и фокусировка внимания вызывают колебания гамма-диапазона на разных частотах в сигналах магнитоэнцефалограммы человека.Журнал когнитивной неврологии. 2006. 18 (11): 1850–1862. pmid: 17069476
- 62. Сигель М., Доннер Т.Х., Остенвельд Р., Фрис П., Энгель А.К. Высокочастотная активность зрительной коры головного мозга человека модулируется силой зрительного движения. Кора головного мозга. 2007. 17 (3): 732–741. pmid: 16648451
- 63. Юнг Дж., Мэйни Н., Кахане П., Минотти Л., Хоффманн Д., Бертран О. и др. Нейронные основы внимательного чтения. Картирование человеческого мозга. 2008. 29 (10): 1193–1206. pmid: 17894399
- 64.Далал С.С., Байлет С., Адам С., Дукорпс А., Шварц Д., Джерби К. и др. Одновременная запись МЭГ и внутричерепной ЭЭГ при внимательном чтении. NeuroImage. 2009. 45 (4): 1289–1304. pmid: 19349241
- 65. Хамаме С.М., Видал Дж. Р., Оссандон Т., Джерби К., Далал С.С., Минотти Л. и др. Чтение мысленным взором: онлайн-определение зрительно-пространственной рабочей памяти и визуальных образов в нижней височной доле. NeuroImage. 2012; 59 (1): 872–879. pmid: 21839843
- 66. Хамаме С.М., Свед М., Шарман М., Видал Дж. Р., Перроне-Бертолотти М., Кахане П. и др.Зона чтения Дежерина повторно с помощью внутричерепной ЭЭГ: избирательные реакции на цепочки букв. Неврология. 2013. 80 (6): 602–603. pmid: 23382370
- 67. Чанг Э.Ф., Эдвардс Э., Нагараджан С.С., Фогельсон Н., Далал С.С., Канолти Р.Т. и др. Корковая пространственно-временная динамика, лежащая в основе обнаружения фонологической цели у человека. Журнал когнитивной неврологии. 2011. 23 (6): 1437–1446. pmid: 20465359
Классификация модальности стимула от высокочастотной активности МЭГ человека между субъектами
Реферат
Анализ отдельных испытаний может выявить значимую динамику мозга, которая не видна при усреднении по испытаниям.Однако низкое отношение сигнал / шум (SNR) может препятствовать использованию методов однократного анализа и декодирования. В этом исследовании мы исследуем применимость однократного подхода к декодированию модальности стимула из высокочастотной активности магнитоэнцефалографии (МЭГ). Чтобы классифицировать слуховое и визуальное представление слов, мы объединяем реконструкцию источника луча с методом случайной классификации лесов. Чтобы сделать вывод на групповом уровне, классификация встроена во внутрипредметную структуру.
Мы показываем, что SNR гамма-излучения за одно испытание обеспечивает хорошую эффективность классификации (точность по предметам: 66,44%). Это означает, что характеристики высокочастотной активности имеют высокую согласованность между испытаниями и субъектами. Классификатор случайного леса присвоил информационную ценность активности как слуховой, так и зрительной коры с высокой пространственной специфичностью. С течением времени мощность гаммы была наиболее информативной во время предъявления стимула. Среди всех частотных диапазонов полоса 75-95 Гц была наиболее информативной частотной полосой как в зрительной, так и в слуховой областях.Широкий диапазон гамма-частот (55-125 Гц), особенно в визуальных областях, способствовал успешной классификации.
Таким образом, мы демонстрируем возможность однократных пробных подходов для декодирования модальности стимула у субъектов от высокочастотной активности и описываем различительную гамма-активность во времени, частоте и пространстве.
Сводка по автору Усреднение мозговой активности по испытаниям — мощный способ повысить отношение сигнал / шум в данных МЭГ.Этот подход, однако, потенциально скрывает значимую динамику мозга, которая разворачивается на уровне одного исследования. Единичные пробные анализы были успешно применены к временной области или низкочастотной колебательной активности; его применение к высокочастотной активности МЭГ затруднено из-за низкой амплитуды этих сигналов. В настоящем исследовании мы показываем, что модальность стимула (визуальное или слуховое представление слов) может быть успешно декодирована из однократной высокочастотной активности MEG путем комбинирования реконструкции источника с алгоритмом классификации случайного леса.Этот подход позволяет выявить закономерности активности выше 75 Гц как в зрительной, так и в слуховой коре, подчеркивая важность высокочастотной активности для обработки доменно-специфических стимулов. Таким образом, наши результаты расширяют предыдущие выводы, выявляя высокочастотную активность в слуховой коре, связанную со слуховыми словесными стимулами в данных МЭГ. Кроме того, принятая межпредметная структура предполагает высокую согласованность между индивидуумами в высокочастотных моделях активности.
Введение
С тех пор, как появились первые сообщения об активности кортикального гамма-диапазона [1, 2], эти высокочастотные отклики были связаны с множеством мозговых процессов и умственных задач, например, зрительного восприятия и обработки [3, 4, 5, 6], слуховое восприятие [7, 8] или память [9, 10, 11, 12].Хотя были предложены многочисленные теории о происхождении и функции этих высокочастотных колебаний и их связи с более низкими частотами, такими как тета и альфа (например, [13, 14, 15]), продолжаются споры о том, отражают ли ответы гамма-диапазона узкополосный колебания или широкополосная мощность увеличивается, что может отражать увеличение пиковой активности [16, 17, 6]. Одним из препятствий в этом поиске является характеристика 1 / f частотного спектра мощности мозга и низкое отношение сигнал / шум (SNR) активности гамма-диапазона в записях МЭГ или электроэнцефалографии (ЭЭГ).Чтобы увеличить SNR, часто используется пробное усреднение для устранения случайной дисперсии. Однако этот подход потенциально может скрыть или отменить значимую активность мозга [18]. В самом деле, потенциалы локального поля и данные электрокортикографии обезьян выявили систематические изменения мощности и частоты гамма-излучения от опыта к опыту в зрительном [19] и в задаче памяти [20]. Важно отметить, что средние значения по испытаниям в этих исследованиях демонстрируют классический устойчивый гамма-эффект, указывая на то, что ответы от одного испытания имеют решающее значение для понимания динамики мозга [18].Одним из эффективных подходов к оценке информации, полученной в результате однократного испытания, являются методы многомерного декодирования. Однако остается неясным, применимы ли такие методы к данным МЭГ в гамма-диапазоне с низким отношением сигнал / шум. В настоящей статье мы исследуем прогностическую ценность мощности гамма-излучения при однократном испытании в отношении модальности предъявления стимула (слуховое или визуальное представление слов) в данных МЭГ человека. Хотя сопоставимые контрасты использовались для тестирования производительности классификатора или в качестве примеров наборов данных (например, [21, 22]), нашей целью было раскрыть однократные высокочастотные паттерны в данных МЭГ человека.Чтобы декодировать информацию о стимуле-модальности из частотно-временных данных, мы использовали комбинацию формирования диаграммы направленности [23] и классификации случайного леса [24]. Этот подход был встроен в структуру перекрестной проверки между предметами, где классификатор был протестирован на невидимых предметах, чтобы оценить общность пространственного частотно-временного паттерна. Наши результаты подтверждают, что гамма-отношение сигнал / шум в единичных испытаниях достаточно высоко для достижения стабильной точности классификации, значительно превышающей вероятность. Интересно, что модель классификации придает высокую информационную ценность широкой полосе пропускания в гамма-диапазоне.Кроме того, мы показываем, что характеристики гамма-активности достаточно схожи в разных испытаниях и даже у испытуемых, чтобы обеспечить надежную классификацию.
Результаты
Чтобы оценить прогностическую ценность мощности гамма-излучения в одном исследовании в отношении модальности стимула, мы использовали данные МЭГ от 20 субъектов и приняли схему классификации между субъектами. Сначала источник данных был восстановлен с помощью формирователя луча с линейно ограниченной минимальной дисперсией (LCMV), впоследствии мы использовали алгоритм случайного леса для классификации модальности предъявления стимула (слуховой или визуальный).
Модель случайного леса классифицирует слуховые и зрительные испытания с точностью 66,44%, что значительно лучше случайности (биномиальный тест, n испытаний = 4270, p <0,001). Как показывает матрица путаницы на Рисунке 1A, точность слуховых проб была немного лучше (69,60%), чем визуальных проб (63,19%). В принятой 20-кратной схеме перекрестной проверки каждая кратность соответствовала данным одного субъекта, следовательно, классификатор всегда тестировался на данных одного субъекта, который не был включен в построение модели.Точность классификатора по 20 точкам перекрестной проверки изображена на рисунке 1B. Эффективность различных складок варьируется от 50,98 до 84,86%, однако точность для всех складок, кроме трех, превышает 60% (обратите внимание, что складки, поскольку они являются частью всей модели классификатора, не проверяются на значимость. ). Хорошая производительность классификатора указывает на то, что модели мощности гамма-излучения являются удивительно стабильными для испытаний и даже для испытуемых.
Рисунок 1: Результаты классификатора.A Матрица неточностей, показывающая долю правильно классифицированных испытаний (диагональ) и неправильно классифицированных испытаний. B Классификатор точности. Средние синие полосы представляют собой точность теста в 20 кратных точках, где каждая складка является предметом, по которому классификатор не был обучен. Темно-синяя полоса показывает общую точность классификатора, которая была проверена на уровне вероятности.
Классификатор случайного леса предоставляет важность переменной в качестве оценки важности для каждого предиктора в модели. Эта мера указывает на информационную ценность данного предиктора для различения двух классов, слуховой и визуальной модальности.На рисунке 2 показаны 2% самых высоких значений важности переменных, то есть те тройки [воксель, момент времени, частотная полоса], которые были наиболее информативными для разделения данных. Не только зрительные, но и слуховые области внесли свой вклад в модель, хотя зрительные области давали больше информации, чем слуховая кора. Интересно, что нижние частотные диапазоны 25-45 Гц и 55-75 Гц не имели такого значения, как диапазон 75-95 Гц. Даже частоты выше 100 Гц внесли свой вклад в модель как в зрительной, так и в правой слуховой коре.Однако мощность гамма выше 125 Гц не внесла существенных изменений в классификационную модель.
Рисунок 2: Переменные значения.2% наиболее важных предикторов показаны во времени, частоте и пространстве. Более высокий показатель важности переменной означает, что этот предиктор имел более высокую информативную ценность в модели случайного леса для разделения данных на испытания со слуховым и визуальным восприятием. Ортогональные виды сосредоточены на вокселе, показывающем наибольшую важность переменной.
Все временные окна, кроме последнего (750-1000 мс), поставляли информацию классификатору, на более высоких частотах более ранние временные окна, казалось, играли более явную роль по сравнению с более низкими гамма-частотами.На рисунке 3 показаны частотно-временные представления переменной важности для вокселей зрительного и слухового пиков: воксель зрительного пика (координаты MNI: [−4 −100 12]) попадает в левую известковую борозду, воксель слухового пика (координаты MNI: [ 68 -20 10]) в правую верхнюю височную извилину (метки определены с помощью атласа автоматизированной анатомической маркировки (AAL) [25]). Частотно-временные представления этих двух пиковых вокселей (рисунок 3) подтверждают закономерность, очевидную для всех вокселей (рисунок 2). Таким образом, полоса 75-95 Гц давала характерный и стабильный паттерн активности как в слуховой, так и в зрительной коре.Зрительный отклик, в частности, характеризовался широкополосным увеличением гаммы в диапазоне от 55 до 125 Гц. Слуховой ответ имел информативную ценность в перекрывающемся, но более узком частотном диапазоне (75-125 Гц).
Рисунок 3: Переменные значения вокселей зрительных и слуховых пиков.A Расположение пиков вокселей для слуховой и зрительной коры (сравните пики вокселей с рис. 2) B Частотно-временное представление переменных значений в этих пиках вокселей. Черные ящики указывают те переменные, которые входили в 2% наиболее информативных предикторов.
Чтобы исследовать лежащие в основе изменения мощности гамма, ранжирование переменных по важности сравнивалось с разницей в мощности между слуховыми и зрительными пробами. С этой целью изменения слуховой и зрительной мощности относительно исходного уровня были усреднены по испытаниям и субъектам, и была вычислена разница между зрительным и слуховым состоянием. Эти различия показаны на рисунке 4A: пространственная диаграмма мощности показана для временного окна 250-500 мс и двух полос частот (75-95 Гц, вверху, и 105-125 Гц, внизу на рисунке 4A).Красные цвета относятся к более высокой мощности гаммы в условиях зрения, а синие цвета — к более высокой мощности в условиях слуха. Черные линии окружают те воксели, которые были среди 2% наиболее информативных предикторов для классификатора. На рисунке 4B показаны лежащие в основе отношения мощности гамма-излучения для тех же пиковых вокселей, что и на рисунке 3. Интересно, что анализ классификатора, основанный на единичных испытаниях, также оценил предикторы как высокоинформативные там, где разница в средних значениях мала, что наиболее очевидно для того времени. -частотное представление состояния слуха (75-95 Гц, 500-750 мс).
Рисунок 4: Базовая гамма-мощность.На этом рисунке показана разница в усредненной мощности гаммы при визуальном и слуховом представлении слов. A Пространственное представление мощности гамма-излучения для двух частотных диапазонов (75–95 Гц, верхняя и 105–125 Гц, нижняя). Красные оттенки представляют более высокую мощность гаммы в среднем по зрительным пробам, синие цвета обозначают более высокую мощность гаммы в среднем для слухового состояния. Черные прямоугольники указывают 2% наиболее информативных предикторов, как показано на рисунке 2. B Мощность гаммы в вокселях визуальных и слуховых пиков. Показана разница между визуальным и слуховым состоянием, черные ящики снова указывают на наиболее информативные предикторы для модели классификатора.
Обсуждение
В данной работе мы исследовали прогностическую ценность однократной гамма-мощности для классификации модальности стимулов. Это было сделано в рамках перекрестной проверки между субъектами, что позволило нам оценить не только стабильность гамма-модели в испытаниях, но и по субъектам.
Декодирование высокочастотной активности MEEG на основе однократного испытания может быть проблематичным из-за низкого отношения сигнал / шум: в то время как интракраниально записанная высокочастотная активность до 180 Гц использовалась для декодирования движений [26, 27], подходы, сравнимые с MEEG данные не были успешными [28, 29]. Некоторые исследования могут показать вклад высокой мощности гамма-излучения (наряду с более низкой колебательной активностью) в общую производительность классификатора [30, 31]. В этом исследовании мы успешно декодировали модальность стимула исключительно из высокочастотной активности: модель классификатора смогла правильно классифицировать 66.44% испытаний, основанных на их источнике, реконструировали структуру гамма-активности, надежно отличая визуальное представление слова от слухового. Таким образом, отношение сигнал / шум гамма-мощности за одно испытание в данных МЭГ на уровне источника было достаточно высоким для успешного применения многомерного анализа за одно испытание. Интересно, что больше слуховых (69,30%), чем зрительных проб (63,19%) были классифицированы правильно, хотя зрительные области дали больше информации классификатору. Одним из возможных объяснений этого может быть то, что присущие классификатору значения отсечки мощности гамма-излучения в зрительных вокселях были довольно консервативными и, следовательно, пропускали небольшое увеличение гамма-излучения в зрительной коре при зрительных испытаниях, но все же надежно определяли отсутствие зрительной активности в слуховых испытаниях.
Классификационная модель была построена по субъектам с использованием 20-кратной перекрестной проверки по субъектам, при которой классификатор был обучен на 19 субъектах, а затем протестирован на данных 20 оставшихся субъектов -го . Следовательно, испытания любого конкретного субъекта были классифицированы с помощью модели, построенной на данных, полученных от разных субъектов. Используя этот подход, мы оценили общие закономерности в испытаниях и субъектах. Шаблон точности для разных складок был выше 60% для всех субъектов, кроме трех.Низкая точность указывает либо на более высокие уровни шума у этих участников, либо на модели активности, которые отклоняются от общепринятого консенсуса, выявленного с помощью модели случайного леса. Общая точность классификации 66,44% сопоставима с предыдущими отчетами о классификации данных МЭГ между субъектами (например, [32]).
Важность переменных указывает, какие предикторы использовались моделью для получения результатов классификации, обеспечивая общую картину испытаний и субъектов, которые различали эти два состояния.Очевидно, что в модели использовалась активность гамма-диапазона как в зрительной, так и в слуховой областях, хотя зрительная кора головного мозга была более важной, чем слуховая кора, что выражалось более высокой важностью переменных ранжирования. В целом, широкий диапазон частот и временной интервал 750 мс включали активацию гамма-диапазона, соответствующую модели случайного леса.
В этом исследовании мы показываем возможность применения алгоритма случайного леса [24] к локализованным временным и частотным данным в одном испытании. Благодаря непараметрическому, нелинейному подходу и способности обрабатывать многомерные наборы данных с сильно коррелированными предикторами, этот метод хорошо подходит для данных MEG (см. Также [33, 34, 35, 36]) и может обнаруживать тонкие скрытые различия. в усредненных данных.
Еще одним преимуществом этого метода является возможность прямого сравнения предикторов. (например, полосы частот) друг другу относительно их важности в модели: например, мы можем заявить, что полоса частот 75-95 Гц является наиболее важной полосой частот, и что зрительная кора имеет более высокую информационную ценность для классификации, чем слуховая кора.
По нашим данным, левая первичная зрительная кора была наиболее информативной для классификации среди всех областей мозга.Кроме того, более высокие зрительные области оцениваются как высокоинформативные, что согласуется с локализацией визуальных ответов гамма-диапазона в исследованиях внутричерепной электроэнцефалографии (iEEG) и МЭГ (например, [37, 38, 5, 39]). Далее классификация определила слуховые области как информативные. Хотя иЭЭГ надежно показывает высокие гамма-ответы на слуховые стимулы [40, 41, 42, 43], слуховая высокочастотная активность выше 75 Гц лишь редко была показана в исследованиях МЭГ: примеры включают высокие гамма-ответы на звук и восприятие высоты звука [44, 45 ].В пределах слуховых областей наиболее важный воксель в наших данных был расположен в правой верхней височной извилине, что согласуется с исследованиями iEEG, изучающими фонемы и обработку текста [7, 43], и упомянутыми выше исследованиями MEG. Следующие важные области включали извилину Хешля и височную плоскость. Интересно, что правая слуховая кора показала более высокую важность с большим количеством вовлеченных вокселов по сравнению с левой слуховой корой, хотя стимулы были словами и обычно должны вызывать связанную с языком активность, локализованную в левом полушарии [42, 43].Это может быть объяснено тем фактом, что в обоих условиях слова использовались в качестве стимулов, и, таким образом, языковая активность левого полушария не способна различать слуховые и зрительные испытания.
Временные окна, наиболее важные для классификации, охватывали от 0 до 750 мс после начала стимула, в то время как последнее временное окно (750-1000 мс) не показывало никаких значений важности переменных высокого ранга, что означает, что гамма-активность была наиболее информативной для классификатора. во время предъявления слова (среднее значение = 700 мс).
Информативные предикторы в слуховых областях, однако, были обнаружены только между 250 и 750 мс, хотя в предыдущих исследованиях сообщалось о ранних слуховых (высоких) гамма-ответах после фонемных или словесных стимулов (например, [7, 43, 46, 47]).
Как в зрительной, так и в слуховой областях мозга наиболее важной полосой частот была полоса 75–95 Гц. Эта полоса частот, особенно в окне 250-500 мс, демонстрирует чрезмерно информативную ценность для классификации. Тем не менее, визуальные области в целом обеспечивали информативные предикторы в широком диапазоне частот (55-125 Гц).Это указывает на лежащую в основе широкополосную гамма-активность в единичных испытаниях, а не на узкополосный отклик, который обычно вызывается высококонтрастными стимулами, такими как решетки (например, [5, 39]). Высокочастотная активность, полезная для классификации, аналогична визуально индуцированной широкополосной гамма-активности, описанной в исследованиях iEEG и MEG [48, 37, 49, 50, 6]. Видал и др. [49], например, описывают более низкую полосу частот 45-65 Гц и высокую гамма-активность 70-120 Гц в своем исследовании MEG по визуальной группировке.Связанная с чтением широкополосная высокочастотная активность выше 50 Гц описана в исследованиях iEEG [51, 52, 53, 54]. Кроме того, по сравнению с узкополосными откликами, вызванными высококонтрастными стимулами, такими как решетки, которые обычно сосредоточены на более низких частотах (например, 50 Гц [5] или 60 Гц [39]), наши результаты дали полосу частот 75-95 Гц как наиболее информативный.
В слуховой области наиболее важные переменные были сосредоточены в полосах частот 75–95 Гц и 105–125 Гц Гц.Это согласуется с исследованиями iEEG по обработке слогов и текстов, которые сообщают о гамма-откликах от 80 Гц до 200 Гц [42, 55]. Таким образом, наши результаты могут отражать нижний предел высокого гамма-отклика, описанного в этих исследованиях, потенциально обрезанный низким SNR выше 125 Гц.
Подводя итог, мы показали, что однократная пробная гамма-активность может быть успешно использована для классификации модальности стимула. Важно отметить, что успешная классификация между субъектами показала, что активность гамма-диапазона в одном испытании имеет высокую согласованность между индивидуумами.Классификатор определил как зрительную, так и слуховую области как информативные с высокой пространственной специфичностью. Наши результаты, кроме того, предполагают, что высокочастотная активность в однократном испытании после визуального представления слова характеризуется скорее широкополосным, чем узкополосным ответом.
Методы
Заявление об этике
Исследование было одобрено институциональным наблюдательным советом Университета Констанца и в соответствии с Хельсинкской декларацией.
Участники
Всего в этом эксперименте MEG приняли участие 24 участника (17 женщин; средний возраст = 22 года, диапазон = 19-26 лет; 21 правша).Трое участников были исключены из-за технических проблем, один из-за чрезмерного шума окружающей среды. Здесь представлены данные остальных 20 участников. Все участники дали письменное информированное согласие до начала эксперимента и получили зачетные единицы курса или номинальную финансовую компенсацию за участие. Все участники были носителями немецкого языка и сообщили о нормальном зрении или его исправлении до нормального, а также об отсутствии в анамнезе неврологических заболеваний.
Части этих данных были опубликованы в [12] в отношении вопросов независимых исследований и анализа.
Дизайн, процедура и материалы
Эксперимент состоял из фазы исследования и последующего теста распознавания. Здесь представлены только данные, полученные на этапе исследования. На этапе исследования участникам предъявляли слова либо визуально (проецируемое по центру экрана), либо слухом (через неферромагнитные трубки к обоим ушам). Продолжительность визуального представления слова определялась продолжительностью соответствующего аудиофайла, то есть временем произнесения слова (средняя продолжительность = 697 мс, с.d. = 119 мс). Каждое слово сопровождалось фиксирующим крестом. Суммарная длительность креста слова и фиксации составила 2000 мс. Участникам было предложено подсчитать количество слогов в слове и указать с помощью нажатия кнопки, имеет ли слово два слога. Вопросительный знак (макс. Продолжительность 1500 мс) побуждал к ответу. Нажатие кнопки завершает отображение вопросительного знака. Перед каждым пунктом предъявлялся фиксирующий крест переменной продолжительности (1000-1500 мс). После фазы кодирования участники выполнили задачу отвлечения внимания и фазу проверки неожиданного распознавания.
Стимулы состояли из 420 несвязанных немецких существительных, сгруппированных в три списка по 140 слов. Половина слов каждого списка состояла из двух слогов, другая половина — из одного, трех или четырех слогов. Два списка были представлены на этапе исследования и один список на этапе тестирования. Распределение списков по этапам изучения или тестирования было сбалансировано для всех участников. Задания были представлены в случайном порядке с ограничением, что не более 5 слов одной и той же модальности и не более 5 слов из одного и того же условия были представлены последовательно.
Сбор и предварительная обработка данных MEG
MEG-данные были записаны с помощью 148-канального магнитометра (MAGNES ™ 2500 WH, 4D Neuroimaging, Сан-Диего, США) в положении лежа на спине внутри комнаты с магнитным экраном. Данные непрерывно записывались с частотой дискретизации 678,17 Гц и полосой пропускания 0,1–200 Гц, а затем субдискретизировались до 300 Гц для уменьшения вычислительной нагрузки. Вся обработка данных перед классификацией выполнялась с помощью FieldTrip [56], набора инструментов MATLAB с открытым исходным кодом для анализа данных MEEG.Данные были разделены на отдельные испытания с периодами от 1500 мс до представления элемента до 4000 мс после представления элемента. Испытания были визуально проверены на предмет артефактов, зараженные испытания были отклонены. После этого испытания были скорректированы на моргания, движения глаз и сердечные артефакты с использованием независимого компонентного анализа (ICA).
Реконструкция источника
Для совместной регистрации с индивидуальным структурным магнитно-резонансным изображением (доступно для 17 из 20 участников; для остальных трех участников мы использовали аффинное преобразование мозга с MNI-шаблоном; Монреальский неврологический институт, Монреаль, Канада), форма головы участника, а также три маркера (назион, левый и правый слуховой проход) и расположение катушек индикатора положения головы (HPI) были оцифрованы перед экспериментом с использованием 3D-сканера Fastrak Polhemus (Polhemus, Colchester, VT , СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ).
Космическая активность от одного пробного источника была реконструирована с помощью формирователя луча LCMV [23] с нормализацией веса (индекс нейронной активности; [23, 57]). Во-первых, пространственный фильтр был рассчитан с использованием реалистичной модели головы с одной оболочкой [58], основанной на отдельном структурном магнитно-резонансном изображении (МРТ) и модели источника с точками сетки, покрывающими весь объем мозга (разрешение: 15 мм). Ковариационная матрица данных рассчитывалась для времени от -500 до 1000 мс относительно предъявления стимула. Впоследствии к единичным испытаниям применялся пространственный фильтр для получения виртуальных электродов во всех точках сетки.
Для классификации колебательной активности были рассчитаны временные частотные представления для каждого виртуального электрода с применением быстрого преобразования Фурье. Активность гамма-диапазона оценивалась с использованием частотного сглаживания (подход с несколькими конусами последовательности Слепана), в результате чего были получены полосы частот шириной 20 Гц с центрами 35, 65, 85, 115 и 135 Гц. Мощность рассчитывалась отдельно для длинных временных окон 250 мс от -500 до 1000 мс, а активность после стимула затем выражалась как относительное изменение базовой мощности.
Классификация случайного леса
Алгоритм случайного леса [24], метод ансамбля, объединяет результаты нескольких классификаторов. Эти так называемые базовые обучающиеся представляют собой деревья классификации и регрессии [59], которые разделяют данные с помощью двоичного разбиения. Целью этого процесса разделения является уменьшение примесей в отношении меток классов в дочерних узлах, которые возникают в результате этого разделения: предпочтительно, чтобы все наблюдения из одного класса приходили в один и тот же узел.При каждом разбиении алгоритм дерева сначала ищет предиктор, который максимизирует чистоту дочерних узлов, а затем лучшую точку разбиения в этом предикторе. В случайных лесах теперь растет множество деревьев; однако каждое из этих деревьев построено на выборке исходных данных, и в каждом разбиении выполняется поиск только случайной подвыборки всех предикторов. Дисперсия, вносимая этой случайностью, приводит к надежному прогнозу агрегированной модели. Кроме того, этот подход позволяет случайному лесу особенно хорошо справляться с высококоррелированными предикторами [60], что представляет особый интерес при работе с данными MEEG.
Кроме того, данные с большим количеством предикторов, чем наблюдений (маленькие задачи n , большие p ) также обрабатываются эффективно, поскольку поиск переменных-предикторов выполняется последовательно [61], что делает этот подход особенно интересным при работе с многомерными источниками. космические данные MEEG. Для каждого предсказателя алгоритм возвращает оценку того, насколько важна эта переменная для предсказания модели. Версия, используемая здесь, основана на уменьшении примесей, введенном переменной-предиктором во всех деревьях, которая измеряется индексом Джини [62, 59, 60].
Классификация случайных лесов была выполнена с использованием модуля scikit-learn для Python [63]. Целью декодирования было классифицировать испытания по модальности стимулов: зрительные или слуховые. Предикторами были тройки [воксель, момент времени, частотная полоса], что дало в целом 16 624 предиктора. Для каждого субъекта более распространенный класс (слуховая или зрительная стимуляция) был подвергнут субдискретизации, так что каждый набор данных содержал равные номера испытаний для обоих случаев. Общее количество испытаний по всем предметам составило 4270 испытаний.
Классификация была встроена в структуру перекрестной проверки по предметам: классификатор был обучен на данных от всех, кроме одного предмета, а затем протестирован на данных этого оставленного предмета. Эта процедура была повторена для всех 20 субъектов, так что каждый набор данных использовался в качестве набора для тестирования один раз. Такой подход гарантирует, что классификатор никогда не тестируется на данных, на которых он был обучен, и, таким образом, контролирует возможное переоснащение классификатора. Кроме того, он позволяет оценить предсказуемость данных в отношении переменной ответа для разных субъектов.
Каждая из 20 моделей перекрестной проверки агрегировала результаты 15 000 классификационных деревьев, где каждое дерево было построено на основе начальной выборки всех наблюдений в обучающем наборе. Чтобы гарантировать, что модель включала достаточное количество деревьев, эффективность классификации была оценена с 25 000 деревьев для двух складок [64], что дало результаты, сопоставимые с более разреженной моделью. При каждом двоичном разбиении алгоритм рассматривал предикторы, чтобы найти лучшее разбиение. Точности тестовых наборов данных, а также значения переменных были объединены в свернутых сводках перекрестной проверки.Затем производительность классификатора была проверена на уровне вероятности 50% с использованием биномиального теста [65], поскольку тест, основанный на перестановках, был невозможен с вычислительной точки зрения.
Благодарности
Благодарности Мы благодарим Анн-Кристин Ромбах, Леону Хеллвиг, Марину Кёпфер и Джанин Вейхерт за помощь в сборе данных, а также Сабину Леске и Цветана Попова за ценные обсуждения. Этот проект был поддержан ERA-Net NEURON через Федеральное министерство образования и исследований Германии (BMBF, грант 01EW1307 для SSD) и Европейский исследовательский совет (стартовый грант 640488 для SSD).Это исследование было дополнительно поддержано грантом программы Emmy Noether от Deutsche Forschungsgemeinschaft, присужденным SH (HA 5622 / 1-1). SH получает поддержку Консолидирующего гранта Европейского исследовательского совета (Соглашение о гранте 647954), Общества Вольфсона и Королевского общества. TS финансируется программой исследований и инноваций Европейского Союза Horizon 2020 в рамках грантового соглашения № 661373.
Ссылки
- [1] .↵
Gray CM, Singer W. Стимул-специфические нейронные колебания в ориентационных столбцах кошки зрительная кора.Труды Национальной академии наук. 1989; 86: 1698–1702.
- [2] .↵
- [3] .↵
- [4] .↵
- [5] .↵
- [6] .↵
- [7] .↵
- [8]. ↵
- [9] .↵
- [10] .↵
- [11] .↵
- [12] .↵
- [13] .↵
- [14] .↵
- [15] .↵
- [16] .↵
- [17] .↵
- [18] .↵
- [19] .↵
- [20] .↵
- [21] .↵
- [22] .↵
- [23].↵
- [24] .↵
- [25] .↵
- [26] .↵
- [27] .↵
- [28] .↵
- [29] .↵
- [30] .↵
- [31] .↵
- [32] .↵
Оливетти Э., Киа С.М., Авесани П. Декодирование MEG по предметам. В: Распознавание образов в нейровизуализации, Международный семинар 2014 г. ; 2014. с. 1–4.
- [33] .↵
- [34] .↵
- [35] .↵
Bentlemsan M, Zemouri ET, Bouchaffra D, Yahya-Zoubir B, Ferroudji K.Случайный лес и общие пространственные паттерны банка фильтров для классификации движений на основе ЭЭГ. В: Интеллектуальные системы, моделирование и имитация (СМИБ), 5-я Международная конференция, 2014 г. ; 2014. с. 235–238.
- [36] .↵
- [37] .↵
- [38] .↵
- [39] .↵
- [40] .↵
- [41] .↵
- [42]. ↵
- [43] .↵
- [44] .↵
- [45] .↵
- [46] .↵
- [47] .↵
- [48] .↵
- [49].↵
- [50] .↵
- [51] .↵
- [52] .↵
- [53] .↵
- [54] .↵
- [55] .↵
- [56] .↵
- [57] .↵
Секихара К., Нагараджан СС. Адаптивные пространственные фильтры для электромагнитной визуализации мозга. Springer Science & Business Media; 2008.
- [58] .↵
- [59] .↵
Брейман Л., Фридман Дж., Стоун С.Дж., Ольшен Р.А. Деревья классификации и регрессии. CRC пресс; 1984.
- [60].↵
Катлер А, Катлер Д. Р., Стивенс-младший. Древовидные методы. В: Анализ многомерных данных в исследованиях рака. Springer New York; 2009. с. 1–19.
- [61] .↵
- [62] .↵
- [63] .↵
- [64] .↵
- [65] .↵
Сенсорные рецепторы | Безграничная анатомия и физиология
Классификация рецепторов по стимулу
Сенсорные рецепторы в первую очередь классифицируются как хеморецепторы, терморецепторы, механорецепторы или фоторецепторы.
Цели обучения
Различать типы раздражителей, на которые реагируют рецепторы
Основные выводы
Ключевые моменты
- Хеморецепторы обнаруживают присутствие химикатов.
- Терморецепторы обнаруживают изменения температуры.
- Механорецепторы обнаруживают механические силы.
- Фоторецепторы обнаруживают свет во время зрения.
- Более конкретными примерами сенсорных рецепторов являются барорецепторы, пропиоцепторы, гигрорецепторы и осморецепторы.
- Сенсорные рецепторы выполняют в нашем теле бесчисленное множество функций, обеспечивая зрение, слух, вкус, осязание и многое другое.
Ключевые термины
- фоторецептор : специализированный нейрон, способный обнаруживать свет и реагировать на него.
- механорецептор : Любой рецептор, который предоставляет организму информацию о механических изменениях в окружающей его среде, таких как движение, напряжение и давление.
- барорецептор : нервное окончание, чувствительное к изменениям кровяного давления.
Сенсорные рецепторы можно классифицировать по типу стимула, который вызывает ответ в рецепторе. В целом сенсорные рецепторы реагируют на один из четырех основных стимулов:
- Химические вещества (хеморецепторы)
- Температура (терморецепторы)
- Давление (механорецепторы)
- Свет (фоторецепторы)
Схема классов сенсорных рецепторов : Сенсорные рецепторные клетки различаются по морфологии, расположению и стимулу.
Все сенсорные рецепторы полагаются на одну из этих четырех способностей для обнаружения изменений в окружающей среде, но могут быть настроены на обнаружение конкретных характеристик каждого для выполнения определенной сенсорной функции. В некоторых случаях механизм действия рецептора не ясен. Например, гигрорецепторы, которые реагируют на изменения влажности, и осморецепторы, которые реагируют на осмолярность жидкости, могут делать это через механосенсорный механизм или могут определять химические характеристики окружающей среды.
Сенсорные рецепторы выполняют в нашем организме бесчисленное множество функций. Во время зрения фоторецепторы палочки и колбочки реагируют на интенсивность и цвет света. Во время слуха механорецепторы волосковых клеток внутреннего уха улавливают колебания, исходящие от барабанной перепонки. Во время вкуса сенсорные нейроны наших вкусовых рецепторов определяют химические свойства наших продуктов, включая сладость, горечь, кислинку, соленость и умами (острый вкус). Во время обоняния обонятельные рецепторы распознают молекулярные особенности доносящихся запахов.Во время прикосновения механорецепторы кожи и других тканей реагируют на изменение давления.
Классификация сенсорных рецепторов
Адекватный стимул
Соответствующий стимул можно использовать для классификации сенсорных рецепторов. Адекватный стимул сенсорного рецептора — это стимул, для которого он обладает адекватным аппаратом сенсорной трансдукции.
| Сенсорные рецепторы с соответствующими стимулами, на которые они реагируют. | |
|---|---|
| Рецептор | Стимул |
| Апмуллы Лоренцини (в основном функционируют как электрорецепторы) | Электрические поля, соленость и температура |
| Барорецепторы | Давление в сосудах |
| Хеморецепторы | Химические раздражители |
| Рецепторы электромагнитного излучения | Электромагнитное излучение |
| Электрорецепторы | Электрополя |
| Гидрорецепторы | Влажность |
| Инфракрасные рецепторы | Инфракрасное излучение |
| Магниторецепторы | Магнитные поля |
| Механорецепторы | Механическое напряжение или деформация |
| Ноцицепторы | Повреждение или угроза повреждения тканей тела (приводит к восприятию боли) |
| Осморецепторы | Осмолярность жидкостей |
| Фоторецепторы | видимый свет |
| Проприорецепторы | Чувство положения |
| Терморецепторы | Температура |
| Ультрафиолетовые рецепторы | Ультрафиолетовое излучение |
Расположение
Сенсорные рецепторы можно классифицировать по местонахождению:
- Кожные рецепторы — это сенсорные рецепторы, обнаруженные в дерме или эпидермисе.
- Мышечные веретена содержат механорецепторы, определяющие растяжение мышц.
Морфология
Соматические сенсорные рецепторы у поверхности кожи обычно можно разделить на две группы в зависимости от морфологии:
- Свободные нервные окончания характеризуют ноцицепторы и терморецепторы.
- Инкапсулированные рецепторы состоят из остальных типов кожных рецепторов. Инкапсуляция существует для специализированного функционирования.
Скорость адаптации
Тонический рецептор — это сенсорный рецептор, который медленно адаптируется к стимулу, а фазический рецептор — это сенсорный рецептор, который быстро адаптируется к стимулу.
Классификация рецепторов по местонахождению
Некоторые сенсорные рецепторы можно классифицировать по физическому расположению рецептора.
Цели обучения
Различать сенсорные рецепторы по местоположению
Основные выводы
Ключевые моменты
- Сенсорные рецепторы, расположенные в одном месте, часто выполняют связанную функцию.
- Сенсорные рецепторы кодируют четыре аспекта стимула: модальность (или тип), интенсивность, местоположение и продолжительность.
- Рецепторы кожного прикосновения и рецепторы мышечного веретена являются механорецепторами, но они различаются по расположению.
Ключевые термины
- кожный рецептор прикосновения : Тип сенсорного рецептора, обнаруженный в дерме или эпидермисе кожи.
- мышечное веретено : сенсорные рецепторы в брюшке мышцы, которые в первую очередь определяют изменения длины этой мышцы.
Типы рецепторов
Поскольку мы существуем в мире, наши тела призваны получать, интегрировать и интерпретировать входные данные окружающей среды, которые предоставляют информацию о нашей внутренней и внешней среде.Наш мозг обычно получает сенсорные стимулы от наших зрительных, слуховых, обонятельных, вкусовых и соматосенсорных систем.
Примечательно, что специализированные рецепторы эволюционировали для передачи сенсорных сигналов от каждой из этих сенсорных систем. Сенсорные рецепторы кодируют четыре аспекта стимула:
- Модальность (или тип)
- Интенсивность
- Расположение
- Продолжительность
Рецепторы чувствительны к дискретным стимулам и часто классифицируются как по системной функции, так и по расположению рецептора.
Сенсорные рецепторы находятся по всему нашему телу, а сенсорные рецепторы, расположенные в одном месте, часто имеют общую функцию. Например, сенсорные рецепторы сетчатки почти полностью являются фоторецепторами. Наша кожа включает в себя сенсорные и температурные рецепторы, а наши внутренние уши содержат сенсорные механорецепторы, предназначенные для обнаружения вибраций, вызванных звуком или используемых для поддержания баланса.
Чувствительные к силе механорецепторы служат примером того, как расположение сенсорного рецептора играет роль в том, как наш мозг обрабатывает сенсорные сигналы.Хотя кожные рецепторы прикосновения, обнаруженные в дерме и эпидермисе нашей кожи, и мышечные веретена, которые обнаруживают растяжение скелетных мышц, являются механорецепторами, они выполняют дискретные функции.
В обоих случаях механорецепторы улавливают физические силы, возникающие в результате движения местной ткани, кожные рецепторы прикосновения предоставляют информацию нашему мозгу о внешней среде, а рецепторы мышечного веретена предоставляют информацию о нашей внутренней среде.
Мышечное веретено : Мышечное веретено млекопитающих, показывающее типичное положение в мышце (слева), нейронные связи в спинном мозге (в центре) и увеличенную схему (справа). Веретено — это рецептор растяжения с собственным двигателем, состоящий из нескольких интрафузионных мышечных волокон. Сенсорные окончания первичной афферентной (группа Ia) и вторичной (группа II) афферентной спирали вокруг неконтрактильных центральных частей интрафузальных волокон.
Способов ощущения — Ноцицепторы — Гипералгезия
Сенсорная система состоит из сенсорных рецепторов на периферических окончаниях афферентных нейронов, восходящих путей в спинном мозге и мозговых центров, ответственных за сенсорную обработку и восприятие.Следовательно, он охватывает как центральную нервную систему (ЦНС), так и периферическую нервную систему (ПНС).
Сенсорные рецепторы существуют внутри и снаружи тела и активируются различными стимулами. Они созданы, чтобы реагировать на взаимодействие организма с внешней средой или его внутренним состоянием. Сенсорные рецепторы специализируются на реакции на следующие раздражители: световые, химические, механические, тепловые или ноцицептивные раздражители. Активированный сенсорный рецептор генерирует потенциал действия, который распространяется вдоль аксона и достигает ЦНС.
В этой статье мы рассмотрим различные типы сенсорных рецепторов и некоторые их свойства.
Сенсорная трансдукция
Стимул сенсорного рецептора приводит к изменению ионной проницаемости клеточной мембраны, что приводит к генерации потенциала действия. Этот процесс преобразования сенсорного сигнала в электрический сигнал известен как сенсорная трансдукция . В общем, стимул более высокой интенсивности будет генерировать более высокую частоту потенциалов действия вдоль нейрона.Однако разные типы рецепторов по-разному адаптируются к длительной стимуляции:
- Тонические рецепторы — медленно адаптирующиеся рецепторы. Они будут реагировать на стимул, пока он сохраняется, и производить постоянную частоту потенциалов действия. Следовательно, они передают информацию о продолжительности стимула.
- Фазические рецепторы — быстро адаптирующиеся рецепторы. Они будут быстро реагировать на раздражители, но перестанут реагировать на постоянную стимуляцию.Следовательно, частота потенциала действия снижается при длительной стимуляции. Этот класс рецепторов передает информацию об изменениях раздражителя, например об интенсивности.
Сенсорная модальность
Сенсорные модальности можно рассматривать как подтипы сенсорных ощущений, таких как боль, температура, давление и т. Д. Каждая сенсорная модальность воспринимается классом специализированных рецепторов:
НоцицепторыНоцицепторы — это рецепторы, которые реагируют на вредных стимулов (стимулы, которые могли бы вызвать повреждение тканей, если бы они продолжались), и их активация приводит к ощущению боли .
Рецепторы — это свободные нервные окончания, расположенные на концах волокон типа Aδ и волокон типа C , которые передают болевые ощущения.
Они подразделяются на:
- Механический — стимулируется растяжением кожи (растяжением) и давлением, например. в воспалении
- Термическое — стимуляция крайними температурами
- Химический — стимулируется экзогенными и эндогенными химическими агентами, такими как простаноиды, гистамины и т. Д.
- Полимодальный — может реагировать более чем на один раздражитель
Более подробную информацию о болевых путях можно найти здесь.
МеханорецепторыОни расположены в суставных капсулах, связках, сухожилиях, мышцах и коже и реагируют на деформацию посредством давления , касания , вибрации или растяжения .
Рис. 1. Диаграмма, демонстрирующая расположение различных механорецепторов в коже. [/ caption]- Диски Меркеля : это тонические рецепторы , присутствующие в коже, около границы дермы и эпидермиса.Они реагируют на давление и особенно чувствительны к краям , углам и точкам . Они играют ключевую роль в различении текстур .
- Тельца Мейснера : это фазные рецепторы , присутствующие в дерме кожи, а именно на ладонях рук, подошвах ног, губах и языке. Они обнаруживают первоначальный контакт с предметами или скольжение предметов, удерживаемых в руке.
- тельца Пачини : это фазных рецепторов , присутствующих в дерме, гиподерме, связках и наружных половых органах.Они реагируют на высокочастотную вибрацию и имеют решающее значение для нашей способности обнаруживать вибрации, передаваемые через предметы в наших руках.
- Тельца Руффини: Это тонических рецепторов , присутствующих в дерме, связках и сухожилиях. Они наименее изучены из механорецепторов. Они реагируют на растяжение и сигнализируют о положении и движениях пальцев.
- Мышечные веретена и органы сухожилия Гольджи : они существуют в скелетных мышцах и обнаруживают растяжение .Обнаруженные раздражители способствуют выработке рефлексов на уровне спинного мозга. Кроме того, сигналы передаются в медиальный лемнискальный путь дорсальной колонны (DCML), затем достигают коры головного мозга и предоставляют информацию о позе, положении и ориентации конечностей и суставов в пространстве — проприоцепция .
Терморецепторы
Терморецепторы находятся в коже, печени, скелетных мышцах и гипоталамусе. Они реагируют на перепады температуры.Те, которые реагируют на высокие температуры, присутствуют в волокнах типа C, тогда как те, которые реагируют на холод, присутствуют в волокнах как типа C, так и типа Aδ. Считается, что холодные терморецепторы встречаются примерно в три раза чаще, чем те, которые реагируют на тепло.
Теплый стимул приводит как к увеличению возбуждения для теплых рецепторов, так и к уменьшению возбуждения для холодных рецепторов (и наоборот для холодных стимулов). Первоначально реакция рецепторов изменяется очень быстро в зависимости от незначительных изменений температуры, однако через некоторое время эта фазовая активность переключается на тоническую реакцию , позволяя адаптироваться к температуре, если она остается постоянной.
Температурные стимулы передаются в центральную нервную систему через боковой спиноталамический тракт , часть переднебоковой системы . Дополнительную информацию о восходящих путях, включая эту систему, можно найти здесь.
У всех модальностей есть разные качества, которые можно почувствовать. Например, боль может быть резкой / ноющей, температура может быть горячей / теплой / холодной. Ощущаемое качество зависит от подтипа активированного рецептора, тогда как его интенсивность зависит от силы стимула .
[старт-клинический]
Клиническая значимость — Гипералгезия
Гипералгезия — это повышенная чувствительность к боли, которая может быть вызвана повреждением ноцицепторов или периферических нервов. Он присутствует при многих болезненных состояниях, например, при воспалении, и может действовать как физиологическая защита от дальнейшего повреждения.
Первичная гипералгезия , как полагают, возникает из-за сенсибилизации ноцицепторов и возникает при таких заболеваниях, как ревматоидный артрит. Вторичная гипералгезия считается результатом центральной сенсибилизации и обычно обнаруживается при невропатической боли.
Также важно знать о гипералгезии, поскольку у некоторых пациентов она может быть вызвана опиоидами и является частой причиной снижения эффективности препаратов на основе опиоидов для снятия боли. В отличие от толерантности к опиоидам, увеличение дозы этих препаратов не уменьшает боль, а только усиливает ее, еще больше повышая чувствительность.
[окончание клинической]
сенсорный_рецептор
В сенсорной системе сенсорный рецептор — это структура, которая распознает стимул во внутренней или внешней среде организма. В ответ на стимулы сенсорный рецептор инициирует сенсорную трансдукцию, создавая градиентные потенциалы или потенциалы действия в той же или соседней клетке.
Рекомендуемые дополнительные знания
Структура
Сенсорный рецептор может быть специализированной частью плазматической мембраны, целой клеткой, связанной с окончанием нейрона, или группой таких клеток.
Функции
Сенсорные рецепторы , отвечающие за вкус и запах, содержат рецепторы, которые связываются с определенными химическими веществами. Рецепторы запаха в нейронах обонятельных рецепторов, например, активируются путем взаимодействия с молекулярными структурами молекулы запаха. Точно так же вкусовые рецепторы (вкусовые рецепторы) во вкусовых рецепторах взаимодействуют с химическими веществами в пище, создавая потенциал действия.
Другие рецепторы, такие как механорецепторы и фоторецепторы, реагируют на физические раздражители.Например, фоторецепторные клетки содержат специализированные белки, такие как родопсин, для преобразования физической энергии света в электрические сигналы. Некоторые типы механорецепторов активируют потенциалы действия при физическом растяжении их мембран.
Сенсорный рецептор функционирует как первый компонент сенсорной системы.
Сенсорные рецепторы реагируют на определенные стимулы. Модальность стимула, на которую реагирует сенсорный рецептор, определяется адекватным стимулом сенсорного рецептора.
Сенсорный рецептор реагирует на свои стимулы, инициируя сенсорную трансдукцию. Это может быть достигнуто за счет чистого сдвига в начальных состояниях рецептора (см. Изображение этих предполагаемых состояний [1] с биофизическим описанием — ссылка [2]).
Классификация
по адекватному стимулу
Адекватный стимул сенсорного рецептора — это стимул, для которого он обладает адекватным аппаратом сенсорной трансдукции. Адекватный стимул можно использовать для классификации сенсорных рецепторов:
- Ампулы Лоренцини реагируют на электрические поля, соленость и температуру, но действуют в основном как электрорецепторы
- Барорецепторы реагируют на давление
- Хеморецепторы реагируют на химические раздражители
- Гидрорецепторы реагируют на изменение влажности
- Механорецепторы реагируют на механическое напряжение или механическое напряжение
- Ноцицепторы реагируют на повреждение тканей тела, приводящее к восприятию боли
- Осморецепторы реагируют на осмолярность жидкости (например, в гипоталамусе)
- Фоторецепторы реагируют на свет
- Проприорецепторы обеспечивают чувство положения
- Терморецепторы реагируют на температуру: тепло, холод или и то, и другое
по местонахождению
Сенсорные рецепторы можно классифицировать по местонахождению:
по морфологии
Соматические сенсорные рецепторы у поверхности кожи обычно можно разделить на две группы в зависимости от морфологии:
- Свободные нервные окончания характеризуют ноцицепторы и терморецепторы и называются так, потому что терминальные ветви нейрона немиелинизированы и распространяются по дерме и эпидермису.
- Инкапсулированные рецепторы состоят из остальных типов кожных рецепторов. Инкапсуляция существует для специализированного функционирования.
Иннервация
Основная статья: Типы сенсорных волокон
Различные сенсорные рецепторы иннервируются разными типами нервных волокон. Мышцы и связанные с ними сенсорные рецепторы инвертируются сенсорными волокнами типа I и II, в то время как кожные рецепторы иннервируются волокнами Aβ, Aδ и C.
См. Также
Сенсорная модальность — обзор
Конвергенция сенсорной информации в миндалевидном теле
Как упоминалось ранее, латеральное ядро является первичной входной областью в миндалевидное тело и получает информацию об окружающей среде от всех сенсорных модальностей (Fanselow and LeDoux, 1999). Есть свидетельства того, что сенсорная информация достигает миндалины двумя путями: один для быстрых, но менее разборчивых реакций, а второй — для более медленных и более разборчивых реакций (LeDoux, 1996).Согласно LeDoux (1996), более медленный и более разборчивый путь включает передачу сенсорной информации от сенсорных рецепторов к сенсорному таламусу, сенсорной коре и затем к миндалевидному телу. Этот маршрут называют «автомобильным». Более быстрый и грязный путь начинается так же, но включает прямые проекции сенсорного таламуса (или других подкорковых структур) в миндалину, минуя сенсорную кору. Этот маршрут называют «бездорожьем». Информация, передаваемая в миндалину сенсорным таламусом, не обрабатывается сенсорной корой и поэтому имеет более низкое разрешение.Считается, что при обходе сенсорной коры низкоуровневая информация происходит быстрее и позволяет миндалине инициировать быстрые реакции на важные стимулы, основанные на менее подробной сенсорной информации низкого уровня. Таким образом, бездорожье действует, чтобы инициировать немедленную реакцию «лучше перестраховаться, чем сожалеть». Напротив, высокодорожная информация обрабатывается корой головного мозга с более высоким разрешением, что приводит к более позднему, но более подходящему отклику (LeDoux, 1996).
Первые свидетельства того, что миндалевидное тело получает информацию как от сенсорной коры, так и сенсорного таламуса, получены из исследований повреждений на животных.Исследователи научили кролика бояться одного тона, сочетая его с шоком. Очень похожий тон не сопровождался шоком. После многократного кондиционирования кролик будет демонстрировать учащенное сердцебиение до условного тона, но не такого же тона. После кондиционирования слуховая кора была повреждена, и тестирование реакции страха на тон возобновилось. После того, как кролики были поражены, они не могли различать два тона и вместо этого проявляли реакцию страха на оба стимула (Schneiderman et al., 1974). Это открытие предполагает, что кора головного мозга нужна только для тонкого сенсорного распознавания, но миндалевидное тело все еще может выполнять обучение страху на основе сенсорной информации низкого разрешения, отправляемой из таламуса (LeDoux, 1996). Аналогичные результаты были получены на моделях грызунов, измеряющих замораживание в парадигмах обусловливания страха (Phillips and LeDoux, 1992). В совокупности эти результаты предполагают, что повреждение сенсорной коры приводит к менее специфическому обучению страху. Тем не менее, если миндалевидное тело и сенсорный таламус все еще не повреждены, менее разборчивые реакции страха все еще могут быть изучены — по-видимому, через бездорожье.
Исследования на людях также предоставили доказательства, подтверждающие эти два пути к миндалине. Больной Г.Ю. имеет корковую слепоту, которая является слепотой в результате повреждения зрительной коры, а не повреждения глаз. Несмотря на отсутствие осознанного визуального опыта (то есть слепоту), Г.Ю. все еще может различать эмоциональные и нейтральные визуальные стимулы. Активность мозга регистрировали с помощью фМРТ, в то время как G.Y. попросили различать зрительные стимулы в их слепом полуполе.Результаты фМРТ показали, что миндалина и верхний бугорок (подкорковая визуальная структура в среднем мозге) активировались в ответ на предъявление эмоциональных стимулов в слепом полушарии. С другой стороны, когда эмоциональные образы предъявлялись к неповрежденному полушарию, области зрительной коры были активными. Эти результаты подтверждают наличие подкоркового пути, который обходит поврежденную зрительную кору, чтобы предоставить миндалине информацию с низким разрешением об эмоционально значимых стимулах в окружающей среде (Morris et al., 2001).
Чтобы приблизить слепое зрение у здоровых людей, можно использовать процедуру «маскирования», чтобы предотвратить сознательную обработку представленного изображения. Одна группа использовала эту технику, представляя счастливые и испуганные лица в течение 33 мс, а затем сразу же представляя нейтральное лицо в том же месте в течение 167 мс; участники сообщили только о том, что видели нейтральное лицо. Выполнение этой задачи в сочетании с фМРТ показало, что миндалевидное тело более активно реагирует на замаскированные испуганные, а не замаскированные счастливые лица, что снова указывает на то, что миндалевидное тело играет роль в обнаружении важных стимулов без необходимости осознания (Whalen et al., 1998). Данные об активности миндалины в отношении замаскированных угроз впоследствии многократно воспроизводились (Armony et al., 2005; Carlson et al., 2010; Carlson et al., 2009b; Liddell et al., 2005; Morris et al., 1998, 1999). ; Rauch et al., 2000; Sheline et al., 2001; Suslow et al., 2006; Whalen et al., 2004; Williams et al., 2009; Williams et al., 2005). Кроме того, исследования маскировки показали, что активация миндалины бессознательной визуальной угрозой связана с активацией подкорковых зрительных структур, таких как верхний бугорок и пульвинарное ядро таламуса (Liddell et al., 2005; Моррис и др., 1999). Взятые вместе, данные исследований поражений животных, слепого зрения и маскировки предоставляют существенную поддержку быстрому, но низкому разрешению подкоркового пути к миндалевидному телу (LeDoux, 1996; Liddell et al., 2005; Morris et al. ., 2001; Pourtois et al., 2013; Tamietto, de Gelder, 2010; Whalen et al., 1998).
В то время как накопилось количество свидетельств в пользу подкоркового пути к миндалевидному телу, не все признают, что этот путь обязательно отвечает за более быструю и менее подробную обработку эмоциональной информации.Одна из недавних теорий, которая прямо противоположна теории двух маршрутов, — это гипотеза множественных волн (Pessoa and Adolphs, 2010). Пессоа и Адольфс (2010) утверждают, что доказательств наличия подкоркового пути, который обрабатывает эмоциональные стимулы быстрее, чем другие стимулы, недостаточно для всех аспектов эмоциональной обработки. Кроме того, учитывая скорость коркового процесса, необходимость в быстром подкорковом пути неясна. С точки зрения бездорожья считается, что пульвинар передает зрительную информацию с низким разрешением в миндалину.Однако некоторые данные свидетельствуют о том, что пульвинар может обрабатывать информацию с высоким разрешением (Greene and Oliva, 2009; Pessoa and Adolphs, 2010; Pessoa and Ungerleider, 2004). В отличие от высоких и низких дорог к миндалевидному телу, теория множественных волн предполагает, что существует множество различных маршрутов для информации, идущей к миндалевидному телу. Хотя информация поступает из пульвинария, в контексте теории множественных волн пульвинар играет важную роль в передаче информации о стимулах с высоким разрешением в миндалину.
В общем, не существует единой общепринятой теории относительно того, как сенсорная информация достигает миндалины или какой конкретный уровень детализации передается через различные сенсорные пути к миндалевидному телу. Тем не менее, рассмотренные здесь теории подтверждают, что и корковые, и подкорковые структуры проецируются на миндалину. Боковая миндалина получает сенсорные сигналы от соматосенсорной, зрительной, слуховой, вкусовой и обонятельной сенсорной коры, а также от связанных с ней таламических сигналов (LeDoux, 1996, 2007).Несмотря на продолжающиеся споры о количестве маршрутов и точном характере информации, передаваемой этими маршрутами, существует общее мнение, что миндалевидное тело получает огромное количество сенсорной информации, что делает его анатомически идеально подходящим для роли в эмоциональной оценке.
Сенсорная модальность — обзор
Избирательная интерференция
В одном типе парадигмы меры избирательной интерференции дали убедительные указания относительно основной модальности внутренне генерируемого представления ( а именно., будь то зрительный, слуховой или иногда кинестетический). Сигал и ее сотрудники обнаружили, что, когда субъект следует инструкциям по формированию визуальных образов, его способность обнаруживать визуальные сигналы, представленные извне, уменьшается (Segal & Gordon, 1969). Более того, это уменьшение связано с мешающим эффектом визуальных образов как таковых , а не с общим снижением внимания к любой форме внешней стимуляции. Дальнейшая работа показала, что обнаружение визуальных сигналов было нарушено больше при представлении изображений, чем при представлении звуков, и, наоборот, что обнаружение акустических сигналов было нарушено больше при слуховых, чем при визуальных изображениях (Segal & Fusella, 1970).
Такие исследования избирательной интерференции показывают, что ментальные образы действительно используют некоторые из тех же механизмов обработки информации, которые используются при нормальном восприятии внешних стимулов. Заманчиво предположить, что в рамках каждой модальности существует ограниченная способность обработки информации. Следовательно, всякий раз, когда субъект вытесняет часть этой способности для целей формирования мысленных образов или работы с ними, его способность к обработке снижается для внешних сигналов, представленных в этой конкретной модальности, но не (или, по крайней мере, не в той же степени) для представленных сигналов. в другой модальности.
Однако не совсем ясно, что это разделение способности обработки информации строго совпадает с разделением между сенсорными модальностями. Возможно, это подходящее деление только тогда, когда, как в экспериментах Segal et al. , требованием задачи является простое сенсорное обнаружение. Когда требуется более сложная когнитивная интерпретация внешних стимулов, более абстрактная структурная форма результирующих внутренних представлений, а не просто модальность первоначального ввода, может иметь первостепенное значение.Например, внутренние представления, соответствующие визуальному сканированию и кинестетическому отслеживанию формы буквы (например, «А»), имеют много общего, даже если они связаны с различными сенсорными модальностями. То же самое верно и для внутренних репрезентаций, соответствующих пассивному слушанию и субголосному артикулированию акустического имени этой буквы (ср. Wickelgren, 1969). Таким образом, на когнитивном уровне наиболее сильным детерминантом избирательного вмешательства может быть такой абстрактный (т.е., немодальные) структурные аспекты внутренних репрезентаций, такие как пространственные свойства определенной формы (независимо от того, воспринимаются ли они изначально в визуальной или кинестетической форме) или временной паттерн ее условного названия (выраженный в акустической или артикуляционной форме). образом).
Естественным продолжением этих соображений, созвучным понятиям, выделенным Бруксом (Brooks, 1968; см. Также Shepard, в печати), является то, что наиболее важные и общие подразделения в когнитивных процессах находятся между целостными связными системами — возможно, наиболее отчетливо пространственные и языковые системы.Хотя эти две системы наиболее эффективно взаимодействуют с внешним миром посредством различных сенсорных модальностей ( а именно, зрительно-кинестетическая, с одной стороны, и слухово-артикуляторная, с другой), тем не менее ясно, что обе системы способны к действуя достаточно эффективно, используя одни и те же основные методы. Например, объекты и события могут быть довольно точно локализованы в одном и том же физическом пространстве независимо от того, видны ли они, касаются или слышат; и предложения могут привести к одинаковому пониманию независимо от того, произносятся ли они, записываются или представлены с использованием шрифта Брайля.Таким образом, независимо от конкретной модальности коммуникации с внешним миром, мы можем ожидать большего вмешательства внутри этих двух общих систем, чем между этими двумя общими системами, в результате либо (а) большего структурного сходства между внутренними репрезентациями внутри каждой системы, как предложено выше, либо ( б) очевидно отличная анатомическая локализация этих двух систем в правом и левом полушариях мозга человека (Gazzaniga, Bogen, & Sperry, 1965; Kinsbourne, 1971; Sperry & Levy, 1970).
Брукс (1968) использовал измерение избирательной интерференции в своем изобретательном экспериментальном различении между внутренними репрезентациями преимущественно вербального или пространственного характера. Он продемонстрировал, что количество времени, необходимое для проведения серийной классификации, сильно влияет на режим ответа, который должны использовать испытуемые. Таким образом, испытуемые медленнее классифицируют последовательные слова в ранее изученном предложении как существительные или не существительные, когда они должны отвечать устно, произнося слова «да» или «нет», чем когда они могут выразить ту же классификацию пространственно, указывая к расположению соответствующих визуальных символов («y» или «n») на листе ответов.И наоборот, испытуемые значительно медленнее показывают, находятся ли следующие друг за другом углы по краю ранее изученной формы (например, печатная буква «F») вверху или внизу (в отличие от внутренней части фигуры), когда они должны реагировать пространственно. , указывая на позиции «y» или «n» в листе ответов, чем когда они могут ответить устно, сказав «да» или «нет». Еще один вывод — избирательное вмешательство все еще происходит в задаче, требующей пространственных образов, когда ответы управляются тактично, а не визуально, — снова предполагает, что решающим фактором является абстрактная (лингвистическая в сравнении с пространственной) структура, а не конкретная сенсорная модальность внутреннего представления, которыми манипулируют.